Oiseau
Les oiseaux (Aves) sont une classe de vertébrés tétrapodes caractérisée par la bipédie, des ailes et un bec sans dents. Survivants de l'extinction Crétacé-Paléogène, les oiseaux modernes (Néornithes) sont les seuls représentants actuels des dinosaures théropodes, tandis que tous les autres groupes de Dinosaures, qualifiés de « non aviens », sont éteints.
Aves

| Règne | Animalia |
|---|---|
| Sous-règne | Bilateria |
| Infra-règne | Deuterostomia |
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Infra-embr. | Gnathostomata |
| Super-classe | Tetrapoda |
Sous-classes de rang inférieur
- † Archaeornithes *
- † Enantiornithes
- † Hesperornithes
- † Ichthyornithes
- Neornithes (les oiseaux modernes)
L'histoire évolutive des oiseaux fait de ces animaux de petite taille apparus au Jurassique moyen les descendants directs des dinosaures à plumes pourvus d'ailes, d'où leur position dans le clade des dinosaures. Les analyses cladistiques actuelles les lient aux dinosaures saurischiens, parmi ceux-ci à l'ensemble des théropodes (carnivores bipèdes) et, parmi les nombreux sous-clades qui le composent, au groupe des coelurosauriens Maniraptora. Au sein des maniraptoriens, tous les oiseaux sont rassemblés dans le clade des Avialae qui forme, avec ses deux groupes-frères, les Troodontidae et les Dromaeosauridae, le groupe des Paraves (les « presque oiseaux »). La miniaturisation rapide de cette lignée de dinosaures à plumes, en une cinquantaine de millions d’années, est probablement liée à l’évolution accélérée des nouveautés anatomiques (formation d’ailes et d’un plumage complexe, fusion des clavicules en furcula, développement du sternum en bréchet essentiel au vol battu, redistribution de la masse du corps) qui leur ont permis de coloniser de nouveaux habitats dans les arbres, sources de nourriture et abri contre les prédateurs[1].
S'il existe, en 2020-2021, autour de 10 700[2] à 11 150 espèces d'oiseaux recensées[3] (dont 9 700 espèces sauvages réunissant entre 50 et 430 milliards d'individus[4], et dont plus de la moitié sont des passereaux[alpha 1]), très différentes tant par leur écologie que par leurs comportements, chacune d'elles présente un ensemble commun de caractéristiques évidentes permettant de les regrouper, en particulier des écailles cornées et des plumes, une mâchoire sans dents enveloppée d'un étui corné formant un bec, une queue osseuse courte, des membres antérieurs transformés en ailes (fonctionnelles ou non, cette caractéristique étant particulièrement rare chez les vertébrés[alpha 2]) et des membres postérieurs qui servent seuls normalement à la progression sur le sol ou dans l'eau. En outre, ils sont tous ovipares, c'est-à-dire qu'ils pondent des œufs entourés d'une fine coquille dure, et ils sont tous homéothermes permanents[alpha 3].
Au début du XXIe siècle, 12 % des espèces d'oiseaux sont reconnues en danger par l'UICN. Au moins 40 % des espèces connaissent une baisse de leur population. Les principales causes de la disparition des oiseaux sont l'effondrement des populations d'insectes, une ressource alimentaire importante, la perte d'habitat et l'usage généralisé des pesticides[5].
L'anatomie et la physiologie de la majorité des oiseaux tendent à favoriser au maximum leur capacité à voler : réduction de poids (colonne vertébrale caractérisée par le nombre élevé de vertèbres fusionnées en une pièce osseuse unique et très solide, le synsacrum ; absence de vessie, d'ovaire droit fonctionnel, peau dépourvue de glandes tégumentaires à l'exception de la glande uropygienne, allègement du squelette par des os pneumatiques, réduction de la musculature ventrale et dorsale).
Le corps aérodynamique et une forte musculature pectorale sont également des facteurs favorables au vol. Enfin un métabolisme très actif favorisé par une nourriture énergétique et un appareil respiratoire efficace par la présence de sacs aériens leur permettent de produire la puissance élevée et durable nécessitée par le vol. La spéciation de l'avifaune terrestre a cependant donné naissance à quelques oiseaux inaptes au vol (Struthioniformes, manchots, etc.).
Cosmopolites, les oiseaux peuplent tous les milieux, des glaces de l'Antarctique aux forêts équatoriales et aux déserts. Cette répartition a été rendue possible grâce à des adaptations anatomiques, physiologiques et comportementales (en particulier le phénomène migratoire) variées. Cette plasticité phénotypique se retrouve également au niveau de la gamme de taille qui va du Colibri d'Elena faisant 5 cm à l'Autruche d’Afrique pouvant atteindre 2,75 m de hauteur.
Les oiseaux et les mammifères regroupent certaines des espèces qui ont les meilleurs résultats dans les tests d'intelligence animale, avec les perroquets et les corvidés qui ont la faculté d'utiliser des outils. Toutes ces caractéristiques sont étudiées par les ornithologues.
Dénomination
Le mot « oiseau » dérive de l’ancien français oisel (attesté en 1060), lui-même issu du bas latin aucellus, forme syncopée de avicellus et diminutif du latin classique avis, « oiseau »[6]. Le nom avis dérive de la racine indo-européenne °aw-, représentée notamment dans le védique véḥ, l’arménien haw « oiseau » et le grec ancien αἰετός (aietos), « aigle » (racine grecque qu’on retrouve dans le nom du genre Gypaetus)[7]. Quant à l’ornithologie (littéralement la « science des oiseaux »), elle tire son nom du grec ancien ὄρνις, ὄρνιθος (ornis, ornithos), « oiseau », qui désigne aussi bien l’oiseau domestique que l’oiseau de proie[8].
Par « oiseau », on entend habituellement l’ensemble des espèces existantes possédant des plumes, qui sont regroupées dans la sous-classe des Neornithes, mais aussi bon nombre d’espèces disparues, ancêtres ou apparentées aux espèces actuelles.
Le petit de l’oiseau porte le nom d’oisillon ou de poussin. Le jeune oiseau est qualifié de juvénile[9] puis d’immature[10].
Plusieurs espèces d’oiseaux comportent le terme « oiseau » dans un de leurs noms vernaculaires, comme l’Esclave palmiste ou oiseau palmiste, les oiseaux-lyres ou ménures, oiseaux-mouches, oiseaux de Paradis, oiseaux-éléphants ou æpyornis, oiseaux des îles ou paradisiers, l’oiseau royal décrit par Buffon[11]…, mais certaines espèces le sont plus génériquement comme l'appellation « oiseaux des tempêtes » qui désigne les Hydrobatidae[6].
Contrairement à la plupart des autres animaux, chacune des espèces d’oiseaux dispose d’un nom français normalisé unique attribué par la Commission internationale des noms français des oiseaux (CINFO).
On regroupe également les oiseaux en fonction de leur habitat, comme les oiseaux marins, ou de la géographie, ou des biomes (oiseaux tropicaux), s’ils migrent ou s’ils sont sédentaires. Ainsi, on parle d’oiseaux aquatiques, marins, terrestres ; oiseaux migrateurs, de passage, voyageurs, nicheurs ; oiseaux coureurs, grimpeurs, plongeurs, sauteurs ; oiseaux de proie ; oiseaux diurnes, nocturnes.
Morphologie et anatomie
Aspect général

Un oiseau est un animal tétrapode (à quatre membres), au corps couvert de plumes (seule apomorphie le rendant facilement identifiable), avec un bec corné dépourvu de dents. Ses membres postérieurs sont des pattes et ses membres antérieurs des ailes (homologues aux bras humains)[12] permettant à la plupart d'entre eux de voler. L'évolution montre qu'il n'y a pas de corrélation entre la présence de plumes et vol, les plumes ayant précédé le vol de plusieurs dizaines de millions d'années. La participation des plumes dans le vol résulte d'un détournement de fonction de structures préexistantes, phénomène évolutif connu sous le nom d'exaptation[13].
Le type de vol et les performances que ces espèces peuvent réaliser sont très divers. Leur masse varie de quelques grammes pour les oiseaux-mouches à plus de 100 kg pour les autruches.
La morphologie des oiseaux est adaptée à leur habitat et à leur mode de vie. Les oiseaux vivant sur les plages ou dans les marais ont, par exemple, des longues pattes. De la forme du bec, il est possible de déduire le régime alimentaire. De l'étude des ailes, il est facile de déduire, par exemple, si l'oiseau peut voler. S'il est migrateur, ses ailes présentent une extrémité plutôt effilée ; des ailes à l'extrémité plus arrondie témoignent d'un caractère plus sédentaire.
La morphologie de la tête comporte également de nombreux éléments particuliers qui permettent d'identifier les espèces, comme la présence d'une huppe, d'une crête, d'une caroncule, de vibrisses, de barbillons ou d'une cire.
La valeur adaptative de la coloration du plumage des oiseaux terrestres est difficile à appréhender car il faudrait pouvoir quantifier l'importance de multiples pressions de sélection[14]. Celle des oiseaux de mer se caractérise essentiellement par l'absence générale de couleurs vives, en lien avec le camouflage[15]. La coloration presque systématiquement blanche du ventre de ces oiseaux piscivores aurait une valeur adaptative mais, s'il existe quelques cas de plumage blanc ayant un rôle dans le camouflage, dans la communication et dans la thermorégulation, le plumage blanc est le plumage par défaut chez les oiseaux, minimisant ce coût de production de pigments qui ne sont pas essentiels pour le vol, la photoprotection ou l'étanchéité[16]. La coloration peut être vue globalement comme un compromis évolutif entre la résistance aux rayonnements solaires (provoquant une photo-oxydation différentielle des pigments) et la capacité à se dissimuler des prédateurs et à être vu par les partenaires sexuels éventuels[17]. Certaines espèces possèdent de surprenantes plumes décoratives comme les plumes de queue des pailles en queue et des oiseaux-lyre ou celles des paons mâles.
Leur taille ou envergure varie de quelques centimètres à plusieurs mètres (du Colibri d'Elena qui fait 5 cm à l'Autruche d’Afrique qui peut atteindre 2,75 m de hauteur)[18]. Un colibri ne pèse que quelques grammes, sans comparaison avec le plus grand oiseau volant, connu par des restes fossilisés découverts en 1983 en Caroline du Sud ; il s'agit de Pelagornis sandersi, une espèce éteinte, plus proche des canards, oies et cygnes que des albatros et pélicans. Cet oiseau vivait il y a environ 25 à 28 millions d'années, et — ailes déployées — avait une envergure de 6,4 m (de bout d'aile en bout d'aile), soit près du double de celle du plus grand des albatros contemporains, Diomedea exulans, dont l'envergure est de 3,5 m). Son humérus mesurait environ 94 cm de long. Les caractéristiques des vestiges fossiles suggèrent un poids total de 22 à 40 kg et une très bonne aptitude au vol plané et de longue distance. Il aurait vécu de 55 millions d'années jusqu'à environ 3 millions d'années. La cause de son extinction est encore inconnue.
| Morphologie d'un vanneau de Malabar (Vanellus malabaricus) | |
|---|---|
|
Les oiseaux n'ont pas d'organes sexuels externes. Les oreilles sont cachées sous le plumage et sans pavillon externe. Les plumes en forme d'oreille chez les hiboux ne sont pas utiles dans l'audition. |
Le plumage

Les oiseaux sont les seuls animaux à posséder des plumes, leur plumage est d'ailleurs un bon moyen de reconnaître une espèce.
Parmi les différentes sortes de plumes qui forment le plumage, on distingue[19] :
- selon leur aspect :
- les filoplumes (situées sous les plumes ordinaires) ;
- le duvet ;
- les vibrisses (raides et ressemblant à des poils) ;
- les plumes stricto sensu
- selon leur point d'attache :

Les plumages et les différentes sortes de plumes spécialisées qui les composent ont de nombreuses fonctions pour l'oiseau, la plus universelle étant l'isolation thermique, qui participe au maintien de la température interne de ces tétrapodes homéothermes. Pour certaines espèces, le plumage peut offrir une tenue de camouflage efficace. Pour d'autres, le plumage des mâles est utilisé dans les parades nuptiales ou est indispensable à la séduction. Ainsi, les juvéniles et les individus sexuellement immatures disposent généralement d'un plumage particulier qui évolue, le plus souvent graduellement, jusqu'à leur première saison de reproduction[20]. L'état du plumage permet par ailleurs à une femelle de déterminer la santé d'un mâle. Enfin, pour bon nombre d'espèces, les plumes sont indispensables au vol. La forme de certaines plumes permet de savoir si l'espèce est capable de voler ou non.
Le nombre de plumes varie de 1 000 pour un oiseau-mouche à plus de 25 000 pour un cygne. Les plumes représentent un poids important, chez les frégates (plus de 50 % du poids total) et chez les passereaux (environ le tiers)[20]. À part chez les autruches, les toucans et les manchots, l'implantation des plumes n'est pas uniforme : elles poussent sur des surfaces déterminées (ptérylies) ; les zones nues sont appelées aptéries[19].
Le bec

Les oiseaux primitifs ont encore une mâchoire osseuse avec des dents ayant la forme du bec[21].
Chez les oiseaux modernes, cette transformation des maxillaires dentés (mâchoires et mandibules) en bec corné dépourvu de dents s'accompagne d'un développement massif des muscles du gésier, qui remplace une bonne partie des fonctions masticatrices[22]. Ces maxillaires sont recouverts de la rhamphothèque, gaine constituée d'une ou plusieurs lames de corne. Elle comprend la rhinothèque pour la mandibule supérieure ou maxille et la gnathothèque pour la mandibule inférieure ou proprement dite[23]. La corne repousse au fur et à mesure de son usure.
La forme du bec est souvent adaptée au régime alimentaire de l'oiseau : le bec sera ainsi généralement crochu et robuste chez les rapaces, massif et conique chez les granivores, fin chez les insectivores, allongé et sensible chez les espèces qui fouillent dans les sédiments, etc. L'évolution de cette forme en fonction des différenciations du régime alimentaire est déjà étudiée par Darwin sur les pinsons des Galápagos en 1835 et est un processus toujours en cours[24].
La base du bec et les narines sont parfois protégées par une zone molle et nue, souvent colorée, appelée cire[19].
Les pattes
Les pattes sont formées de trois os longs (le fémur, le tibiotarse et le tarsométatarse) et de quatre doigts. Le fémur est l'os proximal. Le tibiotarse vient de la fusion de la partie proximale du tarse avec le tibia. Le tarsométatarse, os long distal de la patte, résulte de la fusion des autres tarses et des métatarses. Les oiseaux sont digitigrades. Ainsi, la partie de la patte en contact avec le sol correspond aux différentes phalanges des doigts et non à la plante du pied comme chez les humains. Les deux pattes sont recouvertes d'écailles ou scutelles, dont la couleur varie selon les espèces, et présentent des doigts munis de griffes. La disposition des doigts des pattes est très variable selon les groupes aviens.
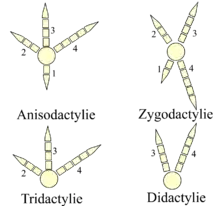
- L'arrangement anisodactyle (trois doigts vers l'avant et un en arrière, aussi appelé hallux) est le plus commun chez les oiseaux et se retrouve tel quel chez leurs ancêtres théropodes, caractérisés par leurs quatre doigts. C'est le cas de la plupart des oiseaux chanteurs, des oiseaux qui doivent se percher et des rapaces.
- L'arrangement syndactyle correspond à une fusion partielle du quatrième et troisième doigt comme chez le Martin-pêcheur d'Amérique et caractérise les Coraciiformes.
- L'arrangement zygodactyle (grec ancien : ζυγον / zugon) « joug », correspond à une migration des doigts, c'est le plus commun des arrangements pour les espèces arboricoles et plus particulièrement celles qui grimpent le long des troncs comme les Picinae, et les Psittaciformes.
- L'arrangement hétérodactyle ressemble au précédent sauf que les doigts 3 et 4 se dirigent en avant et les chiffres 1 et 2 se dirigent en arrière. Cette configuration ne se retrouve que dans les Trogonidae.
- Les pamprodactyles possèdent quatre orteils vers l'avant, c'est une caractéristique des Apodidae.
- La tridactylie est rare et se retrouve dans des espèces très différentes comme la mouette tridactyle ou le pic tridactyle.
- La didactylie se retrouve chez les Paléognathes comme l'Autruche.
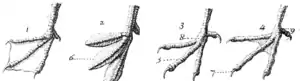
2 – lobée
3 – non palmée
4 – semi-palmée.
En outre, certains oiseaux possèdent une palmure entre leurs doigts comme les sarcelles, les mouettes et les oies. Les espèces anisodactyles, dont les quatre doigts sont reliés par une membrane, comme les fous, les cormorans ou les pélicans sont dites totipalmes. D'autres espèces ont une palmure lobée, comme chez les grèbes et les foulques, ou semi-palmée, comme chez les avocettes, les sternes et les guifettes, ou non palmée, comme chez les martins-pêcheurs, grues, engoulevents, perdrix, rapaces[25].

La forme des pattes dépend essentiellement du mode de vie de l'oiseau et de son régime alimentaire. Ainsi, les rapaces disposent de serres, les oiseaux aquatiques de pattes palmées ou lobées. Les oiseaux coureurs, telle l'Autruche, sont didactyles.
La plupart des espèces possèdent un éperon ou ergot sur les pattes.
Les griffes, les scutelles ou écailles des pattes, l'ergot, tout comme les plumes et le bec, sont des phanères, c'est-à-dire des productions kératinisées issues du système tégumentaire à l'instar de la peau.
Anatomie
Certaines caractéristiques anatomiques des oiseaux (dispositifs physiologiques thermorégulateurs, plumage, appareil reproducteur impair) rappellent celles des reptiles ou celles des mammifères. Ils sont homéothermes et ovipares. L'anatomie des oiseaux montre beaucoup d'adaptations inhabituelles dans le règne animal, dont un grand nombre a pour but de faciliter le vol ; ces adaptations existent même chez les espèces d'oiseaux qui ne savent pas voler.
Les adaptations anatomiques pour le vol se retrouvent évidemment dans la morphologie du squelette de l'oiseau mais aussi dans celle de ses os.
Plusieurs d'entre eux, les os pneumatiques, sont creux et fusionnent avec les sacs aériens qui sont des organes du système respiratoire[26]. En fait, le squelette des oiseaux est très léger (environ 15 % du poids total de l'oiseau). Le système respiratoire des oiseaux est un des plus complexes et des plus performants du règne animal[27]. À la base de la trachée se trouve l'organe vocal des oiseaux, la syrinx.
Les ailes sont également une adaptation au vol mais, comme le précise la théorie de l'évolution, l'aile a précédé le vol. La transformation du membre antérieur en aile s'est accompagnée d'une réduction du nombre d'os, au niveau des phalanges, du nombre de doigts, du carpe et du métacarpe.
On note aussi diverses adaptations au niveau de la colonne vertébrale dont les vertèbres de la région cervicale qui permettent une importante flexion ou rotation du cou pour beaucoup d'oiseaux ; par contre la flexion dans la partie postérieure du corps est très limitée en raison de la fusion de certaines vertèbres (pygostyle et synsacrum). Chez les oiseaux qui volent, le sternum, très développé, porte une crête médiane appelée bréchet, servant à l'insertion des muscles du vol (muscles pectoraux). Un bréchet existe également chez les Chiroptères (chauves-souris), alors que ces mammifères volants n'ont aucune parenté directe avec les oiseaux, ce qui permet de penser que le bréchet serait une adaptation squelettique au vol battu.
Tous les oiseaux, même ceux qui ne volent pas, possèdent une furcula, c'est-à-dire les deux clavicules soudées.
Le squelette est aussi simplifié au niveau des membres postérieurs : le péroné, très réduit, est soudé au tibia ; les os du tarse se sont soudés pour certains avec le tibia (formant le tibio-tarse) et pour d'autres avec le métatarse (formant ainsi le tarso-métatarse).
L'anatomie de leur système digestif répond à la nécessité de satisfaire à un métabolisme élevé[28], le vol étant un moyen de locomotion gourmand en énergie.
Comme les reptiles, les oiseaux sont munis d'un cloaque et, comme eux, les oiseaux sont principalement uricotéliques, excrétant de l’acide urique et des urates et non pas amino-uréotélique (excrétion d'une combinaison d’ammoniaque et d’urée). Parmi les excréments, on trouve de la créatine alors que les mammifères éliminent de la créatinine.
Si, comme pour les mammifères et les Crocodiliens, leur cœur possède quatre chambres[alpha 4], c'est cependant la crosse aortique de droite qui persiste alors que c'est celle de gauche chez les mammifères. De plus, les érythrocytes aviens possèdent des noyaux[27] - [29].

Le système nerveux des oiseaux est, en moyenne, un peu plus volumineux que celui des reptiles mais moins que celui des mammifères. Sauf exception, l'aire visuelle est particulièrement développée, comme en témoignent leurs performances visuelles uniques, et l'aire olfactive est réduite sauf pour certaines espèces comme les vautours ou les kiwis[30]. La taille de leur télencéphale et de leur cervelet leur rend possible des vocalises étonnantes et des capacités d'apprentissage et d'abstraction assez élevées chez certaines espèces. L'œil est chez les oiseaux un organe très complexe. Les organes internes du sens de l'équilibre (canaux semi-circulaires) et de l'audition (cochlée) sont beaucoup plus développés que ceux des reptiles.
L'appareil reproducteur des oiseaux ressemble à celui des reptiles ovipares. La femelle ne possède qu'un seul ovaire fonctionnel, le gauche, tandis que l'ovaire droit peu développé régresse[31].
Certaines adaptations anatomiques peuvent dépendre du mode de vie, comme celles liées à l'alimentation, ce qui est très facilement observable au niveau des becs et des systèmes digestifs. Elles peuvent aussi être liées à la latitude où ils vivent et donc à l'adaptation au froid ; par exemple chez les oiseaux nordiques, la circulation est à contre-courant dans les pattes, de manière à réduire les pertes de chaleur[27]. Certaines de ces adaptations sont temporaires : par exemple certains oiseaux migrateurs ont la capacité de réduire la taille de leurs viscères avant la migration[32].
Biologie de l'oiseau
Alimentation


Les régimes alimentaires aussi bien que les stratégies pour se nourrir sont très variés. Certaines espèces peuvent être opportunistes comme les charognards, ou peuvent trouver leur nourriture dans des lieux spécifiques comme les nectarivores ou frugivores. Les oiseaux peuvent aussi être herbivores, granivores, prédateurs comme les carnivores, insectivores, piscivores, planctonivores, voire cleptoparasites. Le Géospize à bec pointu est même connu pour être hématophage. Certaines espèces piscivores, (Grand cormoran, Spatule blanche, etc.) peuvent chasser ou pêcher en groupe et ont des adaptations morphologiques et anatomiques à la pêche sous-marine (corps profilé, vision polarisée, narines étroites ou obturation nasale, imperméabilisation renforcée du plumage, sacs aériens très développés amortisseurs de choc, squelette du crâne et du cou renforcé)[33]. De nombreux oiseaux sont généralistes ; les autres espèces, dites spécialistes, occupent des niches écologiques spécifiques[34] et ne consomment qu'un seul type de nourriture, ou tout au moins un nombre très limité.
Le métabolisme élevé des oiseaux les oblige à emmagasiner de grandes quantités d'énergie. Ainsi, la masse fraîche de nourriture avalée chaque jour peut équivaloir jusqu'à environ 40 % de la masse corporelle chez les grives, près de 100 % chez les hirondelles, pouillots, roitelets et troglodytes et 200 % chez les colibris[35]. La résistance au jeûne est de fait généralement faible, par exemple 24 h ou moins chez les hirondelles et le Martinet noir, mais peut atteindre plusieurs jours, voire plusieurs semaines, chez les rapaces diurnes[35].
Comme ils n'ont pas de dents, les oiseaux avalent souvent leurs aliments entiers. À la différence des mammifères, ils n'ont pas de voile du palais mou, et doivent donc rejeter leur tête en arrière pour avaler. La nourriture ingérée par becquée peut aussi être écrasée à l'aide du bec, dépecée, plumée, déchiquetée, décortiquée, selon les espèces considérées et la nature de l'aliment. Ils digèrent et broient leurs aliments dans leur estomac à deux chambres : la première dite « estomac glandulaire » assure la digestion chimique par l'action corrosive du suc gastrique (enzymes et acides), la seconde appelée « gésier musculaire » correspond au gésier assurant la digestion mécanique. Les aliments y sont triturés et brassés. Certains oiseaux comme les volailles avalent volontairement de petits cailloux, les gastrolithes, qui sont mélangés avec la nourriture pour faciliter sa fragmentation. Cette lithophagie compense la perte des dents chez ces animaux qui ne peuvent plus mastiquer[36].
Le système digestif aviaire a comme particularité d'être sensible aux toxines alimentaires (assimilation passive et peu sélective), ce qui limite la niche alimentaire des oiseaux. Certains consomment intentionnellement de la terre (phénomène de géophagie connu chez les oies, perroquets, pigeons et certains passereaux), l'argile ingérée étant connue pour former une couche protectrice sur les parois intestinales et empêcher les toxiques de pénétrer dans le sang[37]. L'élimination des selles se fait par le cloaque en même temps que l'urine. Les fientes sont un mélange d’urine et de matières fécales[38]. La couleur en partie blanche est due à l'élimination de l’ammoniac en acide urique[38]. À l'exception des Autruches[39] et des Nandou, les oiseaux ne possèdent pas d'équivalent de vessie[40]. L'urine est évacuée par péristaltisme (contractions musculaires) directement vers le cloaque[40].
Les oiseaux peuvent ingérer des aliments volumineux et capturer rapidement des proies grâce à deux adaptations qu'ils partagent avec les reptiles, la kinésie crânienne et la rhynchokinésie (du (grec ancien : ῥύγχος / rhunkos) « bec » et (grec ancien : kíνησις / kinésis) « mouvement »), qui permettent de mouvoir les deux mandibules de manière complexe[41].
Homéothermie
Les oiseaux sont des animaux homéothermes, c'est-à-dire que leur température interne est maintenue constante, quelle que soit la température extérieure. Leur métabolisme élevé et leur plumage participent à cette régulation ; la température interne moyenne des oiseaux est assez élevée, de 38 à 45 °C mais en général entre 40 et 42 °C soit 3 à 5 °C de plus que celle des mammifères.
Génétique
Le système de détermination sexuelle est de type WZ/ZZ[42]. Dans le système WZ de détermination sexuelle, les femelles ont deux chromosomes sexuels de types différents (WZ), alors que les mâles ont deux chromosomes sexuels de même type (ZZ). Cependant le schéma général de détermination des sexes est mal connu et très différent de celui observé chez les mammifères[42].
Le génome des oiseaux est original sur plusieurs points, la taille physique du génome est faible parmi les vertébrés et de 2 à 8 fois plus courte que chez les mammifères[43]. Le génome est assez homogène avec 60 à 75 % de séquences uniques[43]. La gamme de variations du nombre diploïde est assez étendue : de 40 pour l’Œdicnème criard (Burhinus oedicnemus) à 138 pour le Martin-pêcheur (Alcedo atthis)[43] avec une distribution de type gaussien avec un maximum vers 85. Une des caractéristiques génétiques majeure et originale du génome avien est l'hétérogénéité de tailles des chromosomes. Le rapport entre le plus court et le plus long chromosome est de 1 pour 35 chez la Poule domestique et de 1 à 15 chez les Accipitridae[43]. Les microchromosomes des caryotypes classiques d’oiseaux, contrairement à ce qui était supposé dans les premières études cytogénétiques, constituent des éléments majeurs du génome[43].
Les éleveurs d'oiseaux sont particulièrement attentifs à la génétique de leurs oiseaux pour essayer de faire exprimer certains allèles visibles qui augmentent la valeur de ceux-ci. Cette pratique est courante pour de nombreux passereaux exotiques et les Psittacidae chez qui les éleveurs sélectionnent des couleurs insolites, c'est par exemple le cas de l'Inséparable rosegorge[44].
La génétique s'avère essentielle afin de déterminer leur phylogénie précise et donc de comprendre leur évolution. La nouvelle taxonomie dite de Sibley-Ahlquist est fondée sur les données résultantes des études utilisant les techniques d'hybridation de l'ADN.
Rythmes biologiques

La vie des oiseaux s'organise en fonction de plusieurs rythmes biologiques. Le plus commun aux vertébrés est le rythme circadien. La plupart des oiseaux sont diurnes, mais quelques oiseaux, comme la plupart des hiboux et des Caprimulgidae, ainsi que de nombreuses chouettes, sont nocturnes ou crépusculaires. D'autres espèces, comme la plupart des limicoles, suivent un rythme de vie basé sur la marée[45].
Les oiseaux, en raison de l'existence des saisons, suivent également un rythme circannuel. Les oiseaux qui migrent sur de longues distances subissent généralement des changements anatomiques ou comportementaux comme le Zugunruhe ou une mue pour préparer ce voyage. Suivant les saisons, certaines espèces peuvent également effectuer des migrations journalières, altitudinales par exemple, ou pour se rendre sur des zones de nourrissage.
Les rythmes circadiens et saisonniers semblent liés à la durée du jour. Les cycles de reproduction sont annuels, plusieurs nidifications pouvant avoir lieu dans une saison pour certaines espèces particulièrement prolifiques.
Sommeil
L'activité journalière de la majeure partie des oiseaux comporte des périodes de repos et des périodes courtes de sommeil. Cette période est appelée sommeil vigilant. Les oiseaux ouvrent fréquemment les yeux et observent les alentours pour pouvoir fuir en cas de menace. Le sommeil des oiseaux présente une phase de sommeil paradoxal courte que l'on estime en moyenne à moins d'une minute[46], même si les Spheniscidae, qui détiennent un record dans la classe, capitalisent jusqu'à 80 minutes de sommeil paradoxal par jour[46]. Certaines espèces, comme les albatros ou les martinets, sont réputées pouvoir dormir en volant[46] - [47] ou sont capables de rester plusieurs jours sans dormir. Il n'existe cependant pas de preuve expérimentale. Deux théories principales existent. L'une postule que les oiseaux seraient capables d’alterner veille et sommeil de façon très rapide. L'autre propose qu'ils mettraient en sommeil alternativement leurs hémisphères cérébraux droit et gauche, ce sommeil unihémisphérique pouvant expliquer que la plupart des oiseaux dorment en gardant un œil ouvert[48].

Le comportement associé au sommeil est très différent selon les familles d'oiseaux. Beaucoup d'espèces sont capables de faire diminuer la température de leur corps[49], voire d'entrer en torpeur comme les Trochilidae[50] tandis que l'Engoulevent de Nuttall hiberne[51]. Les Anserinae sont les seuls oiseaux connus chez qui s'observe une perte de tonus musculaire. L'état de sommeil est très reconnaissable chez ces espèces qui réfugient leur bec sous une aile et souvent ne posent qu'une seule de leurs pattes au sol[52]. Beaucoup d'espèces nicheuses au sol, comme les Phasianinae, se réfugient en hauteur à l'abri des prédateurs, quelquefois en commun, pour dormir, tandis que quelques perroquets du genre Loriculus se perchent tête en bas comme des chauves-souris[53]. De nombreuses espèces présentent une insertion particulière des tendons fléchisseurs des muscles des doigts de sorte que, lorsqu'ils sont posés sur une branche, en pliant les « pattes » sous le poids du corps, les tendons (en position « relax ») « verrouillent » mécaniquement les doigts qui s'agrippent ainsi fermement autour de la branche ; la flexion automatique des doigts leur permettrait de dormir perchés sans tomber de l'arbre[19].
Le renouvellement du plumage

Le plumage est régulièrement renouvelé, au minimum une fois par an[20], au cours de la mue. Suivant les espèces, celle-ci peut avoir plusieurs rôles. Elle permet à certaines espèces de changer de couleurs de plumage, aussi bien pour la reproduction que pour se camoufler, mais aussi d'obtenir une meilleure couverture thermique. Elle peut aussi précéder une migration. Les oiseaux réalisent aussi une mue pour passer du plumage juvénile au plumage adulte.
En dehors de la mue, avant de nicher, les femelles de nombreuses espèces perdent le duvet situé sous leur ventre. Sous la peau, à cet endroit précis alors dénudé que l'on appelle plaque incubatrice, se trouvent de nombreux vaisseaux sanguins. Les œufs pourront ainsi être en contact plus étroit avec la peau réchauffée par les vaisseaux sanguins : ceci favorise une bonne incubation[54].
La perte de plumes, tout comme l'absence de mue, peut être également le signal d'une maladie.
La migration

La migration est un déplacement régulier, d'annuel à journalier, observé chez de très nombreuses espèces d'oiseaux. Ces déplacements peuvent prendre des formes très différentes selon les espèces et induisent d'autres comportements, alimentaires par exemple, très spécifiques. C'est un effort important pour bon nombre d'oiseaux qui peuvent même en mourir. Bien connues des hommes, ces migrations sont attendues par les chasseurs, craintes par les gestionnaires d'aérodromes et certains aspects confondent toujours les scientifiques, qui cherchent encore à comprendre comment font les oiseaux pour parvenir à leur destination.
Ces déplacements sont une manière pour les espèces migratrices d'échapper à un changement d'habitat ou une baisse de disponibilité de nourriture liée aux rigueurs d'un climat défavorable, mais peuvent aussi s'interpréter comme une maximisation des chances de reproduction.
Activités de confort
Les oiseaux utilisent une partie de leur temps dans des activités dites de confort. Selon certains auteurs, ils consacrent 9,2 % de leur période active à cela[55].
Une des activités de confort les plus courantes est le lissage de leurs plumes avec les sécrétions cireuses de leur glande uropygienne. L'utilité de cette pratique est discutée mais il semble que cette cire agit sur la flexibilité des plumes et aussi comme un agent antimicrobien qui inhibe la croissance de bactéries dégradant les plumes[56]. Plus de 250 espèces complètent ces sécrétions avec de l'acide formique tiré de fourmis[57]. Les séances de toilettage peuvent être mutuelles.

De nombreux oiseaux aiment également se baigner dans de l'eau et certains, notamment les Phasianidae et les moineaux, dans la terre ou la poussière.
Longévité de l'oiseau
La durée de vie des oiseaux est très variable selon les espèces. Elle peut être de 3 ou 4 ans pour certains passereaux (mais jusqu'à 25 ans pour le Cardinal à poitrine rose[58]) à plus de 50 ans pour les albatros et les puffins ou encore plus de 60 ans pour certaines espèces comme le kakapo ou les perroquets ara. Les psittaciformes (perroquets) présentent de très grandes longévités en captivité[59]. Un cacatoès à huppe rouge atteint l'âge de 92 ans[60] mais le record de longévité semble revenir à un ara bleu de 104 ans[61].
Les records de longévité sont obtenus en captivité, car les oiseaux sauvages sont soumis aux pressions environnementales (prédateurs, maladies, braconnage, accidents). Toutes les espèces ne subissent pas les mêmes pression. Par exemple, le risque annuel d'être tué pour des passereaux en zone tempérée est de 70 % alors qu'il descend à 3 % chez l'albatros royal dont l'espérance de vie peut atteindre 58 ans[62].
Au sein d'une même espèce, les mues dépendent des saisons, mais aussi de l'âge des oiseaux et de l'état général de ceux-ci. Ainsi, la bonne connaissance de ce phénomène permet de déduire l'âge de nombreux oiseaux sauvages[63]. La pneumatisation du squelette peut également servir à estimer l'âge des oiseaux[64].
Parasites des oiseaux
Les parasites les plus courants chez les oiseaux appartiennent aux groupes des acariens, des poux aviaires et des vers. D'autres parasites microscopiques, comme certains protozoaires, provoquent des maladies.

Au moins 2 500 espèces d'acariens réparties dans 40 familles[66] vivent en relation étroite avec les oiseaux, occupant leurs nids, leurs plumes ou même leur bec, comme certains acariens d'oiseaux-mouches. Ces acariens peuvent avoir une relation simplement phorétique ou peuvent perturber leurs hôtes en provoquant des démangeaisons, mais peuvent aussi être des parasites comme les Dermanyssus et Ornithonyssus. Toutes les espèces d'oiseaux sont concernées, même les manchots[66] possèdent des tiques. Le mode de vie d'une tique d'oiseau dépend bien sûr de son espèce ; cependant la larve vit presque uniquement dans le nid. Ces acariens ont des cycles de reproduction courts et sont capables de pulluler très rapidement. Certains acariens sont hémophage et se nourrissent de peaux mortes comme Dermanyssus, d'autres, comme chez les oiseaux-mouches, se font transporter de fleur en fleur et s'y nourrissent de nectar. Dans les nids, on a même découvert des tiques naines parasites de tiques aviaires[66].
Un nombre trop important de tiques peut nuire à la couvée et même à la vie du poussin[66]. Pourtant, certaines études pourraient suggérer que ce commensalisme n'est pas uniquement défavorable aux oiseaux[66]. Il existe de nombreuses études sur ce sujet complexe des interactions entre les acariens et les oiseaux, beaucoup de facteurs entrant en ligne de compte. Il n'est donc pas facile d'établir des règles simples.
Les Ischnocera, insectes aussi appelés « poux broyeurs », sont le plus souvent inféodés à une espèce précise d'oiseau.
Plusieurs espèces de plathelminthes, dont des cestodes ou trématodes, peuvent infecter les oiseaux qui peuvent les transporter d'un continent à l'autre. Par exemple, les oiseaux de mer, en mangeant des coques, favorisent un parasitisme à trématodes (genre Meiogymnophalus[67], Himasthla[68], etc.) qui pourra par la suite atteindre plusieurs espèces d'hôtes, oiseaux ou mollusques
Certains diptères sont ectoparasites et hématophages des oiseaux, leurs cycles de vie étant fortement synchronisés. Quelques imagos pupipares ont une spécificité parasitaire rigoureuse tels que Stenepteryx hirundinis sur les hirondelles ou Crataerina pallida sur les martinets noirs. D'autres, comme Hippobosca equina, peuvent également parasiter des mammifères. Ces espèces sont particulières dans le sens où le développement larvaire s'effectue uniquement dans l'utérus de la femelle et seuls les adultes sont hématophages[69]. Chez la famille Piophilidae, c'est la larve de Neottiophilum praeustum qui vit dans les parois des nids de passereaux et s'accroche périodiquement aux oisillons pour en sucer le sang. Certaines espèces des familles Carnidae et Calliphoridae ont un comportement similaire[70]. Ces espèces sont en principe très répandues. Une espèce ubiquiste comme Ornithomya avicularia est établie sur la région holarctique entière. Les espèces du genre Olfersia (en) subissent les immenses déplacements des oiseaux océaniques comme les Frégates, Pétrels et Albatros, leurs hôtes habituels. À l'inverse, Crataerina pallida ne suit pas les migrations du Milan noir et vit uniquement dans ses zones de nidification[69]. Il est possible qu'un grand nombre d'individus puisse entraver le développement des oisillons ; cependant de nombreuses études suggèrent un impact minime[70] - [71]. Par contre, ils peuvent transmettre des germes pathogènes à leurs hôtes. C'est notamment le cas de Pseudolynchia canariensis (en) qui inocule le protozoaire Haemoproteus columbae (en) aux pigeons[69] - [72].
Maladies aviaires

Comme d'autres espèces animales domestiquées (volailles notamment) ou sauvages[73] (insectes, mammifères notamment), les oiseaux peuvent jouer un rôle épidémiologique limité[74] et écoépidémiologique important vis-à-vis des humains en étant vecteurs et souvent porteurs-sains de maladies et parasites qu'ils peuvent propager à longues distances via leurs migrations. C'est le cas par exemple de l'ornithose, la salmonellose, le campylobactériose, la mycobactériose (tuberculose aviaire), la grippe aviaire, la lambliase, et la cryptosporidiose[75] ou de diverses maladies émergentes comme la maladie de Lyme[75].
Ces zoonoses sont donc étudiées, sous l'égide de l'OIE et de l'OMS et leur propagation soigneusement observée du fait de l'importance économique et écologique des élevages de volaille (victimes de maladies « sauvages » et sources de maladies émergentes parfois problématiques (avec antibiorésistance, risque nosocomial ou apparition de virus réassorti). C'est pourquoi les découvertes de foyers de maladies aviaires peuvent amener les autorités sanitaires et/ou vétérinaires à prendre des mesures radicales vis-à-vis de ces élevages : on apprend régulièrement par les journaux l’abattage de centaines de milliers de volailles (canards, poules ou dindes) pour endiguer ces épidémies. Certaines maladies peuvent être plus spécifiques à un ordre comme la maladie de Pacheco pour les Psittaciformes.
Autres maladies :
- Paludisme aviaire
- Maladie de Newcastle
- Mycoplasmose
- Trouble de la kératine aviaire
- choléra des poules ou aviaire
- Maladie de Gumboro
- Laryngotrachéites[76] de l'oiseau
- Variole aviaire (forme diphtérique)
- PBFD
- PSD
- Candidose aviaireCandidose aviaire[77].
- Aspergillose
Comportement
Les sens
Les sens des oiseaux ne diffèrent pas fondamentalement de ceux des mammifères, mais pour certains restent mal connus : on ne sait pas très bien, par exemple, comment les oiseaux parviennent à se repérer lors de leur migration.
En revanche, on sait que leur vision est souvent remarquable. Ils ont une vision à la fois monoculaire et binoculaire combinée. Certaines espèces perçoivent les ultraviolets[78]. Les rapaces notamment ont une acuité visuelle importante, deux à trois fois plus élevée que celle de l'homme[78] ; la fovéa d'une buse variable possède environ 100 000 cônes par millimètre carré, contre 20 000 chez l'homme[35], soit 5 fois plus. Les yeux des oiseaux sont très volumineux ; ceux de l'étourneau, par exemple, ont une masse correspondant à 15 % de la masse crânienne (à titre de comparaison, ils ne représentent que 1 % de la masse crânienne chez l'homme)[35]. Les oiseaux possèdent une membrane nictitante, ainsi qu'un organe interne, situé dans l'humeur vitrée, appelé peigne, au rôle mal défini. Certaines espèces diurnes possèdent deux fovéas au lieu d'une (par exemple : les hirondelles, sternes, martinets, faucons, colibris…)[35]. Les oiseaux diurnes, dont les yeux sont positionnés plus latéralement, ont une vision binoculaire restreinte à la zone située devant le bec, mais un grand champ panoramique et une grande mobilité de la tête (qui peut tourner parfois sur plus de 250°)[35]. En raison de cette vision, leur perception du relief est réduite, ce qui expliquerait pourquoi ils déplacent la tête d'un côté à l'autre ou la penchent en avant quand ils regardent quelque chose attentivement[79]. Les oiseaux prédateurs tels que les rapaces, qui ont besoin de viser pour tuer leurs proies, ont des yeux frontaux qui assurent un champ binoculaire plus important[80].

À de rares exceptions près (Kiwi, Vautour pape[35]), l'olfaction est, chez les oiseaux, considérée comme peu performante, voire absente. Certaines études[81] - [82] tendent cependant à montrer que les oiseaux de mer de la famille des Procellariidae sont capables de repérer leurs proies, en particulier l'odeur de l'huile de poisson, à l'odorat[83].
Bien que les oiseaux n'aient pas de pavillon auriculaire, leur ouïe est développée, surtout chez certaines espèces comme les Strigidae, chez qui elle est suffisante pour localiser une proie dans l'obscurité complète[78]. Cependant l'absence de pavillon les oblige à effectuer des rotations de la tête pour percevoir les sons provenant de différentes directions. Les Salanganes ou le Guacharo des cavernes sont capables, grâce à leur ouïe performante couplée à leurs cris, de se déplacer par écholocation. Contrairement aux Mammifères, l'oreille moyenne des oiseaux ne présente qu'un seul osselet, la columelle. Au niveau de l'oreille interne, la cochlée n'est pas en colimaçon, mais droite[35].
Les papilles gustatives ne se trouvent pas sur l'extrémité de la langue, mais sur le fond de langue et dans la gorge ; en outre elles sont peu nombreuses (200 pour un Anatinae contre 9 000 pour un humain) mais d'autres mécanismes pourraient être mis en œuvre pour le goût. Ainsi le sens du toucher (notamment au niveau du bec), pour beaucoup d'oiseaux, semble intervenir lors de la recherche de nourriture.
Le sens du toucher est conféré aux oiseaux par divers corpuscules : les corpuscules encapsulés de Merkel (au niveau de la peau et à l'intérieur de la bouche) et de Grandry (au niveau du palais) participent à la sensibilité tactile générale ; ils seraient les correspondants des corpuscules de Meissner des Mammifères[84]. Les corpuscules de Herbst (qui correspondraient aux corpuscules de Pacini des Mammifères[84]) seraient davantage sensibles aux vibrations : ils sont particulièrement nombreux sur le bec et les pattes, particulièrement chez des espèces qui doivent trouver leur nourriture « à tâtons » : langue des Picinae, becs des Anatidae (oies, canards) et de nombreux Scolopacidae (bécasses, bécassines et bécasseaux), mais aussi bourrelets colorés qui bordent le bec des oisillons de nombreux passereaux et Picinae[35].
Les oiseaux possèdent plusieurs organes d'équilibre indépendants, l'oreille interne comme chez les mammifères, et un organe situé dans le bassin.
Un des sens les plus mystérieux est la détection du champ magnétique terrestre, l'organe qui le détecterait serait situé dans le bec ou près des yeux[78]. Ce sens a été pour la première fois démontré expérimentalement en 1967 par Wolfgang Wiltschko sur des rouges-gorges.
Intelligence

Bien qu'« avoir une cervelle d'oiseau » signifie ne pas avoir une grande intelligence dans plusieurs cultures, certaines espèces d'oiseaux font preuve de capacités cognitives relativement élevées. Les Corvidae sont réputés être les plus intelligents des oiseaux[85] ; les Psittacidae sont aussi capables de performances remarquables, mais avec beaucoup d'inégalités selon les espèces. D'autre part, il est difficile de définir le terme « intelligence » et aussi de distinguer ce qui fait partie du domaine de l'inné ou du domaine de l'acquis, et donc d'évaluer leur capacité de raisonnement.
Les oiseaux sont capables d'apprendre ; on sait par exemple que les petits coucous apprennent le chant de leurs parents adoptifs ou que les corbeaux font leur apprentissage en imitant leurs semblables[86].
Les plus communes de leurs capacités sont certainement leur représentation spatiale (qui leur permet de s'orienter, de retrouver une source de nourriture ou de construire des nids sophistiqués) et leurs capacités de communication.
Une des capacités les plus surprenantes est l'aptitude assez répandue[87] à se servir d'un objet comme outil. Le Corbeau calédonien, par exemple, est capable de se servir d'un bâton pour déloger les insectes dont il se nourrit (c'est aussi le seul à savoir tailler ce bâton en fonction de l'arbre).
Certains oiseaux sont même capables de compter, comme les Psittacidae, qui sont aussi connus pour reproduire la voix humaine. Les mainates sont même réputés pouvoir apprendre à parler[88].
On a également observé des oiseaux capables de se soigner, par exemple en ingérant de l'argile destinée à combattre les effets néfastes des toxines alimentaires[89].
Certaines facultés sont pratiquement uniques, le Guêpier d'Orient est capable de se « mettre à la place » de son prédateur, sous-entendu d'extrapoler ce que le prédateur peut voir ou non, faculté partagée uniquement par les Hominidae[90].
Reproduction

L'appareil reproducteur aviaire est caractérisé par l'absence d'organes sexuels externes, si bien qu'il n'est souvent possible de déterminer le sexe des oiseaux que par les caractères sexuels secondaires (couleur des plumes, présence de plumes ornementales, chant, présence de barbillons, crête, taille). La copulation est un processus fugitif (ne durant généralement que quelques secondes[alpha 5]) que les ornithologues appellent « baiser cloacal » : le mâle se balance sur le dos de sa partenaire, vient presser les lèvres de la sortie de son cloaque sur celui de la femelle, et transfère dans l'orifice génital une goutte de son sperme[91] - [92].
Les déplacements
La plupart des oiseaux sont capables de voler, c'est-à-dire de se déplacer dans les airs ; il n'existe qu'une soixantaine d'espèces d'oiseaux coureurs, qui en sont incapables[93]. On estime que tous les ancêtres des oiseaux modernes étaient capables de voler. La perte de cette aptitude, souvent dans des endroits isolés, pourrait être due à l'absence de prédateur[93] (comme pour le Dronte) ou à la faiblesse des ressources alimentaires, rendant périlleuse la grande dépense d'énergie que nécessite le vol[94] ou encore à une adaptation extrême.

Il existe plusieurs techniques de vol qui se traduisent par la forme spécifique des ailes, en fonction du type de vol auquel chaque espèce est adaptée. L'étude de la longueur des os du bras permet d'en préciser l'analyse.
Certaines espèces comme les Spheniscidae (dont les manchots), les petits pingouins, les puffins et les cincles se déplacent sous l'eau comme s'ils volaient[95].
Les autres oiseaux coureurs disposent en général d'os des pattes plus robustes et une absence de bréchet. Une étude attentive du squelette permet de savoir si un oiseau peut voler ou pas ; ces connaissances sont utilisées pour l'étude des fossiles d'espèces disparues.
Personnalité
Une étude britannique sur des verdiers, menée en 2011, suggère que les oiseaux ont individuellement une personnalité, en mettant en évidence différents traits de caractère (courage, timidité, curiosité), qui se reflètent au niveau de leur stress oxydant[96].
Communication
Le degré de sociabilité varie selon les espèces et les saisons. La plupart des espèces sont, à une période de leur vie au moins, sociables, même si les différences de comportement entre espèces sont très importantes. Les oiseaux utilisent, dans ce cadre, plusieurs moyens de communication. Bien que leur plumage en soit un, ils utilisent surtout différents signaux visuels constitués de divers mouvements. Ils peuvent également émettre des vocalisations ou d'autres types de sons. La complexité de ces signaux est maximale lors des parades nuptiales et constitue un des moyens de communication les plus codifiés du règne animal.
Les relations sont principalement de trois types :
- relations parentales ;
- relations mâle-femelle et concurrence entre mâles ;
- relations interspécifiques (plusieurs types de commensalismes, surtout alimentaires, sont connus de la part d'oiseaux, vis-à-vis d'autres espèces, mais également le cleptoparasitisme et le parasitisme des nids).
Types de couple

95 % des espèces d'oiseaux sont monogames. Dans un certain nombre de cas (en particulier chez les grandes espèces à longue durée de vie), ces couples sont constitués pour la vie. Les grues du Japon sont même des symboles de fidélité en Extrême-Orient. D'autres espèces ne sont monogames que pour l'année, on parle de monogamie en série. Parmi les espèces monogames strictes, nombreuses sont celles chez qui la copulation avec d'autres partenaires peut intervenir, selon les circonstances[97]. Dans ces cas, on observe des comportements collaboratifs du père génétique avec le couple[98]. Pour certaines espèces, la monogamie est davantage liée à l'adoption du même territoire qu'à une véritable monogamie[99]. La polygynie se retrouve chez 2 % des espèces, la polyandrie ou la polygynandrie chez moins de 1 % pour chaque. Chez les Anatidae par exemple, le « viol » est également pratiqué[100].
Chez les oiseaux, la monogamie sociale mais pas sexuelle aurait pour origine la nécessité des soins paternels pour assurer le succès reproducteur[101] : « l'association entre homéothermie, oviparité et nécessité d’avoir une croissance très rapide pour raccourcir la période exposée à la prédation rendrait indispensable une collaboration étroite entre les deux sexes pour mener à bien la reproduction »[102].
Si, pour beaucoup d'espèces, les mâles aident les femelles dans l'élevage des petits, ceci n'est pas systématique, mais ce comportement est beaucoup plus courant chez les oiseaux que chez les autres classes de vertébrés. La présence des mâles est donc souvent nécessaire à l'incubation et/ou à l'élevage des oisillons, et dans certains cas les femelles n'y participent pas (cas des phalaropes). Chez certaines espèces, les tâches sont strictement divisées : incubation, défense du nid, nourrissage des petits[103], etc.
L’appariement
Les femelles sont sensibles à de nombreux critères et mettent les mâles en concurrence[104] bien que, chez les Phalaropes, ce sont les mâles qui choisissent les femelles[105].

Un des critères les plus importants est la qualité du territoire proposé par le mâle. Cela peut aller de quelques cailloux pour un manchot à un nid très sophistiqué ou un vaste territoire pour les espèces de rapaces solitaires. Le mâle peut également se mettre en valeur par des vocalisations, des signaux visuels très sophistiqués et codifiés, comme la « danse de ballerine » du Paradisier de Carola, voire des combats. La santé du mâle peut également être évaluée par la couleur des plumes. Chez certaines espèces peu territoriales, ces parades sont faites par des groupes de mâles sur une petite surface appelée aire de parade. Le cas le plus connu est celui du Combattant varié.
L’élevage des jeunes

Après la fécondation, sauf exception, par exemple chez le guillemot de Troïl qui pose son œuf sur la roche nue, la femelle pond dans un nid. Ce nid a des dimensions et une composition variable selon les espèces.
La ponte comprend un nombre d'œufs qui varie selon les espèces de 1 (par exemple chez les Procellariidés ou les grands rapaces) jusqu'à des pontes records de 20 chez la perdrix. Un nombre important d'œufs dans un même nid peut être occasionné par la ponte de plusieurs femelles : ce phénomène est régulier chez l'Autruche et occasionnel chez de nombreuses espèces comme le Tadorne de Belon et le Canard colvert chez les Anatidae.

Ces œufs possèdent une coquille dure, composée surtout de carbonate de calcium, de couleur propre à chaque espèce. Leur dimension est variable, ainsi que leur masse qui varie d'un minimum de 0,027 g chez le Colibri jusqu'à un record de 2 350 g chez l'Autruche.
L'embryon, pour se développer, doit être maintenu à une température constante. La plupart des oiseaux incubent leur(s) œuf(s) en le(s) couvant. Cette pratique peut être gourmande en énergie[106] et a une durée variable (de 9 à 10 jours chez le Gros-bec à un maximum de 82 jours chez l'Albatros hurleur).


Pour certaines espèces dites précoces ou nidifuges, les oisillons sont capables immédiatement de suivre leurs parents, le cas extrême étant celui des Mégapodes où les petits sont complètement indépendants ou celui des oisillons du Guillemot à cou blanc (en) qui abandonnent le nid la nuit après leur éclosion, en suivant les appels des parents vers la mer, où ils sont élevés loin des prédateurs terrestres[107].
A contrario, chez les espèces dites nidicoles comme les Ardeidae, les poussins naissent aveugles et sans plumes. C'est la Frégate du Pacifique qui détient le record de durée de dépendance avec un poussin qui n'acquiert son plumage qu'au bout de six mois et qui est nourri par ses parents durant quatorze mois[108]. Les efforts que doivent faire les parents pour les nourrir sont donc également très différents selon les espèces.
Chez certaines espèces, le couple se fait aider par un autre membre de l'espèce[109]. Ce phénomène est particulièrement présent chez les espèces de Corvidés mais il a été également observé chez des espèces aussi diverses que le Xénique grimpeur, le Milan royal ou la Pie australienne. Les jeunes de la couvée précédente participent régulièrement au nourrissage des poussins de la nichée suivante chez la Poule d'eau.
Durant les quelques heures qui suivent l'éclosion, l'objet animé que l'oisillon voit, est considéré par lui comme son parent. Ce comportement, particulièrement fort chez les oiseaux, s'appelle l’imprégnation.
Relations sociales

La majeure partie des espèces sont sociables, elles peuvent l'être toute l'année comme le Corbeau freux, les sternes, les goélands ou sur une période restreinte de l'année comme les bécasseaux. Dans ce dernier cas, les regroupements peuvent avoir un objectif précis comme la migration ou la nidification. Certaines espèces vivent en groupes pluri-spécifiques, d'autres en groupes de la même espèce. Lorsqu'elles sont grégaires, elles font preuve de comportements sociaux ou collectifs qui peuvent être très complexes comme la chasse en groupe, la constitution de hiérarchie, la coopération filiale dans l'élevage des jeunes, la présence de guetteurs (Bernache du Canada)[110], la constitution de nuées ou volées mono ou pluri-spécifiques, ou tout simplement la constitution de colonies. La hiérarchie s'exprime notamment pour obtenir la meilleure nourriture (hiérarchie de picorage), le meilleur perchoir ou le nid le mieux placé. Les hiérarchies de dominance s'établissent par des postures d'aplomb, d'intimidation voire des combats[111].
Ces comportements sont complémentaires au besoin de territorialité; ainsi les adultes de certaines espèces vivant en colonie deviennent très agressifs vis-à-vis de tout intrus en deçà d'une distance minimale de leur nid (Fou de Bassan) correspondant le plus souvent à la distance que peut atteindre le couveur avec son bec sans quitter le nid.
La nidification en colonies peut être liée à une répartition irrégulière des ressources alimentaires ou au manque potentiel de sites de nidification, ce qui explique que 90 % des oiseaux marins nichent en colonies[112]. L'hypothèse que ce type de nidification apporte comme avantage évolutif une meilleure protection contre les prédateurs (fonction d'alerte des individus en sentinelle à la périphérie de la colonie) n'est pas probante[113].
Parasitismes interspécifiques
Certains oiseaux sont cleptoparasites, c'est-à-dire qu'ils se nourrissent, au moins en partie, de proies dérobées à d'autres espèces. Il s'agit en particulier des Labbes et de espèces de Laridés, goélands et mouettes[114], mais un grand nombre d'espèces sont concernées[115], notamment des rapaces et des limicoles[116].

Certaines espèces d'oiseaux pondent volontairement dans le nid d'autres oiseaux, de la même espèce ou non. Ce parasitisme de couvée existe sous deux formes, certaines espèces sont obligées d'utiliser cette méthode car elles sont incapables d'élever leurs propres jeunes, et d'autres n'utilisent cette méthode que ponctuellement, comme pour certains Phaethornithinae[117]. Il existe environ une centaine d'espèces parmi les Cuculidae, Indicatoridae, Icterinae, Estrildidae, Anatidae qui sont des parasites obligatoires[118] et les oisillons de la plupart d'entre eux tuent les petits des espèces parasitées et sont nourris par leurs hôtes.
Les femelles de Coucou gris, par exemple, pondent un œuf directement dans le nid de différentes espèces de passereaux et ceux-ci l'élèvent souvent aux dépens de leur propre couvée. Ce coucou apprend même le chant de ses parents adoptifs. Le Coucou geai parasite presque exclusivement la Pie bavarde. Ce comportement existe également chez plusieurs canards comme la Nette rousse qui pond généralement dans son propre nid mais dépose aussi fréquemment des œufs dans les nids d'autres espèces apparentées, en particulier le Fuligule morillon[119] - [120] - [121].
Ce type de parasitisme, même chez la Fuligule à dos blanc de la part de la Fuligule à tête rouge où le but du parasitisme se limite à l'utilisation du nid, peut avoir un effet négatif significatif à plus ou moins long terme, même dans le cas où les poussins supplémentaires ne tuent pas ou ne concurrencent pas alimentairement les poussins de la portée[122].
Prédation
Les oiseaux adultes des plus grandes espèces ont peu de prédateurs exception faite des hommes, d'autres espèces d'oiseaux mais aussi des félins, des Mustelinae et de certains Viverridae.
D'autres espèces d'oiseaux comme le Xénique de Stephen ont disparu ou sont gravement menacées par la prédation d'animaux domestiques comme le chat. Mais ce sont les œufs et les oisillons qui sont les plus vulnérables et qui subissent une prédation importante.
Certains oiseaux sécrètent des composants chimiques contre leurs prédateurs, par exemple les Procellariiformes sont capables d'éjecter une graisse d'odeur désagréable[123].
Il existe par ailleurs quelques oiseaux vénéneux. On trouve par exemple de la batrachotoxine dans la peau et les plumes de l'Ifrita de Kowald[124] et de quelques espèces de Pitohui[124] - [125] - [126], tous de Nouvelle-Guinée.
Commensalisme des oiseaux

Certains oiseaux comme les piquebœufs s'alimentent en débarrassant les gros mammifères ou les reptiles de leurs parasites. Certains oiseaux aquatiques se nourrissent parfois en profitant des déplacements de proies occasionnés par d'autres espèces s'alimentant. De tels comportements commensaux ont été observés en Europe chez le Grèbe castagneux[127], notamment en France, vis-à-vis du Cygne tuberculé et du Canard chipeau, et à Madagascar vis-à-vis de l'Érismature à dos blanc, du Dendrocygne veuf, de l'Anserelle naine et du Canard à bec rouge[128]. Ils ont également été notés chez l'Aigrette garzette, la Grande aigrette et le Héron cendré vis-à-vis du Grand cormoran[129].
Chez le Puffin de Parkinson, ce commensalisme alimentaire est semble-t-il obligatoire et s'exerce aussi bien sur d'autres espèces d'oiseaux de mer que sur des cétacés comme les dauphins[130].
Certaines espèces d'oiseaux granivores ou frugivores favorisent la dissémination des graines d'espèces végétales, comme le gui par la Grive draine, et d'autres, comme les oiseaux-mouches, sont les seules à pouvoir polliniser certaines fleurs[131].
Bon nombre d'espèces, par leurs cris d'alarme, préviennent les proies potentielles de leur espèce et d'autres de l'approche d'un prédateur.
Répartition
Zones de répartition
Les oiseaux constituent un des groupes les plus homogènes du vivant, leur architecture est en effet conditionnée par de rigoureuses adaptations au vol. On en répertorie en 2020 autour de 11 430 espèces[3]. Leur répartition est mondiale et dépend des saisons car de nombreuses espèces sont migratrices. La diversité la plus élevée se trouve dans les régions tropicales, en domaine continental, la moins élevée se situe au niveau des régions polaires.
Bon nombre d'espèces sont grégaires, les populations sont donc concentrées sur certains sites, principalement sous les latitudes élevées. Ceci peut être dû à une spéciation ou à une extinction plus élevée dans ces régions[132].

Certains oiseaux comme les Pétrels des neiges sont capables de s'aventurer jusqu'à 400 km en Antarctique[133], plus loin qu'aucun autre vertébré. Certains manchots peuvent plonger à plus de 450 mètres[134]. Plusieurs espèces d'oiseaux marins se sont si bien adaptées aux milieux marins qu'elles ne regagnent la terre que pour la nidification[135].
Il semble que l'on puisse répartir les familles d'oiseaux en six groupes en fonction de leurs origines géographiques[136] : la zone pantropicale, la zone panboréale ou holarctique, la zone pan-américaine devenue la zone néotropicale, la zone paléarctique, la zone néarctique, et le dernier étant un groupe formé à partir des oiseaux très mobiles. La dérive des continents a joué un rôle évident dans l'endémisme de certaines familles, par exemple les oiseaux-mouches ne sont présents qu'en Amérique tandis que les émeus ne sont présents qu'en Australie. La zone australasienne est, comme pour la classe des mammifères, l'écozone qui possède le plus fort taux d'endémisme avec 16 familles sur 64 et par exemple 308 espèces de passereaux sur 385[137].
| écozone | Néarctique | Paléarctique | Afrotropical | Néotropique | Australasien | Océanien | Indomalais | Antarctique |
|---|---|---|---|---|---|---|---|---|
| familles[alpha 6] | 750 | 1025 | 1550 | 3700 | 900 | 960 | ||
| espèces[alpha 6] | 69 | 65 | 30 | 64 | 66 | |||
| Statut migratoire | Nombreux migrateurs | Migratoire pour la plupart | résidents | Migrateur à 40 % env[138]. | résidents | |||
| Liste d'espèces par pays ou région |
Voir aussi : La liste par pays
Évolution des répartitions
La principale cause naturelle d'expansion de l'aire de répartition d'un oiseau est la recherche de nourriture. Elle peut se faire soit au hasard d'une migration erronée, soit d'une façon lente par adaptation.
L'extension des aires de répartition pour certaines espèces au détriment d'autres croît fortement depuis quelques années et elle est probablement due à l'action de l'homme sur l'environnement, comme le réchauffement climatique ou la perturbation des habitats d'origine. Ces changements d'aire ont toujours existé et provoquent à long terme des spéciations, et par conséquent ils sont un processus normal de l'évolution.

Cependant, par l'accélération de ce phénomène, les changements d'aire témoignent d'un changement global de l'environnement. Outre le réchauffement, l'urbanisation et la déforestation sont aussi en cause. En Amérique du Nord, on a observé que Scardafella inca est peu à peu remplacée par la Tourterelle triste, plus adaptée à la présence de l'homme[139]. Certaines espèces sont devenues invasives après introduction directe par les humains, par exemple les pigeons bisets, les étourneaux sansonnets ou les martins tristes en Australie. Ces espèces ont été introduites pour lutter contre les criquets. Le Faisan de Colchide a été introduit dans de nombreux endroits pour la chasse[140]. D'autres, comme la Conure veuve, oiseau domestique, se sont simplement échappés. On en retrouve des colonies installées en Amérique du Nord[141], en Espagne ou en Belgique. Quelques espèces sauvages comme le Héron garde-bœufs[142], Milvago chimachima[143], le Cacatoès rosalbin[144] se sont étendues au-delà de leurs aires naturelles à la suite de l'extension de certaines pratiques agricoles.
Enfin les espèces domestiquées se sont répandues dans le cadre des élevages humains comme le Coq doré devenu coq domestique. Il est à noter que le moineau domestique qui n'est domestique que dans la mesure où il a suivi l'homme partout, vit des activités humaines.
Ces espèces colonisatrices, par leurs stratégies reproductives ou alimentaires, concurrencent et finissent par supplanter les espèces autochtones, entraînant à l'échelle humaine une baisse de la diversité génétique. Globalement, les populations d'oiseaux sont en déclins.
Classification
Liste des ordres actuels
Liste des ordres d'oiseaux modernes selon BioLib (26 septembre 2020)[145] et d'après la classification du Congrès ornithologique international (version 5.1, 2015):
- ordre Aberratiodontuiformes Enpu Gong, Lianhai Hou & Lixia Wang, 2004 †
- ordre Accipitriformes Vieillot, 1816
- ordre Aepyornithiformes Newton, 1884 †
- ordre Alexornithiformes Brodkorb, 1976 †
- ordre Ambiortiformes Kuročkin, 1982 †
- ordre Anseriformes Wagler, 1831
- ordre Apatornithiformes †
- ordre Apodiformes Peters, 1940
- ordre Apterygiformes Haeckel, 1866
- ordre Archaeopterygiformes Lambrecht, 1933 †
- ordre Bucerotiformes Fürbringer, 1888
- ordre Caprimulgiformes Ridgway, 1881
- ordre Cariamiformes Fürbringer, 1888
- ordre Casuariiformes Sclater, 1880
- ordre Cathartiformes Coues, 1884
- ordre Cathayornithiformes Zhou, Jin & Zhang, 1992 †
- ordre Chaoyangiaformes †
- ordre Charadriiformes Huxley, 1867
- ordre Ciconiiformes Bonaparte, 1854
- ordre Coliiformes
- ordre Columbiformes
- ordre Confuciusornithiformes Hou & al., 1995 †
- ordre Coraciiformes Forbes, 1884
- ordre Cuculiformes
- ordre Dinornithiformes Gadow, 1893 †
- ordre Enantiornithiformes Chiappe, 1992 †
- ordre Eurolimnornithiformes Bock & Buhler, 1996 †
- ordre Eurypygiformes Hackett & al., 2008
- ordre Falconiformes Sharpe, 1874
- ordre Galbuliformes
- ordre Galliformes Temminck, 1820
- ordre Gansuiformes Lianhai Hou & Zhicheng Liu, 1984 †
- ordre Gastornithiformes †
- ordre Gaviiformes Wetmore & Miller, 1926
- ordre Gobipterygiformes Elzanowski, 1974 †
- ordre Gruiformes Bonaparte, 1854
- ordre Hesperornithiformes Fürbringer, 1888 †
- ordre Iberomesornithiformes Sanz & Bonaparte, 1992 †
- ordre Ichthyornithiformes †
- ordre Leptosomiformes Sharpe, 1991
- ordre Lithornithiformes Houde, 1988 †
- ordre Mesitornithiformes Wetmore, 1960
- ordre Musophagiformes Seebohm, 1890
- ordre Opisthocomiformes
- ordre Otidiformes Wagler, 1830
- ordre Palaeocursornithiformes Kessler & Jurčák, 1986 †
- ordre Passeriformes Linnaeus, 1758
- ordre Patagopterygiformes Alvarenga & J.F. Bonaparte, 1992 †
- ordre Pelecaniformes Sharpe, 1891
- ordre Phaethontiformes Sharpe, 1891
- ordre Phoenicopteriformes Fürbringer, 1888
- ordre Piciformes Meyer & Wolf, 1810
- ordre Podicipediformes
- ordre Procellariiformes
- ordre Psittaciformes
- ordre Pteroclidiformes
- ordre Remiornithiformes Martin, 1992 †
- ordre Rheiformes Forbes, 1884
- ordre Sandcoleiformes Houde & Olson, 1992 †
- ordre Sinornithiformes Sereno & Rao, 1992 †
- ordre Sphenisciformes
- ordre Strigiformes Wagler, 1830
- ordre Struthioniformes Latham, 1790
- ordre Suliformes Reichenbach, 1849
- ordre Tinamiformes Huxley, 1872
- ordre Trogoniformes
.JPG.webp)

 Colibri cyanotus (Apodiformes).
Colibri cyanotus (Apodiformes)..jpg.webp)





%252C_delta_del_Okavango%252C_Botsuana%252C_2018-07-31%252C_DD_11.jpg.webp)
.jpg.webp)


_by_N.A._Nazeer.jpg.webp)


_strutting.jpg.webp)



.jpg.webp)
%252C_parque_nacional_Kruger%252C_Sud%C3%A1frica%252C_2018-07-26%252C_DD_33.jpg.webp)
.jpg.webp)



.jpg.webp)

.jpg.webp)
_(14).JPG.webp)

.JPG.webp)
_male.jpg.webp)



%252C_cabo_de_Buena_Esparanza%252C_Sud%C3%A1frica%252C_2018-07-23%252C_DD_87.jpg.webp)


_female.jpg.webp)



.jpg.webp)
 Jeholornis prima (Jeholornithiformes).
Jeholornis prima (Jeholornithiformes). Lithornis (Lithornithiformes).
Lithornis (Lithornithiformes).
Phylogénie des ordres
Cladogramme illustrant la phylogénie des différents ordres d'oiseaux actuels d'après Jarvis, E.D. et al. (2014)[146] et Yury, T. et al. (2013)[147] :
| Neornithes |
| ||||||||||||||||||||||||||||||||||||
Histoire évolutive

 Modèle d'un Théropode, Compsognathus.
Modèle d'un Théropode, Compsognathus. Modèle d'un Archaeopteryx.
Modèle d'un Archaeopteryx.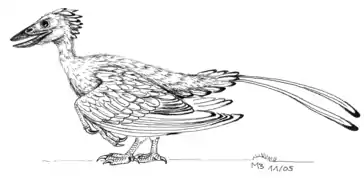 Modèle d'un Confuciusornis.
Modèle d'un Confuciusornis. modèle d'un Enianthornithe, Iberomesornis.
modèle d'un Enianthornithe, Iberomesornis.
L'étude de l'histoire évolutive des oiseaux est rendue difficile du fait que les fossiles d'oiseaux en bon état sont difficiles à trouver. Cependant, les plus anciens fossiles considérés comme appartenant à cette classe remontent au Jurassique supérieur, il y a plus de 150 Ma. Si l'on veut supposer les dinosaures comme un clade monophylétique, il faut considérer les oiseaux comme seuls représentants actuels des dinosaures. Ils sont issus du groupe des Théropodes[151] (dans lequel on trouve le Tyrannosaure ou encore le Vélociraptor).
Cette classe a connu une première spéciation principalement en deux grandes branches qui allaient survivre à l'extinction du Trias-Jurassique, les Enantiornithomorpha et les Ornithurae.
Les Ornithurae et plus particulièrement les Neornithes vont connaître une importante radiation évolutive. De nombreuses espèces vont disparaître à l'extinction du Crétacé, y compris l'ensemble des Enantiornithomorpha. En fait, seule la branche issue des Neornithes, les super-ordres des Paleognathae et des Neognathes, dont les Anseriformes, Galliformes et Neoaves, vont survivre[151].
Les oiseaux vont continuer à évoluer et à se différencier tout au long du cénozoïque (depuis 65 Ma). Ainsi les Craciformes sont issus de la branche des Galliformes ou les Passeriformes sont issus des Neoaves. On considère aujourd'hui 27 ordres différents.
| Caractéristiques anatomiques | Théropodes (non aviens) 220 à 65 Ma |
Archaeopteryx 156 à 150 Ma |
Confuciusornis 125 et 110 Ma |
oiseaux plus développés Ornithothoraces à partir du Jurassique moyen |
|---|---|---|---|---|
| Fusion des deux fenêtres temporales avec l'orbite | - | + | - | + |
| Disparition des dents | - nombreuses | - | + | Beaucoup pour les Oiseaux du Crétacé |
| La queue à Pygostyle raccourcie, vertèbres de queue fusionnées | en petit nombre | - | + | + |
| Clavicules fusionnées (furcula)| | - Accolées (forme de boomerang) | |
+ Forme de boomerang |
+ Forme de boomerang |
+ Forme en V, à angle aigu |
| longueur du coracoïdes long, en forme de ruban | - | - | + | + |
| Sternum avec bréchet | - | - | - Sternum plat avec quille très réduite |
+ Seulement les Neognathae, disparition secondaire chez les paléognathes |
| Côtes liées au sternum | avec une | - | + | + |
| Disparition des côtes abdominales (Gastralia) | - | - | - | + |
| côtes avec processus unciné | avec une côte supplémentaire ou présent chez les Maniraptora | - | + | + Plusieurs |
| Humérus dilaté avec crête deltopectorale perforée (autapomorphie) | - | - | + | - |
| os de métacarpe des pattes avant I, III à carpe-métacarpe fusionné | - | - | + (fusion après la base) |
+ |
| os médian du pied II-IV fusionné (tarso-métatarse) | un | + | + | + |
| os médian du pied IV présent | + | + | + | - |
| Plumes de vol actif, asymétriques | quelques-uns | + | + | + |
| Alula | peu abondantes | - | - | + |
Relations entre les oiseaux et l'homme
Élevage de volailles

Les oiseaux élevés pour la consommation sont appelés volailles. La volaille la plus consommée est le poulet domestique. L'espèce ancestrale nommée Gallus gallus a été domestiquée entre le VIe et le IIIe millénaire av. J.-C. et de nombreuses races ont été sélectionnées. Depuis, l'élevage des volailles n'a pas cessé d'être un enjeu économique important. La production mondiale de volaille a décuplé avec la transformation de l'aviculture par la méthode des élevages en batterie. La production de poulets domestiques a été multipliée par quatre entre 1964 et 2004[152], celle des autres volailles usuelles dans une proportion au moins égale. En 2004, la production de l'ensemble des autres volailles ne dépasse pas, en volume, 10 % de celle des poulets[152] et représente au total environ 81 millions de tonnes en 2005.
Les volailles élevées en fonction des caractéristiques de leur race sont dites de chair ou pondeuses. Certains éleveurs se sont spécialisés dans la production de poussins, d'autres en engraissement ou dans la production d'œufs. De ce type d'élevage résultent plusieurs controverses qui concernent par exemple le bien-être animal, la production de lisier polluant en trop grande quantité, la menace sur la diversité biologique des volailles par la réduction des races, l'introduction de résistance aux antibiotiques par une utilisation non raisonnée de ceux-ci, une sécurité alimentaire hasardeuse du fait à la fois de la mauvaise nutrition des oiseaux, de la multiplication des zoonoses et de la diffusion de zoonose par le transport.
À la stricte production des volailles sont associées une industrie spécialisée dans l'abattage, une industrie de transformation en aliment consommable, une industrie de transformation des produits dérivés et de conditionnement, une industrie de fabrication des aliments pour les volailles, une industrie du transport, ce qui fait de ce secteur agricole un enjeu industriel majeur. L'enjeu éclipse les problèmes posés.
Certaines espèces sauvages sont aujourd'hui domestiquées et élevées pour leur viande, leurs œufs et leurs plumes, comme les autruches ou les cailles.
Chasse et pêche

La chasse aux oiseaux, oiseaux qualifiés dans ce contexte de gibier, a été historiquement un enjeu alimentaire important[153], ce qui a même mené des dizaines d'espèces à l'extinction ou au bord de l'extinction[154].
Aujourd'hui, la chasse aux oiseaux n'est plus une nécessité alimentaire dans les pays développés, mais elle est considérée soit comme une distraction soit comme un sport.
Certaines chasses traditionnelles utilisent un oiseau pour la capture du gibier, comme la fauconnerie. En Asie du Sud-Est, des cormorans étaient autrefois utilisés pour la pêche[155]. En Occident, les chasseurs préfèrent utiliser des chiens pour débusquer les oiseaux et ensuite les tirer au fusil. Les chasses au collet et à la glu ont également été utilisées mais sont aujourd'hui interdites. En France, la chasse de certaines espèces est très controversée, comme la chasse à la palombe : les pratiques de chasse aux oiseaux y sont souvent en contradiction avec les lois européennes.
La faune aviaire devenant rare du fait des pratiques de chasses ou agricoles, certaines associations de chasse organisent des élevages d'oiseaux à l'origine sauvages, comme des faisans ou des cailles, pour être relâchés afin que les membres de ces associations aient des cibles pendant la période de chasse. Ces oiseaux sont souvent des hybrides stériles. Les observateurs remarquent qu'ils n'ont pas le comportement d'oiseaux nés en liberté.
Rôles dans les sociétés humaines

D'autres oiseaux d'élevages peuvent également être utilisés à d'autres fins qu'alimentaires. Diverses races domestiquées à partir du Pigeon biset sont utilisées comme messagers ou comme aliments. Ils ont joué, comme messagers, un rôle important durant la Seconde Guerre mondiale. Un de ces pigeons, « Cher Ami », a même été décoré de la Croix de Guerre américaine[156]. Aujourd'hui ces pratiques ont changé. La colombophilie est devenue une activité ludique.
Les coqs de combat sont sources de paris ; dans certains pays comme l'Afrique du Sud ou les États-Unis, on organise des courses d'autruches[157].

Plusieurs espèces d'oiseaux sont capturées et enfermées pour distraire l'homme, soit par leurs chants, soit pour leurs plumes. L'élevage d'oiseaux comme animaux de compagnie s'est généralisé.
La fauconnerie s'est transformée presque partout avec d'autres objectifs que la chasse, par exemple des rapaces sont des centres d'attraction destinés à faire venir les touristes dans certains parcs à thèmes comme le château des Milandes[158], le château de Fauquemont[159], le château de Bouillon[160], etc.
L'engouement pour les oiseaux a créé un tourisme spécialisé qui consiste en voyages d'observations. Des aménagements ont été réalisés sur certains sites pour faciliter ces observations. Pour les particuliers, un secteur devenu financièrement important s'est spécialisé dans la nourriture des oiseaux sauvages ou la construction de nichoirs. Une étude a estimé qu'au Royaume-Uni 75 % des ménages nourrissaient les oiseaux[161].
Les plumes d'oiseaux sont utilisées pour la confection de vêtements, de duvets et oreillers, etc. Les eiders ou les oies sont réputés pour leur duvet par exemple. Ces pratiques ont tendance à diminuer en ce qui concerne l'utilisation décorative des plumes.
Le guano, issu des excréments des oiseaux de mer, est utilisé comme engrais et est très recherché, son exploitation a été source de revenus considérables pour le Pérou au XIXe siècle.
D'autre part, plusieurs types de commensalisme entre oiseaux et humains sont connus : par exemple, les indicateurs sont une famille d'oiseaux utiles aux humains pour leur permettre trouver des ruches sauvages.
Rôle écologique des oiseaux
Le rôle et l'impact des oiseaux sur l'écologie est un sujet de plus en plus soumis à l'étude. Ils jouent notamment un rôle important dans la lutte contre les nuisibles. On retrouve par exemple la mésange bleue (Cyanistes caeruleus) ou encore la mésange charbonnière (Parus major), prédatrices efficaces des insectes et des larves dans les jardins et les vergers. La chouette effraie (Tyto alba) se révèle également être une redoutable alliée des fermiers du Moyen-Orient, où elle est utilisée pour contrôler les populations de rongeurs, véritable fléau pour les cultures.
Utilisations commerciales de leur image
Dans la publicité, comme symboles d'entreprise ou comme mascottes, outre les oiseaux de fiction célèbres, les images d'oiseaux sont très utilisées d'une manière plus ou moins stylisée (cf. Oiseau postal). Aux États-Unis, l'image des rapaces prédateurs, donc sans les vautours, est largement utilisée, par exemple par l'équipe de football américain des Seahawks de Seattle. La chouette, symbole d'Athéna, déesse de la sagesse dans la Grèce antique, est utilisée par diverses banques et compagnies d'assurances et est un symbole utilisé par l'État grec.
Les oiseaux sont également très représentés sur les timbres postaux.
Oiseaux dans la culture humaine

Les oiseaux ou leurs œufs, de par leur répartition, sont connus de toutes les cultures humaines et ceci a directement influencé leurs coutumes, leurs religions et leur vie, par le symbolisme qui leur est associé.
Symbolique de l'oiseau
.svg.png.webp)
On leur a donc attribué, comme à tous les animaux familiers à l'homme, des caractéristiques plus ou moins anthropomorphiques suivant les espèces mais aussi d'une manière générale. L'oiseau ou ses caractéristiques associées (comme l'œuf, les plumes, les ailes ou les serres) peuvent être considérés comme symbole. Le vol de l'oiseau est naturellement porteur d'un symbole de liberté comme l'exprime le mythe grec d'Icare. Par exemple, l'aigle est un symbole de majesté, c'est pourquoi on le retrouve sur les armoiries de divers empires. La paix est symbolisée par la colombe.
La plupart des oiseaux sont considérés comme symboles positifs, mais il existe cependant des exceptions en fonction des cultures. Les vautours ont une réputation négative pour beaucoup de peuples, cependant les parsis, et les zoroastriens avant eux, donnent à ces oiseaux un rôle singulier. Ils offrent le cadavre de leurs morts aux vautours, l'inhumation ou la crémation étant une offense à la terre et au feu. Au Moyen Âge en Occident, les rapaces, surtout les nocturnes comme les Strigiformes (chouettes, hiboux), bien que très bénéfiques aux paysans car mangeant des espèces ravageuses comme les petits rongeurs, étaient jugés comme maléfiques ou nuisibles. Une coutume consistait à clouer sur les portes ces oiseaux pour chasser les mauvais esprits et tous les autres Strigiformes. D'ailleurs plusieurs espèces de rapaces ont été pourchassées. Ces animaux sont aujourd'hui protégés et leur image actuelle de majesté est la plus forte, bien que les vautours, avec leur aspect ingrat, soient toujours mal acceptés[162].
En Afrique, certains chants et cris sont interprétés et traduits par certains peuples qui en tirent une signification particulière, qu'elle soit positive ou néfaste. Les oiseaux produisant ces cris sont alors considérés comme bénéfiques ou maléfiques. L'art martial d'oiseau imite l'agilité de l'oiseau pour stimuler les poumons : Il travaille sur l'amplitude de la respiration, et sur le relâchement des articulations des membres supérieurs.
Oiseaux dans la langue française
En français, par exemple, chanter comme un oiseau signifie chanter gaiement. Avoir un appétit d'oiseau signifie manger très peu. Une cervelle ou tête d'oiseau est un esprit léger, insouciant, étourdi, et un individu qui vit comme un oiseau, le fait sans souci du lendemain. Être libre comme un oiseau signifie n'avoir aucune entrave. Se faire donner des noms d'oiseaux signifie se faire copieusement insulter. Dans le langage populaire, un oiseau désigne un individu, avec un sens plutôt négatif. Un oiseau de passage désigne un individu dont on n'entendra plus parler une fois parti, un oiseau de mauvais augure est un porteur de mauvaises nouvelles. Un oiseau de nuit est un noctambule. Un oiseau rare est une personne que l'on cherche à contacter mais qui se rend peu disponible.
Bon nombre d'expressions sont liées à des espèces précises en fonction des symboles particuliers liés aux espèces considérées : faire le paon pour faire le beau, être un pigeon pour être un naïf, une oie pour une jeune femme naïve, être une mère poule pour une mère très attentionnée. Les termes vautour, corbeau, rapace et ainsi que ceux partagés par d'autres charognards ne sont pas des qualificatifs plaisants.
L'utilisation du mot « oiseau » et du nom des oiseaux pour d'autres usages qu'une dénomination simple est très fréquente. De nombreuses œuvres artistiques font référence, dans leur titre, au terme « oiseau » comme L'Oiseau de feu, L'Oiseau et l'Enfant, L'Oiseau au plumage de cristal, Les Oiseaux, L'Oiseau bleu, Le Roi et l'Oiseau, L'Oiseau d'argile, L'Oiseau d'Amérique, L'Expérience avec l'oiseau dans la pompe à air sans qu'il y ait nécessairement un rapport direct avec les oiseaux. D'autres œuvres font naturellement référence à des espèces particulières, par exemple dans L'Affaire Pélican ou, d'une manière très indirecte, dans Le Faucon maltais ou dans Bécassine, la bécassine étant un oiseau réputé stupide. Il peut aussi n'y avoir aucun rapport avec les oiseaux, comme le film Les Oies sauvages. Plusieurs plantes comportent aussi ce terme, par exemple la Néottie nid d'oiseau, l'Oiseau de paradis. On retrouve enfin ce terme dans des lieux comme la commune de Champ-d'Oiseau ou pour des objets manufacturés divers qui vont de l'Oiseau buveur à l'Oiseau de Feu.
Oiseaux de fiction
Dans la littérature et la poésie, nombre d'œuvres prennent l'oiseau pour sujet, par exemple Chantecler, d'Edmond Rostand ou L'Albatros de Charles Baudelaire
Oiseaux mythiques

Parmi les oiseaux mythiques, on peut citer le Rokh des Mille et Une Nuits et de Sinbad le marin, qui est un rapace géant capable de capturer des bateaux et des hommes.
Le Pouākai, selon les légendes māori, était capable de capturer un humain. Cet oiseau mythique étant probablement la sublimation de l'aigle géant de Haast, disparu avec l'extermination des moas par ces mêmes Māori[163].
Le Sphinx grec, la harpie, Pégase, les sirènes, les chimères sont des créatures hybrides possédant des traits d'oiseaux et sont plutôt hostiles à l'homme.
Le Phénix, symbole de la renaissance est une créature mythique qui a volontiers été utilisée par les chrétiens qui y voient un symbole de la résurrection de Jésus-Christ.
Garuda est un oiseau géant de la mythologie indienne, incarnation de Vishnu.
Le Simurgh, oiseau de la mythologie perse, est également présent dans l'Avesta ou le Shahnameh.
Le Nok thet, oiseau fantastique issu de la mythologie hindouiste.
Contes et légendes
Il existe de très nombreux contes et légendes mettant en scène des oiseaux[164]. Les fables de Jean de La Fontaine par exemple, mettent en scène des personnages zoomorphes comme Le Corbeau et le Renard, La Colombe et la Fourmi ou Le Renard et la Cigogne.
En Occident, dans certaines régions, pour cacher l'existence de la sexualité aux enfants, on raconte que les bébés sont apportés aux parents par les cigognes, faisant de cet animal un symbole positif. Cette légende a probablement sauvé de l'extinction les populations de ces espèces dans l'est de la France.
La Poule aux œufs d'or est à la fois un mythe sur la chance et une allégorie sur l'importance économique de la volaille.
Animations

En Occident, il existe des centaines de dessins animés ou bandes dessinées comportant des oiseaux comme personnages principaux et secondaires. Ce sont des caricatures auxquelles on attribue un caractère que l'on prête à ces oiseaux, ainsi, par exemple, la poule est mère poule ou frivole idiote, le coq est hardi et arrogant comme dans les dessins animés de Charlie le coq produits par Looney Tunes, même si la graphie de certains longs métrages, comme Birds in the Spring, est plus réaliste. Orville, l'albatros de Bernard et Bianca, n'a pas simplement une démarche comique, il l'est. Les petits oiseaux sont gentils ou malins comme le canari Titi de Looney Tunes. Souvent, les canards sont colériques, bruyants et roublards comme Donald Duck de Disney et Daffy Duck de Looney Tunes. Les rapaces sont en général des méchants, comme Beaky Buzzard, Hennery le faucon ou plus récemment le faucon de Stuart Little 2, et s'affrontent à de gentilles souris ou autres oiseaux comme Woody Woodpecker. Face aux héros de ces œuvres manichéennes, les méchants finissent toujours bredouilles. Parmi ces oiseaux célèbres, on peut citer également Woodstock (en), le meilleur ami de Snoopy, Chilly Willy le manchot, Yakky Doodle (en) le pigeon de Hanna-Barbera et Bip Bip, le Grand géocoucou de Bip Bip et Coyote.
Dans les dessins animés plus provocateurs, ces stéréotypes sont moins vrais comme dans Cléo et Chico, les personnages étant des anti-héros. À travers ces dessins animés, certains types de relations humains-oiseaux peuvent être évoqués, par exemple l'élevage en batterie dans Chicken Run et être ainsi dénoncés.
Cinéma
Le cinéma est riche de situations où les oiseaux sont montrés, évoqués, ou même au centre de l'action. Une œuvre qui a particulièrement marqué les esprits du XXe siècle est celle d’Alfred Hitchcock, Les Oiseaux, tirée de la nouvelle éponyme de Daphné du Maurier qui présente les oiseaux comme une menace, sous forme de nuées dangereuses. Birdy, film d'Alan Parker, est une œuvre où un homme se prend pour un oiseau.
Plusieurs films documentaires portant sur les oiseaux, sortis au cinéma, ont été réalisés, comme La Marche de l'Empereur de Luc Jacquet ou Le Peuple Migrateur de Jacques Perrin.
Oiseaux dans l’art

Les oiseaux apparaissent dans la culture et l'art pariétal depuis la préhistoire où ils sont représentés sur des peintures rupestres[165] - [166], par exemple dans la scène du puits de la grotte de Lascaux. Homère s'est servi des oiseaux dans son œuvre et notamment du Rossignol dans l'Odyssée. Il fut suivi par de nombreux poètes, du fait de l'élégance et de la complexité du chant de cet oiseau[167].
Dans l'art médiéval aussi, le poète soufi iranien Farīd ad-Dīn ʿAṭṭār a écrit l'histoire d'une bande de trente oiseaux pèlerins partant sous la conduite de la huppe à la recherche du Simorgh dans son livre La Conférence des oiseaux en 1177. À la fin de leur quête, ils découvrent leur moi profond (jeu de mots sur Simurgh = 30 oiseaux).
Les oiseaux n'ont pas cessé d'inspirer les artistes par leurs formes, leurs couleurs et les symbolismes auxquels ils sont associés, comme le trône du Paon. Certains artistes se sont spécialisés avec succès sur les oiseaux comme Jean-Jacques Audubon qui allait donner son nom à la Société nationale Audubon. Chez Ximena Armas, l’oiseau est la victime première d’une nature outragée ; il n’est souvent présent que par quelques plumes éparses.
La relation entre les albatros et les marins, thème central de La Complainte du vieux marin de Samuel Taylor Coleridge, est un exemple d'allégorie poétique. Les oiseaux ont inspiré de nombreux poèmes, comme L'Albatros de Charles Baudelaire dans Les Fleurs du mal ou La mort de l'aigle de José-Maria de Heredia dans le recueil Les Trophées[168].
Oiseaux célèbres
Quelques oiseaux sont devenus célèbres comme Alex et William d'Orange.
Religion
Si plusieurs religions ont directement utilisé certains oiseaux comme symbole religieux, la plupart ont utilisé l'œuf, les plumes ou les ailes. Certaines religions considèrent certains oiseaux comme porteurs d'une essence divine car symbole de leur(s) dieu(x). Par exemple, à l'instar de plusieurs autres types d'animaux, la religion égyptienne antique élevait avec un grand faste et respect les spécimens de Faucon, d'Ibis, de Huppe fasciée dans leur temple.
Certaines populations dravidiennes considèrent les paons bleus comme divins, manifestation de la Terre mère[169].
D'autres peuples, plus simplement, représentent leur(s) dieu(x) sous la forme d'oiseau, comme les yézédis qui représentent Melek Ta'us sous la forme d'un paon, comme les chrétiens utilisent la Colombe comme manifestation d'une des essences de la Sainte Trinité, le Saint-Esprit. La Bible se sert aussi de la colombe avec plusieurs significations associées, effroi, passivité, deuil et beauté. La couleur de la « blanche colombe » en fait un symbole de pureté qui était très commune au Moyen-Orient durant l'antiquité[170].
Les interprétations humaines de ces messages ne sont pas uniformes puisque la Huppe fasciée est sacrée en Égypte antique, elle est symbole de vertu en Perse et elle est signe de guerre dans les pays scandinaves[171].

Certains cultes ou mythologies ont attribué des pouvoirs à certains oiseaux, sans en faire des dieux, comme le rôle de messager des dieux attribué à Hugin et Munin, deux grands corbeaux dans les mythologies nordiques[172].
Ils peuvent également être simplement instrument sans conscience d'un dieu. La Bible, qui reprend le récit de l'Épopée de Gilgamesh, raconte, dans le Livre de la Genèse, que Noé lâche une colombe puis un corbeau pour trouver une terre après le déluge[173]. Dans le Livre des Rois, des corbeaux sont envoyés par Dieu pour nourrir Élie[174]. Dans le Coran, un corbeau envoyé par Allah apprend à Caïn comment enterrer son frère.
Ils peuvent faire office de messagers des dieux pour les prêtres comme dans la mythologie Rapa Nui (en) où Tangata manu, l'Homme-oiseau, le roi de l'Île de Pâques, était désigné par une course à la découverte d'un œuf. Ceci était vraisemblablement le cas pour toutes les cultures méditerranéennes antiques. Philon d'Alexandrie, un philosophe juif hellénisé, reconnaît ce rôle de messager aux oiseaux[175]. Dans la religion romaine par exemple, le vol des oiseaux ou leurs entrailles étaient interprétés par les haruspices et augures pour déterminer la volonté des dieux.
Également, pour certains cultes, les oiseaux peuvent être offerts en sacrifice. Le Lévitique précise que la colombe et les pigeons sont les seuls oiseaux offrables en sacrifice, le riche, lui, pouvant sacrifier du bétail.
Si les oiseaux adultes sont utilisés comme symbole religieux, il en est de même pour les œufs, symboles par excellence de la renaissance et de la pureté des formes, mais aussi pour certains de leurs comportements. Le vol des oiseaux est considéré comme un signe de la perfection divine dans le Coran[176].
Les oiseaux sont perçus plutôt comme des symboles bénéfiques, une illustration possible de ce fait est qu'en iconographie occidentale les ailes des anges, créatures bénéfiques, sont représentées sous forme d'ailes d'oiseaux, animaux diurnes, alors que celles des démons, créatures maléfiques, sont représentées avec des ailes de chiroptères, animaux nocturnes.
Étude des oiseaux
La discipline qui étudie les oiseaux se nomme l'ornithologie.
Vous pouvez consulter ici une liste d'ornithologues.
Différentes sciences étudiant les oiseaux

L'étude des oiseaux est une science pratiquée par une large majorité d'amateurs et non par les seuls ornithologues. Leur observation et la collecte d'informations relèvent souvent de techniques simples nécessitant peu de matériel. Les scientifiques « officiels » (peu nombreux dans la majorité des pays) s'appuient sur des réseaux d'observateurs amateurs, parfois très étoffés (deux millions de Britanniques adhèrent à la Royal Society for the Protection of Birds).
On distingue usuellement plusieurs branches à l'ornithologie, par exemple l'Oologie pour l'étude des œufs, la Paléozoologie pour l'étude des œufs fossiles. L'Ethnoornithologie étudie, elle, les rapports entre les hommes et les oiseaux.
L'ornithologie scientifique s'appuie aujourd'hui sur la systématique, l'éthologie et la psychologie comparative, la biologie moléculaire, la génétique, la dynamique des populations, mais aussi la paléontologie (pour comprendre la phylogénétique des oiseaux). Les pinsons de Darwin sont par exemple à l'origine de notre compréhension du phénomène de spéciation et des mécanismes de l'évolution des espèces.
Elle a notamment montré que de nombreuses espèces étaient menacées (en particulier par la destruction de leur habitat et la généralisation de l'utilisation des pesticides et avicides) et que les oiseaux situés au sommet des chaînes alimentaires sont des indicateurs très sensibles de la pollution et de l'état des écosystèmes. Les oiseaux communs ne sont pas épargnés (à titre d'exemple, selon le programme STOC, de 1989 à 2007, leurs populations ont globalement décliné de 18 % en France).
Oiseaux de laboratoire
La classe des oiseaux est une des classes les plus étudiées en laboratoire. De nombreuses espèces y sont étudiées du fait de la facilité des conditions d'élevage. Certaines espèces sont plus étudiées pour leurs vocalisations comme les diamants mandarins (espèce ayant également fait l'objet d'études génétiques approfondies[177]), d'autres pour leur intelligence comme celle du genre Corvus. Les poulets et les pigeons sont également populaires pour la biologie et la psychologie comparative.
Désignations, classification
Initialement, la langue française disposait, pour désigner les oiseaux, de noms génériques basés uniquement sur la morphologie (rapace, échassier, canard, perroquet, hirondelle, oie, cygne, etc.) ou sur l'usage qu'on en fait (volaille, sauvagine...), certains de ces termes ne désignant pas les mêmes oiseaux d'une région à l'autre. Avec l'apparition de la systématique, les ornithologues ont créé des noms fondés sur l'anatomie pour faire des regroupements adaptés à un classement taxinomique. Ceux-ci sont passés dans le langage courant, mais beaucoup se sont avérés polyphylétiques, par exemple palmipède. Au cours du XXe siècle, la classification a évolué en regroupant les différentes espèces selon des affinités plus subtiles. Une autre révolution dans la taxinomie s'est produite au cours des années 1990, avec l'apparition d'une classification basée sur la génétique, en l'occurrence des techniques d'hybridation de l'ADN. Celle-ci est dans l'ensemble acceptée en Amérique du Nord, tandis que les réticences sont plus importantes ailleurs dans le monde, comme l'illustre la publication, entre 1992 et 2013, de l'encyclopédie des oiseaux Handbook of the Birds of the World (HBW) en 17 volumes, adoptant une classification qui ignore l'analyse génétique[178].
La nécessité de répertorier au niveau mondial les très nombreux oiseaux existant et les fossiles d'espèces disparues a nécessité rapidement la création de noms scientifiques. Un nom scientifique international a été attribué à chacun d'eux et le latin a été préféré pour cela.
Contrairement à la plupart des autres animaux, chacune des espèces d'oiseaux dispose d'un nom scientifique français normalisé unique attribué par la Commission internationale des noms français des oiseaux. Ces noms scientifiques normalisés en français n'ont pas le rayonnement mondial des noms scientifiques en latin. En français, les noms vernaculaires sont donc souvent accompagnés d'un nom binomial en latin, de portée mondiale, écrit en italique, et construit selon le Code international de nomenclature zoologique.
Histoire de la classification scientifique des oiseaux
Le naturaliste suisse Conrad Gessner publie en 1555 le troisième volume de son ouvrage de zoologie Historiae animalium, consacré aux oiseaux[179]. La même année paraît l'Histoire de la nature des oyseaux, avec leurs descriptions et naïfs portraicts retirez du naturel du naturaliste français Pierre Belon. Ces deux ouvrages constituent un premier effort de classification depuis Aristote dont ils s'inspirent. Celui de Gessner est plus érudit mais celui de Belon, plus original, propose la classification la plus intelligente et la plus solide pour l'époque[180]. Belon décrit tous les oiseaux qu'il connaît et les regroupe suivant leur comportement et leur anatomie : les oiseaux de proie, les oiseaux d'eaux, les omnivores, les petits oiseaux (subdivisés à leur tour en insectivores et en granivores)[181]. De nouveaux termes, forgés sur des racines latines, sont peu à peu créés et passent dans le langage populaire, comme palmipède. Au début du XVIIe siècle, le naturaliste italien Ulisse Aldrovandi fait paraître les trois livres constituant le volume XII de son Histoire naturelle. Son œuvre, comme celle de Gessner, lui vaudra de virulentes critiques, mais ses illustrations sont meilleures et sa classification plus évoluée[182].
La première classification scientifique est publiée en 1676 par Francis Willughby et John Ray sous le nom de Ornithologiae[183].

Petit à petit, surtout grâce aux travaux de Carl von Linné parus en 1758, cette classification va, cette classification va se perfectionner grâce (notamment) aux travaux sur l'anatomie comparée et l'embryologie.
La classification de Georges Cuvier marque une rupture avec les méthodes précédentes basées sur des critères anatomiques isolés, l'anatomiste français utilisant la taxonomie et l'anatomie comparée comme un outil et non comme une fin en soi[184]. Elle divise les oiseaux en six groupes : rapaces, passereaux, grimpeurs, gallinacés, échassiers, palmipèdes.
La classification la plus communément admise est ensuite celle de Sharpe dans le Catalogue of the Birds in the British Museum, qui propose les ordres des palmipèdes, échassiers, gallinacés, colombins ou pigeons, grimpeurs, passereaux, rapaces, coureurs. Les dénominations d'échassier ou palmipède sont aujourd'hui jugées comme obsolète d'un point de vue taxonomique mais continuent d'être utilisées[185].
Une des systématiques les plus importantes de la fin du XXe siècle est celle élaborée par James Lee Peters. La classification de Howard et Moore en dérive[186]. Dans la vision traditionnelle de l'évolution des oiseaux modernes (Neornithes), on place à la base de l'arbre phylogénétique, après les ratites et tinamous, les groupes d'oiseaux marins tels les manchots, les grèbes, les plongeons et pélicans, etc.
Vers la fin des années 1970 et durant toute la décennie suivante, Charles Gald Sibley et Jon Edward Ahlquist mènent des études fondées sur des méthodes d'hybridation de l'ADN, ce qui modifie profondément les connaissances sur la phylogénie des oiseaux[187]. La nouvelle classification (Phylogeny and classification of birds : a study in molecular evolution de Sibley et Ahlquist en 1990) montre que les canards et les gallinacés sont les parents les plus proches des ratites (qui intègrent les tinamous) formant les Paléognathes. Les Galloanserae constituent une lignée ancienne chez les Néognathes. Les groupes d'oiseaux marins traditionnellement considérés comme archaïques sont maintenant placés dans l'ordre des Ciconiiformes élargi, qui comprend aussi les rapaces diurnes (Accipitridae, Sagittariidae et Falconidae) et les limicoles (Scolopacidae, Charadriidae, Laridae, etc.). Dans cette nouvelle taxinomie du vivant, dite classification phylogénétique, les oiseaux font partie des Archosauriens qui comprennent un grand nombre de fossiles appelés dinosaures. Cette systématique[188] est dans l'ensemble acceptée en Amérique tandis que les réticences sont plus importantes en Europe.
Les recherches pour comprendre la position relative de chaque groupe d'oiseaux continuent et d'autres domaines que la génétique sont explorés. Un chercheur russe a démontré que la structure de la coquille des œufs pourrait être utile dans la détermination des relations entre oiseaux[189].
Maladies communiquées aux humains
Les oiseaux peuvent être porteurs, sains ou non, de maladies transmissibles à l'homme (voir le paragraphe « Maladies aviaires »). Les oiseaux peuvent entraîner également des allergies liées en particulier aux plumes.
Lutte contre les oiseaux


Certains oiseaux sont considérés comme nuisibles. Les causes en sont diverses et certaines très controversées. Par exemple, ils consomment de petits fruits ou du maïs, comme les carouges à épaulettes, les moineaux, certains étourneaux, les gros-becs, certaines corneilles et certains corbeaux[190]. Les pigeons sont quant à eux une nuisance pour les bâtiments, bien qu'en Angleterre on les voie attaquer certaines cultures.
Les moyens mis en place pour lutter contre eux peuvent être très différents.
L'abattage par certains chasseurs de rapaces particulièrement utiles aux agriculteurs, car chassant surtout les vermines mais perçus comme une concurrence déloyale pour la chasse aux lièvres et lapins, a fortement réduit certaines populations. Les vautours, dans le sud de la France, accusés sans preuves formelles de tuer moutons et vaches, ont été exterminés, avant d'y être réintroduits[191].
D'autres espèces, devenues invasives à la suite de l'introduction humaine car jugées initialement utiles, font l'objet de mesures visant soit la réduction de leurs populations soit l'éradication.
Pour les espèces sauvages autochtones, les agriculteurs cherchent avant tout à empêcher les oiseaux de consommer les plantes qu'ils cultivent. Les moyens mis en œuvre sont divers, répulsifs sonores, répulsifs visibles tels les épouvantails, chasse, destruction des nids, piégeage ou protection des cultures par filets ou sacs, par des produits chimiques répulsifs et par la culture d'espèces végétales répulsives[190].
Certains rapaces sont élevés pour effrayer les autres oiseaux autour des aéroports afin de protéger les avions des collisions. Ils sont également réintroduits dans les villes pour lutter contre les pigeons considérés comme des fléaux. En effet, ces derniers, en plus de transmettre certaines maladies, endommagent les bâtiments publics avec leurs déjections. Il est d'ailleurs interdit de les nourrir dans de nombreuses villes. Des épines sont posées sur ces bâtiments pour les empêcher de se poser, des dispositifs analogues existent aussi sur certains panneaux solaires photovoltaïques de façon que leurs déjections ne fassent pas baisser les rendements.
Certaines nuées saisonnières, comme celle du jaseur boréal, sont considérées comme une malédiction dans de nombreux pays de l'est européen[192]. Ils se déplacent en nuées vers l'ouest durant les hivers particulièrement rigoureux, semblant amener le froid avec eux.
Action de l'homme dans les disparitions actuelles
L'évolution se produit à une échelle trop lente pour être clairement perçue par les humains, mais on note une diminution certaine du nombre d'espèces et surtout du nombre d'individus au sein de nombreuses espèces. Ainsi 12 % des espèces étaient au début du XXIe siècle reconnues en danger par l'UICN, réalité confirmée par une étude de Birdlife International en 2018 selon laquelle au moins 40 % des espèces connaissent une baisse de leur population[193] - [4]. Les causes de disparition sont
- la disparition et/ou la fragmentation de leurs habitats[194].

- la chasse (pour certaines espèces) : l'homme, par sa prédation directe, est la cause de la disparition de plusieurs espèces d'oiseaux comme le Moa ou le Dronte,
- l'empoisonnement ou un affaiblissement immunitaire par les pesticides et d'autres polluants. Beaucoup d'oiseaux qui se trouvent au sommet de la pyramide alimentaire sont à ce titre victimes de la pollution en concentrant les polluants.
- Certaines espèces connaissent une augmentation plus ou moins insoupçonnée de l'hybridation ; certaines espèces proches, voyant leur habitat se morceler, se croisent en produisant même des hybrides stériles en quantité si importante que cela peut mettre ces espèces en péril. Par exemple, les nombreux oiseaux-mouches hybrides trouvés dans le nord ouest de l'Amérique du Sud peuvent représenter une menace pour la conservation de ces espèces en espèces distinctes.
- Certaines espèces rares font l'objet d'un trafic (les perroquets par exemple).
- La perte de capital génétique est un des problèmes pour le long terme, qui concerne aussi et plus encore les oiseaux domestiqués et d'élevages[195] (poules, canards, oies en particulier, à la suite de la production industrielle de poussins d'un jour à partir de reproducteurs sélectionnés). Le CNRS estimait en 2005 que 50 % des races domestiques étaient en voie de disparition.
- La diffusion de zoonoses, à cause des modalités d'élevages industriels et de transport, est également une menace, les élevages constituant des réserves potentielles de maladies ou des foyers sensibles d'incubations infectieuses.
Protection des oiseaux
La liste des espèces d’oiseaux menacés d’extinction tend à s’allonger, bien que les oiseaux soient mobiles et semblent pouvoir plus facilement échapper que d'autres espèces aux menaces qui pèsent sur eux. La liste rouge des espèces menacées, éditée par l’UICN et réactualisée en 2020, compte 1 486 espèces d’oiseaux menacées d’extinction sur 10 930, soit 13,6 % du total[196].
Les gouvernements, et de nombreuses associations de protection de la nature ou de sociétés ornithologiques, travaillent à la protection des oiseaux de diverses manières, comme la loi, la protection et la restauration des sites, et l’élevage de populations captives dans la perspective de leur réintroduction. Certaines d'entre elles visent aussi à la protection des races domestiques dont la diversité diminue.

Parmi les ONG les plus représentatives travaillant dans le domaine de la conservation des oiseaux se trouvent BirdLife International et ses délégations nationales, régionales et départementales, l'UICN, etc.
Des conventions et accords internationaux réglementent certains aspects ayant trait à la protection des espèces : Convention de Washington (CITES), Convention de Berne, Convention de Bonn, Accord sur la conservation des oiseaux d'eau migrateurs d'Afrique-Eurasie (AEWA), Migratory Bird Treaty Act, etc.
Les efforts menés par les différents partenaires ont permis, en 2004, que seize espèces d’oiseaux qui auraient disparu sans mesures conservatoires, soient toujours existantes[197].
Notes et références
Notes
- Il est impossible de donner un nombre exact car les statuts taxonomiques sont en constante évolution.
- En dehors des oiseaux, les chauves-souris sont les seuls vertébrés capables de vol battu.
- Il existe cependant de rares cas d'hétérothermie ; ainsi, les colibris entrant à l'état de torpeur sont considérés comme hétérothermes temporaires, voir Adrien Cristinelli, Torpeur et hibernation chez les vertébrés homéothermes, , PDF (lire en ligne).
- Les oiseaux ont des battements cardiaques très élevés : 600 pulsations par minute chez le colibri, 1 000 chez la mésange en activité, 70 chez l'autruche. Cf Georges B. Johnson, Peter H. Raven, Kenneth A. Mason, Jonathan B. Losos, Susan R. Singer, Biologie, De Boeck Supérieur, , p. 715.
- Le perroquet Grand Vaza fait exception avec son hémipénis externe et largement visible en période de reproduction. Sa copulation, une des plus longues du monde aviaire, dure jusqu'à une heure et demie.
- Espèces originellement endémiques ou devenues endémiques, le nombre donné est nécessairement approximatif car il dépend de la taxonomie et de l'époque retenue.
Références
- (en) Michael S. Y. Lee, Andrea Cau, Darren Naish, Gareth J. Dyke, « Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds », Science, vol. 345, no 6196, , p. 562-566 (DOI 10.1126/science.1252243).
- Jérome Fush et Marc Pons (Muséum National d'Histoire Naturelle - MNHN), « Pourquoi et comment les oiseaux font-ils leurs nids ? », sur caminteresse.fr, Ça m'intéresse,
- (en) « iucnredlist search : aves », sur iucnredlist.org (consulté le ).
- Émeline Férard, « Combien y a-t-il d'oiseaux sur Terre ? Une étude répond », sur geo.fr, (consulté le ).
- « Dans les jardins français, près de la moitié des espèces d’oiseaux observées au printemps sont en déclin », Le Monde.fr, (lire en ligne, consulté le )
- Informations lexicographiques et étymologiques de « oiseau » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales.
- Alain Rey, Dictionnaire historique de la langue française, Dictionnaires Le Robert, , p. 2449.
- Alain Rey, Dictionnaire historique de la langue française, Dictionnaires Le Robert, , p. 2491.
- Glossaire, « juvénile », sur le site Oiseaux.net, 2020 (consulté le 15 avril 2020).
- Glossaire, « immature », sur le site Oiseaux.net, 2020 (consulté le 15 avril 2020).
- Buffon, « HISTOIRE NATURELLE, GÉNÉRALE ET PARTICULIÈRE, AVEC LA DESCRIPTION DU CABINET DU ROI. Tome Vingt-deuxième », sur Oiseaux.net, (consulté le ), p. 317-325.
- Georges B. Johnson, Peter H. Raven, Kenneth A. Mason, Jonathan B. Losos, Susan R. Singer, Biologie, De Boeck Supérieur, , p. 497.
- Jean Chaline, Didier Marchand, Les Merveilles de l'Évolution, Éditions Universitaires de Dijon, , p. 200.
- (en) B.E. Lyon, R.D. Montgomerie, « Conspicuous plumage of birds: sexual selection or unprofitable prey ? », Animal Behaviour, vol. 33, no 3, , p. 1038-1040 (DOI 10.1016/S0003-3472(85)80046-4).
- (en) Donald L. J. Quicke, Mimicry, Crypsis, Masquerade and other Adaptive Resemblances, John Wiley & Sons, (lire en ligne), p. 309.
- (en) W. L N. Tickell, « White Plumage », Waterbirds : The International Journal of Waterbird Biology, vol. 26, no 1, , p. 1-12 (résumé).
- (en) John Endler, « On the measurement and classification of colour in studies of animal colour patterns », Biological Journal of the Linnean Society, vol. 41, no 4, , p. 315–352 (DOI 10.1111/j.1095-8312.1990.tb00839.x).
- (en) Fred Cooke, The Encyclopedia of Animals, University of California Press, , p. 246.
- Collectif, La nature, Encyclopédie en 14 volumes (1982) Hachette, (ISBN 2-245-01629-7).
- Damien Thiebault, « Le plumage et cycles », sur oiseaux.net, .
- Michel Delsol, L'évolution biologique, J. Vrin, , p. 287.
- Pierre-Paul Grassé, Traité de zoologie. Anatomie, systématique, biologie, Masson, , p. 99.
- (en) Josep del Hoyo, Andrew Elliott, David A. Christie et Stephen Moss, Handbook of the birds of the world : Bush-Shrikes to Old World Sparrows, vol. 14, Lynx Edicions, , 893 p. (présentation en ligne), p. 46.
- Depuis les années 1970, la longueur du bec des mésanges charbonnières en Grande-Bretagne augmente : cette longueur pourrait avoir évolué en réponse à la nourriture supplémentaire donnée par les Britanniques qui dépensent deux fois plus d'argent pour acheter des graines d'oiseaux et des mangeoires que dans le reste de l'Europe, les individus au bec plus long y ayant accès plus facilement. D'après (en) Mirte Bosse, Lewis G. Spurgin, Veronika N. Laine, Ella F. Cole, Josh A. Firth, Phillip Gienapp, Andrew G. Gosler, Keith McMahon, Jocelyn Poissant, Irene Verhagen, Martien A. M. Groenen, Kees van Oers, Ben C. Sheldon, Marcel E. Visser, Jon Slate, « Recent natural selection causes adaptive evolution of an avian polygenic trait », Science, vol. 358, no 6361, , p. 365-368 (DOI 10.1126/science.aal3298).
- Peter Hayman, Rob Hume et Marc Duquet, Reconnaître les oiseaux sans peine, Nathan Nature, coll. « Grands Guides », , 384 p. (ISBN 978-2-09-261023-7).
- (en) Ritchison, Gary, « Avian Respiration », Department of Biological Sciences Eastern Kentucky University.
- Antoine Morin, « Les Oiseaux », Université d'Ottawa.
- (en) La Vie des Oiseaux (en), Princeton University Press, (ISBN 0-691-01633-X).
- (en) Robert B. Scott, « Comparative hematology: The phylogeny of the erythrocyte », Annals of Hematology, vol. 12, no 6, , p. 340-351 (résumé).
- (en) Paul R. Ehrlich, David S. Dobkin et Darryl Wheye, « The Avian Sense of Smell », sur Birds of Stanford, Stanford University, .
- Simone Gilgenkrantz, « Le sexe des oiseaux », Médecine/Sciences, vol. 20, no 11, , p. 1004-1008 (lire en ligne, consulté le ).
- (en) P.F. Battley, T. Piersma, M.W. Dietz, S. Tang, A. Dekinga et K. Hulsman, « Empirical evidence for differential organ reductions during trans-oceanic bird flight », Proceedings of the Royal Society of London Series B-Biological Sciences, vol. 267, no 1439, , p. 191-195 (lire en ligne [PDF]).
- Pierre-Paul Grassé, Oiseaux, Masson, , p. 866.
- (en) M. Baker et A. Baker, « Niche Relationships Among Six Species of Shorebirds on Their Wintering and Breeding Ranges », Ecological Monographs, vol. 43, no 2, , p. 193–212 (résumé).
- Grande Encyclopédie Alpha des Sciences et Techniques, III: Zoologie, t. II, Paris, Grange Batelière, , 13-15 p.
- Lauralee Sherwood, Hillar Klandorf et Paul Yancey, Physiologie animale, De Boeck Supérieur, , 904 p., p. 681.
- (en) Michael Wink, Angelika Hofer, Martin Bilfinger, Elke Englert, Martinus Martin et Dietrich Schneider, « Geese and dietary alleochemicals: Food palatability and geophagy », Chemoecology, vol. 4, no 2, , p. 93-107 (lire en ligne).
- « Pourquoi le caca des oiseaux est-il blanc ? », sur Animalaxy, (consulté le )
- Gary E. Duke, A. Allan Degen et James K. Reynhout, « Movement of Urine in the Lower Colon and Cloaca of Ostriches », The Condor, vol. 97, no 1, , p. 165–173 (ISSN 0010-5422 et 1938-5129, DOI 10.2307/1368994, lire en ligne, consulté le )
- « Le système urinaire des oiseaux », Ornithomedia.com, (lire en ligne)
- Luc et Muriel Chazel, Les oiseaux ont-ils du flair ?, Quae éditions, (lire en ligne), p. 79.
- Simone Gilgenkrantz, « Le sexe des oiseaux », Médecine/science, vol. 20, no 11, (lire en ligne [PDF]).
- Bertrand Bed’Hom, Etude des caryotypes atypiques des Accipitridae (aves, falconiformes) par cytogénétique classique et moléculaire, et modélisation de leur évolution (Thèse de Biologie), , p. 1-4.
- « Agapornis roseicollis », sur Club Agapornis Français.
- (en) K. N. Mouritsen, « Day and Night Feeding in Dunlins Calidris alpina: Choice of Habitat, Foraging Technique and Prey », Journal of Avian Biology, vol. 25, no 1, , p. 55-62 (résumé).
- Mehdi Tafti, « Le Rêve », Sciences et Avenir, Hors-Série, (lire en ligne).
- (en) Niels Rattenborg, « Evidence that birds sleep in mid-flight », sur Nature, , p. 413-425.
- (en) Niels Rattenborg, Steven Lima et Charles Amlaner, « Half-awake to the risk of predation », Nature, vol. 397, no 6718, , p. 397–398 (résumé).
- (en) Buttemer W, « Energy relations of winter roost-site utilization by American goldfinches (Carduelis tristis) », Oecologia, vol. 68, no 1, , p. 126-132 (résumé).
- (en) Karl-L. Schuchmann et al., « Torpor in Hummingbirds », Bonner Zoologische Beiträge, vol. 34, , p. 273-277 (lire en ligne).
- (en) A. McKechnie, R. Ashdown, M. Christian et R. Brigham, « Torpor in an African caprimulgid, the freckled nightjar Caprimulgus tristigma », Journal of Avian Biology, vol. 38, no 3, , p. 261–266 (résumé).
- Yves Thonnerieux, « À propos du sommeil chez les canards », sur oiseaux.net, .
- (en) F.G. Buckley et P.A. Buckley, « Upside-down Resting by Young Green-Rumped Parrotlets (Forpus passerinus) », The Condor, vol. 70, no 1, , p. 89 (lire en ligne [PDF]).
- (en) J.S. Turner, « On the thermal capacity of a bird's egg warmed by a brood patch », Physiological Zoology, vol. 70, no 4, , p. 470-480 (résumé).
- (en) B. Walther, D. Clayton, « Elaborate ornaments are costly to maintain: evidence for high maintenance handicaps », Behavioral Ecology, vol. 16, no 1, , p. 89-95 (lire en ligne, consulté le ).
- (en) M. Shawkey, S. Pillai et G. Hill, « Chemical warfare? Effects of uropygial oil on feather-degrading bacteria », Journal of Avian Biology, vol. 34, no 4, , p. 345-349 (lire en ligne).
- (en) Ehrlich, P.R.; Dobkin, D.S.; Wheye, D., « The Adaptive Significance of Anting », The Auk, vol. 103, no 4, , p. 835 (lire en ligne [PDF]).
- (en) « Rose-breasted Grosbeak », sur All About Birds.
- (en) K. Brouwer et al., « Longevity records for Psittaciformes in captivity », sur ZSF Let's work for wildlife, .
- (en) Anna M. Young et al., « Survival on the ark: life history trends in captive parrots », sur NCBI, .
- « Les dix espèces qui vivent le plus longtemps au monde », sur Groupe Swiss Life (consulté le )
- (en) « Diomedea epomophora, Royal albatross : Lifespan/Longevity », sur Animal Diversity Web (ADW).
- (en) Lukas Jenni et Raffael Winkler, Moult and Ageing of European passerines : Second Edition, Christopher Helm, (ISBN 9781472941510, résumé).
- (en) Robert G. McKinney, « Skull Pneumatization in Passerines: A Table of Last Dates Many Passerines in the Northeast Can be Aged Safely by Skulling », North American Bird Bander, vol. 29, no 4, , p. 164-170 (lire en ligne).
- Commission Ornithologique de Watermael-Boitsfort, liste élaborée grâce aux données de baguage, (lire en ligne), consultée le 21 avril 2022.
- (en) Gary Ritchison, « Avian Reproduction: Nests : Mites and birds », Ornithology, Department of Biological Sciences, Eastern Kentucky University.
- (en) E. A. Bowers, « A description of Meiogymnophallus jamesoni sp. nov. (Trematoda: Gymnophallidae) from the intestine of the common scoter, Melanitta nigra L. », Annals and Magazine of Natural History, , p. 277-283 (lire en ligne).
- (en) X. de Montaudouin et al., « Infection characteristics of Himasthla elongata cercariae in cockles as a function of water current », Eases of Aquatic Organisms, , p. 1-8 (lire en ligne).
- L. Falcoz, Diptères pupipares, Paris, Fédération Française des Sociétés de Sciences Naturelles, coll. « Faune de France » (no 14), , 64 p. (présentation en ligne).
- (en) N. P. Krivosheina, N. S. Morozov et V. V. Khudyakov, « On the biology of the bird parasite Neottiophilum praeustum (Meigen, 1826) (Diptera, Neottiophilidae) », Entomological Review (pl), Pleiades Publishing Ltd, vol. 97, no 8, , p. 1048-1052 (ISSN 0013-8738, résumé).
- (en) D. M. Tompkins, T. Jones et D. H. Clayton, « Effect of vertically transmitted ectoparasites on the reproductive success of Swifts (Apus apus) », Blackwell Science, Oxford, vol. 10, no 6, , p. 733–740 (ISSN 0269-8463, résumé).
- Eugène Séguy, « Les mouches parasites des oiseaux : biologie et moyens de destruction », La Terre et la vie, Société nationale de protection de la nature et d'acclimatation de France, Paris (FRA), vol. 09, , p. 520-532 (lire en ligne)
- (en) Roy G. Bengis et al., « The Role of Wildlife in Emerging and Re-Emerging Zoonoses », Revue Scientifique et Téchnique, vol. 23, numéro 2, , p. 497-511.
- (en) S. Tsiodras, I. Kelesidis, T. Kelesidis et U. Bauchinger, « Human infections associated with wild birds », The Journal of infection, vol. 56, no 2, , p. 83-98 (résumé).
- (en) Kurt D. Reed, Jennifer K. Meece, James S. Henkel et Sanjay K. Shukla, « Birds, Migration and Emerging Zoonoses: West Nile Virus, Lyme Disease, Influenza A and Enteropathogens », Clinical Medicine & Research, vol. 1, no 1, , p. 5–12 (lire en ligne).
- « Laryngotrachéite Infectieuse Aviaire », sur WikiVet (consulté le ).
- Jean-Luc Guérin et Cyril Boissieu, « La candidose aviaire », sur AVIcampus, Toulouse, .
- (en) Gary Ritchison, « Nervous System: Brain and Special Senses II », Ornithology, Department of Biological Sciences, Eastern Kentucky University.
- Pierre-Paul Grassé, Traité de zoologie: anatomie, systématique, biologie, Masson, , p. 239.
- Nathalie Noire, Ophtalmologie des oiseaux de compagnie et des oiseaux sauvages (thèse), École Vétérinaire, Nantes, , 141 p., PDF (lire en ligne), p. 52.
- (en) G. Nevitt et K. Haberman, « Behavioral attraction of Leach's storm-petrels (Oceanodroma leucorhoa) to dimethyl sulfide », Journal of Experimental Biology, vol. 206, no 9, , p. 1497-1501 (lire en ligne).
- (en) G. Nevitt, « Sensory ecology on the high seas: the odor world of the procellariiform seabirds », Journal of Experimental Biology, vol. 211, no 11, , p. 1706-1713 (lire en ligne).
- Luc et Muriel Chazel, Les oiseaux ont-ils du flair ?, Quae éditions, , p. 88.
- Stéphane Tanzarella, Perception et communication chez les animaux, Louvain, De Boeck Université, , 334 p. (ISBN 2-8041-4896-3, lire en ligne).
- (en) Paul Rincon, « Crows and jays top bird IQ scale », sur BBC News, .
- (en) T. Bugnyar et K. Kotrschal, « Observational learning and the raiding of food caches in ravens, Corvus corax: is it 'tactical' deception? », Animal Behaviour, Vienne, Institute of Zoology, vol. 64, , p. 185–195 (lire en ligne [PDF]).
- (en) Nathan J. Emery, « Cognitive ornithology: the evolution of avian intelligence », Philosophical Transactions of The Royal Society B, Biological Sciences, vol. 361, , p. 23–43 (lire en ligne).
- Jean-Pierre Ternaux et Francois Clarac, Le Bestiaire cérébral, CNRS, , 370 p. (ISBN 9782271074065, présentation en ligne).
- Nicole Bouglouan, « Perroquets et argile… une vieille histoire ! », sur Oiseaux-Birds.
- (en) Watve Milind et al., « Bee-eaters (Merops orientalis) respond to what a predator can see », Animal Cognition, vol. 5, no 4, , p. 253-259 (résumé).
- Tim Birkhead, L'Oiseau et ses sens, Buchet/Chastel, , 370 p. (présentation en ligne).
- (en) David Lack, Ecological Adaptations for Breeding in Birds, Methuen, , 409 p. (présentation en ligne).
- (en) C. Roots, Flightless Birds, Greenwood Press, , 211 p. (ISBN 978-0-313-33545-7, présentation en ligne).
- (en) B. McNab, « Energy Conservation and the Evolution of Flightlessness in Birds », The American Naturalist, vol. 144, no 4, , p. 628-642 (résumé).
- (en) C. Kovacs et R. Meyers, « Anatomy and histochemistry of flight muscles in a wing-propelled diving bird, the Atlantic Puffin, Fratercula arctica », Journal of Morphology, vol. 244, no 2, , p. 109-125 (lire en ligne).
- (en) Kathryn Herborn et al., « Oxidative profile varies with personality in European greenfinches », Journal of Experimental Biology, vol. 214, no 10, , p. 1732-1739 (lire en ligne).
- (en) David F. Westneat et Ian R.K. Stewart, « Extra-pair paternity in birds: Causes, correlates, and conflict », Annual Review of Ecology, Evolution, and Systematics, vol. 34, no 1, , p. 365-396 (lire en ligne).
- (en) G. Wei, Z. Yin et F. Lei, « Copulations and mate guarding of the Chinese Egret », Waterbirds, vol. 28, no 4, , p. 527-530 (résumé).
- (en) Leonard A. Freed, « The Long-Term Pair Bond of Tropical House Wrens: Advantage or Constraint? », The American Naturalist, vol. 130, no 4, , p. 507-525 (résumé).
- (en) P.A. Gowaty et N. Buschhaus, « Ultimate causation of aggressive and forced copulation in birds: Female resistance, the CODE hypothesis, and social monogamy », Integrative and Comparative Biology, vol. 38, no 1, , p. 207-225 (résumé).
- Étienne Danchin, Luc-Alain Giraldeau et Frank Cézilly, Écologie comportementale, Dunod, , 664 p. (présentation en ligne).
- (en) L.W. Oring, « Avian Mating systems », Avian Biology, S. Farne, J. R. King et C. Parkes, vol. 6, , p. 1-92.
- (en) A. Cockburn, « Prevalence of different modes of parental care in birds », Proceedings of The Royal Society B, Biological Sciences, vol. 273, no 1592, , p. 1375–1383 (lire en ligne [PDF]).
- (en) Robert Burton et al., Bird Behavior, Alfred A. Knopf, , 224 p. (ISBN 0-394-53957-5).
- (en) D. Schamel, D.M. Tracy, D.B. Lank et D.F. Westneat, « Mate guarding, copulation strategies and paternity in the sex-role reversed, socially polyandrous red-necked phalarope Phalaropus lobatus », Behavioral Ecology and Sociobiology, vol. 57, no 2, , p. 110-118 (résumé).
- (en) John Warham, The Petrels - Their Ecology and Breeding Systems, Londres, Academic Press, , 440 p. (présentation en ligne).
- Gaston, A. J. (1994). Ancient Murrelet (Synthliboramphus antiquus). In The Birds of North America, No. 132 (A. Poole and F. Gill, Eds.). Philadelphia: The Academy of Natural Sciences; Washington, D.C.: The American Ornithologists' Union.
- (en) V.G. Metz & E.A. Schreiber, « Great Frigatebird (Fregata minor) », The Birds of North America, Philadelphia :Academy of Natural Sciences, A. Poole, A. and F. Gill, no 681, .
- (en) J. Ekman, « Family living amongst birds », Journal of Avian Biology, vol. 37, no 4, , p. 289-298 (résumé).
- (en) K. E. Arnold, « Group Mobbing Behaviour and Nest Defence in a Cooperatively Breeding Australian Bird », Ethology, vol. 106, no 5, , p. 385-393 (résumé).
- Pierre-Paul Grassé, Oiseaux, Masson, , p. 439.
- Luc et Muriel Chazel, Les oiseaux ont-ils du flair ?, Quae éditions, , p. 31.
- Blaise Mulhauser, La faune disparaît : Nature & Environnement, Presses Polytechniques et Universitaires Romandes, coll. « Le Savoir Suisse », , p. 65.
- (en) C.J. Barnard et D.B.A.Thompson, Gulls and Plovers : The Ecology and Behaviour of Mixed-Species Feeding Groups, Columbia University Press, , 302 p. (présentation en ligne), chap. 8 (« Kleptoparasitism: Host and Prey Selection by Gulls »), p. 217-254.
- (en) H.J. Brockmann & C.J. Barnard, « Kleptoparasitism in birds », Animal Behaviour, vol. 27, no 2, , p. 487-514 (résumé).
- F. Sueur, « Quelques cas de kleptoparasitisme chez les oiseaux », L'Oiseau et la Revue Française d'Ornithologie, vol. 60, , p. 298-299.
- (en) Alaine Camfield et Kari Kirschbaum, « Trochilidae », sur Animal Diversity Web.
- (en) N. B. Davies, Cuckoos, Cowbirds and other Cheats, T. & A. D. Poyser, , 11-25 p. (ISBN 0-85661-135-2, lire en ligne), chap. 2 (« One hundred brood parasites and some puzzles »).
- A. Bernard, « Le parasitisme chez la Nette rousse (Netta rufina) », L'Effraie, revue de la LPO Rhône, vol. 3, , p. 33-34.
- A. Bernard et P. Crouzier, « Rassemblements de couvées et parasitisme de nichées chez la Nette rousse (Netta rufina) en Dombes (Ain, France) », Nos Oiseaux, vol. 50, , p. 279-280.
- N. Flamant, « Cas de parasitisme d'une nichée de fuligules morillons (Aythya fuligula), par la Nette rousse (Netta rufina) en Bassée (77) », Bulletin de l'association des naturalistes de la vallée du Loing (ANVL), vol. 82, .
- (en) M. Sorenson, « Effects of intra- and interspecific brood parasitism on a precocial host, the canvasback, Aythya valisineria », Behavioral Ecology, vol. 8, no 2, , p. 153-161 (résumé).
- (en) J. Warham, « The Incidence, Function and ecological significance of petrel stomach oils. », Proceedings of the New Zealand Ecological Society, vol. 24, , p. 84-93 (lire en ligne).
- (en) J. P. Dumbacher, T. Spande et J. W. Daly, « Batrachotoxin alkaloids from passerine birds: A second toxic bird genus (Ifrita kowaldi) from New Guinea », PNAS, vol. 97, no 24, , p. 12970-12975 (résumé, lire en ligne [PDF], consulté le ).
- (en) J.P. Dumbacher et al., « Homobatrachotoxin in the genus Pitohui: chemical defense in birds? », Science, vol. 258, no 5083, , p. 799-801 (résumé, lire en ligne).
- Dietrich Mebs (trad. Max Goyffon), Animaux venimeux et vénéneux [« Venomous And Poisonous Animals »], Tec&Doc, , 345 p. [détail des éditions] (ISBN 2743008172 et 9782743008178).
- F. Sueur, « Comportement commensal du Grèbe castagneux Podiceps ruficollis. », Aves, vol. 16, nos 3-4, , p. 161 (lire en ligne [PDF]).
- O. Langrand, Guide des oiseaux de Madagascar, Delachaux & Niestlé, coll. « Règne animal », , 415 p. (ISBN 978-2-603-00999-4, présentation en ligne).
- F. Sueur, « Commensalisme de trois espèces d’Ardéidés vis-à-vis du Grand Cormoran. », Alauda, vol. 66, no 2, , p. 168.
- (en) Robert L. Pitman et Lisa T. Ballance, « Parkinson's Petrel Distribution and Foraging Ecology in the Eastern Pacific: Aspects of an Exclusive Feeding Relationship with Dolphins », The Condor, vol. 94, no 4, , p. 825-835 (lire en ligne [PDF]).
- (en) E.J. Temeles, Y.B. Linhart, M. Masonjones et H.D. Masonjones, « The Role of Flower Width in Hummingbird Bill Length–Flower Length Relationships », Biotropica, vol. 34, no 1, , p. 68-80 (résumé).
- (en) Jason T. Weir et Dolph Schluter, « The Latitudinal Gradient in Recent Speciation and Extinction Rates of Birds and Mammals », Science, vol. 315, no 5818, , p. 1574-1576 (résumé).
- (en) M. Brooke (présentation du livre), « Albatrosses and Petrels across the World », Marine Ornithology, Oxford University Press, vol. 33, no 1, , p. 73 (ISBN 0-19-850125-0, lire en ligne [PDF]).
- Anne-Sophie Tassart, « Des manchots empereurs ont battu le record d'apnée de l'espèce : Oiseaux », Sciences et Avenir, (lire en ligne).
- (en) Elizabeth A. Schreiber et Joanne Burger, Biology of Marine Birds, Boca Raton, Floride, CRC Press, , 740 p. (ISBN 0-8493-9882-7).
- (en) Ernst Mayr, « History of the North American bird fauna », Wilson Bulletin, Wilson Ornithological Club, vol. 58, no 1, , p. 3-41 (lire en ligne [PDF]).
- (en) Gary Ritchison, « Bird Biogeography », Ornithology, Department of Biological Sciences, Eastern Kentucky University, Richmond.
- (en) K. Chan, « Partial migration in Australian landbirds: a review », Emu, vol. 101, no 4, , p. 281-292 (résumé).
- (en) Paul R. Ehrlich, David S. Dobkin et Darryl Wheye, « Range Expansion », sur Birds of Stanford, Stanford University.
- (en) D. A. Hill et P. Robertson (présentation du livre), The pheasant: ecology, management, and conservation, Oxford, BSP Prof. Books, (lire en ligne).
- (en) M. F. Spreyer et al., « Monk Parakeet (Myiopsitta monachus) », The Birds of North America Online, Philadelphia, PA., A. Poole and F. Gill, eds, vol. 322, , p. 1-22 (lire en ligne).
- (en) Wayne J. Arendt, « Range Expansion of the Cattle Egret,(Bubulcus ibis) in the Greater Caribbean Basin », Colonial Waterbirds, vol. 11, no 2, , p. 252-262 (résumé).
- (en) Josep del Hoyo, Andrew Elliott, Jordi Sargatal et Walter J Bock, Handbook of the birds of the world : New World Vultures to Guineafowl, vol. 2, Lynx Edicions, , 638 p. (présentation en ligne), chap. 5 (« Falconidae »).
- (en) Tony Juniper et Mike Parr, Parrots: A Guide to the Parrots of the World, New Haven, Yale University Press, , 584 p. (ISBN 0-7136-6933-0).
- BioLib, consulté le 26 septembre 2020
- Jarvis, E.D. et al. (2014) Whole-genome analyses resolve early branches in the tree of life of modern birds. Science, 346(6215):1320-1331.
- T. et al. Yuri, « Parsimony and Model-Based Analyses of Indels in Avian Nuclear Genes Reveal Congruent and Incongruent Phylogenetic Signals », Biology, vol. 2, no 1, , p. 419–444 (DOI 10.3390/biology2010419).
- Avec les cinq principaux clades représentés : Agnathes (lamproies), Chondrichthyens (requins, raies), Placodermes (fossiles), Acanthodiens (fossiles), Osteichthyens (poissons osseux).
- Les poissons, amphibiens et reptiles sont des groupes paraphylétiques.
- « Systématique : ordonner la diversité du vivant », Rapport sur la Science et la technologie No 11, Académie des sciences, Lavoisier, 2010, p. 65
- (en) Bradley C. Livezey et Richard L. Zusi, « Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion », Zoological Journal of the Linnean Society, vol. 149, no 1, , p. 1–95 (lire en ligne).
- « production d'animal vivant », sur FAOStats, FAO.
- (en) A. Simeone et X. Navarro, « Human exploitation of seabirds in coastal southern Chile during the mid-Holocene », Revista Chilena de Historia Natural, vol. 75, no 2, , p. 423–431 (lire en ligne [PDF]).
- (en) A. Keane, M.D. Brooke et P.J.K. Mcgowan, « Correlates of extinction risk and hunting pressure in gamebirds (Galliformes) », Biological Conservation, vol. 126, no 2, , p. 216-233 (résumé).
- (en) M. Manzi et O.T. Coomes, « Cormorant fishing in Southwestern China: a Traditional Fishery under Siege », Geographic Review, vol. 92, no 4, , p. 597-603 (résumé).
- « Un pigeon voyageur décoré de la Croix de Guerre Américaine », Le Miroir numéro 287, (lire en ligne).
- (en) « Ostrich Riding South Africa », sur SouthAfrica.
- « Château & Jardins des Milandes ».
- « Château de Fauquemont ».
- « Fauconnerie ».
- (en) Dan Chamberlain et al., « Annual and seasonal trends in the use of garden feeders by birds in winter », The Ibis, vol. 147, no 3, , p. 563-575 (résumé).
- « Vautour - Protection », sur Vautours.info.
- (en) Kate Evans, « Return of the lost birds », New Zealand Geographic, Auckland, (lire en ligne).
- José Corti, L’Aile bleue des contes, l’oiseau : Une anthologie commentée de 101 contes sur les oiseaux, Corti, coll. « Merveilleux », (présentation en ligne).
- (en) Clement Meighan, « Prehistoric Rock Paintings in Baja California », American Antiquity, Cambridge University Press, vol. 31, no 3, , p. 372-392 (résumé).
- (en) Paul R. Ehrlich, David S. Dobkin et Darryl Wheye, « Bird Biology and the Arts », sur Birds of Stanford, Stanford University, .
- (en) A. Chandler, « The Nightingale in Greek and Latin Poetry », The Classical Journal, vol. 30, no 2, , p. 78-84 (résumé).
- José-Maria de Heredia, « La Mort de l’Aigle », sur Un jour Un poème.
- (en) P. Thankappan Nair, « The Peacock Cult in Asia », Asian Folklore Studies, vol. 33, no 2, , p. 93-170 (résumé).
- Frédéric Manns, « Le symbolisme de la colombe au Proche-Orient ancien », sur interbible.org, .
- (en) Nancy Hatch Dupree, « An Interpretation of the Role of the Hoopoe in Afghan Folklore and Magic », Folklore, vol. 85, no 3, , p. 173-193 (lire en ligne).
- (en) Jackie Chappell, « Living with the Trickster: Crows, Ravens, and Human Culture », PLoS Biology, vol. 4, no 1, , p. 16-17 (lire en ligne [PDF]).
- « Genèse : versets 8.6 à 8.12 », sur AELF.
- « Premier Livre des Rois : chapitre 17.4 à 6 », sur AELF.
- Philon d'Alexandrie, De Somniis II, 212-214.
- « Sourate XVI, An-Nahl / Les abeilles : verset 79 », sur Coran-Français.
- (en) R.A. Zann, The Zebra Finch : A Synthesis of Field and Laboratory Studies, Oxford, Oxford University Press, , 335 p. (présentation en ligne).
- (en) Josep del Hoyo, Nigel J. Collar et al., HBW and BirdLife International Illustrated Checklist of the Birds of the World : Non-passerines, vol. 1, Lynx Edicions, , 903 p. (présentation en ligne).
- (en) James Fisher, Birds as Animals, Hutchinson's University Library, , p. 30.
- (en) James Fisher, Les Oiseaux dans le règne animal, Payot, , p. 12.
- Valérie Chansigaud, Histoire de l'ornithologie, Delachaux et Niestlé, , p. 26.
- Valérie Chansigaud, Histoire de l'ornithologie, Delachaux et Niestlé, , p. 34.
- (en) Josep del Hoyo, Andrew Elliott, Jordi Sargatal, Handbook of the birds of the world : Ostrich to Ducks, vol. 1, Lynx Edicions, , 696 p., « Class Aves ».
- Valérie Chansigaud, Histoire de l'ornithologie, Delachaux et Niestlé, , p. 85.
- (en) Josep del Hoyo, Andrew Elliott, David A Christie et Bruce Murray, Handbook of the birds of the world : Broadbills to Tapaculos, vol. 8, Lynx Edicions, , 845 p., p. 29.
- Valérie Chansigaud, Histoire de l'ornithologie, Delachaux et Niestlé, , p. 200.
- Valérie Chansigaud, Histoire de l'ornithologie, Delachaux et Niestlé, , p. 202.
- (en) B.L. Monroe et C.G. Sibley, A World Checklist of Birds, New Haven, Yale University Press, , 416 p. (ISBN 978-0-300-07083-5, présentation en ligne).
- M. Walters, L'inventaire des Oiseaux du Monde, Lausanne, Paris, Delachaux et Niestlé, coll. « La bibliothèque du naturaliste », , 381 p. (ISBN 978-2-603-01107-2).
- Jean Duval, « LES OISEAUX NUISIBLES AUX CULTURES », sur eap.mcgill.ca, Ecological agriculture project, McGill University, .
- François Sarrazin et E. Danchin, Dynamique des populations réintroduites : le cas du Vautour fauve dans les Causses (thèse de doctorat en écologie), Université de Paris 06, (présentation en ligne).
- Eduardo Camacho-Hübner, « Jaseurs, tsunamis et décalage négatif de la connaissance », EspacesTemps.net, (résumé).
- Ana Benabs, « Une espèce d'oiseau sur huit est menacée d'extinction, affirme une étude menée au niveau mondial », sur france24.com, .
- (en) K. Norris et D. Pain, Conserving Bird Biodiversity: General Principles and their Application, Cambridge University Press, , 337 p. (ISBN 978-0-521-78949-3), chap. 10 (« Fragmentation, habitat loss and landscape management »).
- Elisabeth Blesbois et al., « Première constitution d’une réserve génétique ex-situ pour l’espèce Gallus gallus », Sixièmes Journées de la Recherche Avicole, Saint-Malo, , p. 539-543 (lire en ligne [PDF]).
- (en) « Summary Statistics, Summary Tables », sur IUCN, (consulté le ) Red List version 2020-1.
- (en) S. Butchart, A. Stattersfield et N. Collar, « How many bird extinctions have we prevented? », Oryx, vol. 40, no 3, , p. 266-278 (lire en ligne [PDF]).
Voir aussi
Références taxinomiques
- (en) Référence Tree of Life Web Project : Aves
- (en) Référence Catalogue of Life : Aves (consulté le )
- (en) Référence Fauna Europaea : Aves (consulté le )
- (en) Référence Paleobiology Database : Aves Linnaeus 1758
- (fr+en) Référence ITIS : Aves
- (en) Référence World Register of Marine Species : taxon Aves (+ liste espèces)
- (en) Référence Animal Diversity Web : Aves
- (en) Référence NCBI : Aves (taxons inclus)
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023
- (en) Référence Fonds documentaire ARKive : Aves
- (en) Liste d'Alan P. Peterson
Ouvrages généraux
- Guilhem Lesaffre, Nouveau Précis d’ornithologie, Paris, Vuibert, , 216 p. (ISBN 2-7117-7160-1)L'ouvrage présente de nombreuses informations sur l'anatomie, la physiologie, l'écologie, l'éthologie et la systématique. Il convient particulièrement à ceux qui aimeraient aller plus loin dans la connaissance des oiseaux.
- (en) C. Perrins, The New Encyclopedia of Birds, Oxford, Oxford University Press, , 656 p.L'ouvrage présente les principales caractéristiques des oiseaux et un panorama complet des familles (caractéristiques principales, nombre d'espèces et de genres, principales espèces, distribution, habitat, taille, plumage, voix, nid, œufs, régime alimentaire et statut de conservation) selon la classification classique.
Oiseaux du monde
- (en) coll. « Handbook of the Birds of the World » édition Lynx Edicions, Barcelone
- del Hoyo J., Elliott A. & Sargatal J.
- : vol. 1, Ostrich to Ducks (1992), ICBP, 696 p
- : vol. 2, New World Vultures to Guineafowl (1994), BirdLife International, 638 p
- : vol. 3, Hoatzin to Auks (1996), BirdLife International, 821 p
- : vol. 4, Sandgrouse to Cuckoos (1997), B. I., 679 p
- : vol. 5, Barn-owls to Hummingbirds (1999), B. I., 759 p
- : vol. 6, Mousebirds to Hornbills (2001), B. I., 589 p
- : vol. 7, Jacamars to Woodpeckers (2002), B. I., 613 p
- del Hoyo J., Elliott A. & Christie D.
- : vol. 8, Broadbills to Tapaculos (2003), B. I., 845 p
- : vol. 9, Cotingas to Pipits and Wagtails (2004), B. I., 863 p
- : vol. 10, Cuckoo-shrikes to Thrushes (2005), B. I., 895 p
- : vol. 11, Old World Flycatchers to Old World Warblers, B. I., 798 p
- : vol. 12, Picathartes to Tits and Chickadees, B. I., 816 p
- (en) Monroe B.L. & Sibley C.G. (1997) A World Checklist of Birds, Yale University Press, New Haven.
- Walters M. (1998) L’Inventaire des Oiseaux du Monde, Delachaux et Niestlé, Lausanne, Paris, 381 p.
- Agnès Giannotti, Oiseaux d'Afrique, les plus belles histoires, 144 p, 340 photographies couleur, éditions Grandvaux, (ISBN 978-2-909550-59-6)
Oiseaux par aires géographiques
- Barré N., Barau A. & Jouanin C. (1996) Oiseaux de la Réunion. Éditions du Pacifique, Paris, 208 p.
- Burfield I. & van Bommel F. (2004) Birds in Europe : population estimates, trends and conservation status. BirdLife International, Cambridge, 374 p.
- Dubois P.J., Le Maréchal P., Olioso G. & Yésou P. (2000) Inventaire des oiseaux de France. Nathan, Paris, 397 p.
- Groupe d'étude et de protection des oiseaux en Guyane (2003) Portraits d'oiseaux guyanais. Ibis Rouge Éditions, Guyane, Guadeloupe, Martinique, Paris, Réunion, 479 p.
- Langrand O. (1995) Guide des Oiseaux de Madagascar. Delachaux & Niestlé, Lausanne, Paris, 415 p.
- Collectif (2013). Merci les oiseaux! : le magazine qui vous donne des ailes. Terre&Nature, Lausanne, 98 p. (ISBN 9782839912945)
- Todd F.S. & Genevois F. (2006) Oiseaux & Mammifères antarctiques et des îles de l'océan austral. Kameleo, Paris, 144 p.
- Tucker G.M. & Heath M.F. (1994) Birds in Europe. Their Conservation Status. BirdLife International, Cambridge, 600 p.
- Roger Toy Peterson (1989) Les oiseaux de l'est de l'Amérique du Nord. Broquet, Québec, 386 p.
- Morin Jérôme. (2009) Guide des oiseaux des Villes et des Jardins Le livre. Collection « Fous de nature » Éditions Belin, Borché, 224 Pages.
Oiseaux par groupes taxonomiques
- Clement P., Harris A. & Davis J. (1999) Finches & Sparrows. Christopher Helm, Londres, 500 p.
- Dupuyoo M. (2002) Diamants, Papes et Capucins. Estrildés de l'Indo-Pacifique. Jardin d'Oiseaux Tropicaux, La Londe les Maures, 240 p.
- Forshaw J.M. (2006) Parrots of the World. An identification guide. Princeton University Press, Princeton, Oxford, 172 p.
- Géroudet P. (1999) Les Palmipèdes d'Europe. Delachaux et Niestlé, Lausanne, Paris, 510 p.
- Gille D. & François B. (2003) La famille des Inséparables. CDE, Sainte-Reine-de-Bretagne, 151 p.
- Madge S. & Burn H. (1996) Corbeaux et Geais. Guide des Corbeaux, Geais et Pies du monde entier. Vigot, Paris, 184 p.
- Mario D. & Conzo G. (2004) Le grand livre des perroquets. de Vecchi, Paris, 287 p.
- Ravazzi G. (1995) Guide des oiseaux exotiques. Les Diamants et autres Estrildidés. De Vecchi, Paris, 157 p.
- Taylor D. (2006) Guide des limicoles d'Europe, d'Asie et d'Amérique du Nord. Delachaux & Niestlé, Paris, 224 p.
Revues scientifiques spécialisées
- Alauda, revue internationale d'ornithologie, seul périodique ornithologique français à vocation internationale survivant du début du XXe siècle
- Ardea, revue néerlandaise publiée en anglais
- Bird Study, revue britannique
- The Auk, revue américaine
- The Ibis, revue britannique
Revues de vulgarisation
- Aves, revue francophone belge
- British Birds, revue britannique consacrée à l'ornithologie de terrain
- Nos Oiseaux, revue suisse francophone
- Ornithos, revue bimestrielle de la Ligue pour la protection des oiseaux
- L'Oiseau Magazine, revue trimestrielle de la Ligue pour la protection des oiseaux
Voir aussi Orientation bibliographique en ornithologie et Liste de périodiques ornithologiques.
Articles connexes
- Anatomie des oiseaux
- Atlas ornithologique
- Avialae
- Guide phylogénétique illustré du monde animal
- Identification des oiseaux
- Indice d’abondance des populations d’oiseaux communs
- Liste d'espèces d'oiseaux décrites entre 2001 et 2005
- Liste d'espèces d'oiseaux décrites entre 2006 et 2010
- Lutte contre les oiseaux
- Volatile

Liens externes
- Ressources relatives au vivant :
- Global Biodiversity Information Facility
- (en) Animal Diversity Web
- (en) Australian Faunal Directory
- (sv) Dyntaxa
- (en) EPPO Global Database
- (en) EU-nomen
- (en) Fauna Europaea
- (en) Paleobiology Database
- (mul + en) iNaturalist
- (en) Interim Register of Marine and Nonmarine Genera
- (nl) Nederlands Soortenregister
- (en + en) New Zealand Organisms Register
- (en) Plazi
- (en) Système d'information taxonomique intégré
- (en) World Register of Marine Species
- (en) ZooBank
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Britannica
- CALS Encyclopedia of Arkansas
- Enciclopedia De Agostini
- Encyclopædia Iranica
- The Encyclopedia of Oklahoma History and Culture
- Encyclopedia of the Great Plains
- L'Encyclopédie canadienne
- Gran Enciclopedia Aragonesa
- Gran Enciclopèdia Catalana
- Handbook of Texas Online
- Larousse
- Nationalencyklopedin
- Maine: An Encyclopedia
- New Georgia Encyclopedia
- Store norske leksikon
- Universalis
- The West Virginia Encyclopedia
- Oiseaux.net, portail et guide encyclopédique de l'avifaune. Fiches descriptives des oiseaux du Monde, avec photos, dessins et chants.
- Ornithomedia.com, le web de l'ornithologie.
- Les Oiseaux des Hautes-Fagnes, livre présentant aussi les facteurs contrôlant les populations d'oiseaux
- "Les oiseaux, vers un printemps silencieux", Le Temps d'un Bivouac, 14 août 2019
- Marc Mennessier, « Oiseaux: comment installer des nichoirs au jardin ? », sur Le Figaro, (consulté le )
- Jardin, « 5 oiseaux à observer dans son jardin en hiver », sur Le Figaro, (consulté le )