Communication animale
La communication animale regroupe l'ensemble des échanges d'information entre des individus d'une même espèce (communication intraspécifique) ou d'espèces différentes (communication extraspécifique) chez les animaux.
Différence entre communication et information
Les animaux communiquent entre eux pour différentes raisons : séduction du partenaire, compétition pour les ressources, recherche de nourriture. Ils utilisent pour cela un signal ou un message (support physique de l'information), comme une odeur, un son, un mouvement, ou un signal électrique. Dans la communication, le signal émis par un animal entraîne une modification du comportement ou de l'état physiologique d'un autre animal, qui utilise l'information pour prendre des décisions qui se traduisent par une réponse comportementale. La réponse de l'animal est susceptible d'avoir des conséquences sur la survie de l'animal à l'origine du message, aussi bien que sur la sienne propre.
Toute communication implique au moins deux individus, qu'on nomme émetteur et récepteur en suivant une métaphore mécaniste. La situation est généralement plus complexe : les systèmes naturels s'organisent souvent en « réseaux de communication », où chaque individu peut être à la fois émetteur et récepteur. La présence d'auditeurs non souhaités, parasitant l'information, est également fréquente.
Cependant tout échange d'information entre animaux n'est pas assimilable à une communication. Par exemple, une souris qui fait du bruit en se déplaçant ne communique pas avec la chouette qui la chasse. C'est la différence entre indice (purement causal, sans intention, devenant signal interprétable) et signe intentionnel[1] - . De même, l'écholocation qui permet à un animal de s'orienter dans l'espace ou de repérer ses proies grâce à l'écho de ses propres sons ne relève pas d'un processus de communication (production intentionnelle permettant d'obtenir des informations sur l'environnement, pour soi-même[2]).
On se restreint ici à la communication intraspécifique, éliminant de fait les situations plus ou moins litigieuses — mais parfois réelles — où une espèce utilise les signaux d'une autre.
Différentes approches de la communication animale
La compréhension des phénomènes de communication animale comprend différents éléments, correspondant à des angles distincts :
- la compréhension de la chaîne de transmission de l'information ;
- la diversité des signaux utilisés ;
- les modalités de production de signaux par l'animal émetteur ;
- la transmission des signaux dans l'environnement ;
- la réception des signaux par l'animal récepteur ;
- les modalités de codage de l'information ;
- les modalités d'acquisition (angle ontogénétique, génétique du comportement de communication, apprentissage de la production des signaux et de leur contexte d'utilisation ;
- les rôles biologiques (fonctions) de la communication ;
- l'étude des implications évolutives des communications (évolution des processus de communication, et rôle des communications dans l'évolution des espèces).
- les applications de la connaissance de la communication animale (par exemple pour la protection des espèces animales, ou la lutte contre les nuisances).
L'étude du comportement animal est ainsi au carrefour de nombreuses disciplines : la physiologie (dont les neurosciences mais aussi l'étude de l'ensemble des grandes fonctions vitales telles que la nutrition et la reproduction), l'écologie et l'évolution bien sûr, mais aussi l'anatomie ou la génétique, supposant de changer d'échelle dans l'explication des phénomènes[3].
La chaîne de transmission de l'information
Les signaux, supports physiques de l'information
- signaux chimiques (phéromones : molécules, souvent émises en « bouquets », milieux aériens et aquatiques, peuvent être rémanents ou non, plus ou moins volatils, souvent grande intraspécificité),
- signaux visuels (ondes électromagnétiques, statiques et/ou dynamiques, dépendants de la qualité et de la quantité de lumière ambiante sauf en grand profondeur sous-marine où les photons sont générés par l'animal lui-même, propagation sensible aux obstacles physiques telle la végétation),
- signaux acoustiques et vibrations du substrat (ondes mécaniques, produites par vibrations de structures anatomiques, soit via passage de flux d'air, soit par frottement entre deux surfaces dures, utilisables dans l'eau comme dans l'air, grande portée potentielle, faible rémanence),
- signaux haptiques (ou tactiles)
- signaux électriques (dans l'eau seulement, nécessité d'organes spécialisés d'origine musculaire, utilisés essentiellement par des poissons, surtout en milieu trouble)
- signaux kinésiques (mouvements du corps) ou proxémiques (distances)
Un signal est donc un processus physique (en principe une onde ou assimilable à une onde), qui code de l'information via des paramètres de fréquence, d'amplitude et temporels (paramètres de fréquence : ex. son aigu/grave ; paramètres d'amplitude : ex. intensité sonore ; paramètres temporels : rythme d'émission et modulations de fréquence et d'amplitude). Les signaux chimiques peuvent être assimilés à une onde : spectre de fréquence : composition moléculaire du signal + concentration relative des composés, amplitude : concentration du bouquet phéromonal, paramètres temporels : variations dans le temps de la composition et de la concentration du bouquet phéromonal.
La production des signaux
- Des organes souvent spécialisés.
Un exemple d'organe producteur de signal de communication montrant :
- l'existence de structures anatomiques spécialisées dans l'émission d'un signal donné (ex. syrinx des oiseaux chanteurs, cymbale des homoptères, stridulation du grillon, glandes phéromonales des abeilles, larynx des mammifères …)
- la présence d'un contrôle nerveux et éventuellement hormonal (ex. noyaux du chant dans le cerveau des oiseaux).
Certains signaux sont plus difficiles à émettre, plus coûteux en énergie. Les propriétés des signaux sont fonction de la biophysique et de la biochimie de l'émission et de l'adéquation entre l'organe émetteur et l'environnement.
- Des organes pilotés par la génétique, l'apprentissage et le contexte.
La capacité à émettre un signal de communication peut être piloté par divers facteurs, susceptibles d'interagir de façon compliquée.
- Possible contrôle génétique de la structure des signaux produits (ex. expériences de croisement inter-spécifique avec effet sur le rythme du chant chez grillon, drosophile).
- Rôle limitant des capacités anatomophysiologiques de l'animal (ex. seuls de « gros » animaux peuvent émettre des sons graves ; l'homme ne peut pas émettre d'ultrasons…).
- Possible apprentissage de la production des signaux (le modèle expérimental le plus abouti est celui des oiseaux chanteurs : il faut décrire ici des expériences d'isolement des oisillons permettant de mettre en évidence que les jeunes doivent apprendre leur chant en copiant un tuteur. Ce modèle est également l'occasion de changer d'échelle pour l'étude des processus biologiques en jeu : c'est dans les noyaux cérébraux moteurs pilotant le chant chez l'oiseau qu'a été mise en évidence pour la première fois une neurogenèse chez un vertébré adulte).
- Possible apprentissage du contexte d'utilisation (un exemple livresque est celui des signaux d'alarme chez les singes vervets où des cris de trois structures acoustiques différentes sont émis dans trois contextes de prédation différents : attaque aérienne par un aigle, attaque terrestre léopard par exemple, et arrivée d'un serpent. L'utilisation adéquate de ces cris par les jeunes est soumise à un apprentissage).
La décision d'envoyer ou non un signal à un moment donné, son niveau de complexité et de puissance (son intensité) sont fonction de l'état énergétique et psychologique de l'émetteur, ainsi que des risques encourus (prédation, présence de congénères compétiteurs ou alliés, modulation hormonale, etc.).
La transmission des signaux dans l'environnement
Pour passer de l'émetteur au récepteur, tout signal devra traverser un « canal de transmission » selon le terme consacré (on dit aussi « canal de propagation »). Cette traversée n'a rien d'anodine pour la communication : le canal de transmission va agir comme un filtre, modifiant le signal émis. La conséquence est majeure : le signal reçu par l'animal récepteur ne peut être identique au signal envoyé par l'émetteur. Cette contrainte, potentiellement majeure, pèse sur tout processus de communication.
La portée d'un signal dépend de ses caractéristiques physiques et des contraintes du milieu. On peut citer comme exemple la différence entre les milieux ouverts (i.e., sans végétation) et les milieux forestiers pour la propagation des signaux acoustiques et visuels (ex. réverbération des sons en forêt ⇒ modification de la durée des signaux), la différence eau/air pour la propagation des sons (le milieu aquatique est idéal pour la transmission des ondes sonores), l'importance des courants d'eau et d'air pour la propagation des phéromones.
La réception des signaux
La réception du signal par l'animal émetteur est bien sûr une étape cruciale du processus de communication. Cette partie doit détailler les processus par lesquels les signaux sont intégrés par l'organisme au niveau de structures réceptrices (transduction en signaux nerveux), comment l'information est décodée et les conséquences sur l'animal récepteur sur le plan de l'état physiologique et de la réponse comportementale.
- Chimioréception, photoréception, électroréception, audition
Un exemple détaillé d'une des modalités permet de présenter l'ensemble de la chaîne de réception depuis la stimulation de l'organe récepteur, le codage nerveux, jusqu'à l'intégration par le système nerveux central (exemples pouvant être utilisés : organe auditif chez le grillon champêtre ; organe voméronasal des mammifères ; réception des phéromones par l'antenne du Bombyx ; œil des Chordés, etc.).
En vue d'une généralisation, les principes suivants doivent être explicités : un récepteur sensoriel est une fenêtre ouverte sur le monde extérieur (c'est un filtre, sensible à une certaine bande passante du signal ; ex. l'oreille des chauve-souris est sensible aux ultrasons, pas l'oreille humaine), chargée de la transduction du signal de communication en un signal nerveux.
Interprétation des signaux et décision
- Perception et classification du signal par le système nerveux central : reconnaissance, éventuelle discrimination entre les signaux et classification des signaux perçus.
- Interprétation du signal et extraction de l'information du signal. Traitement de l'information (ex. rôle des aires associatives corticales chez Vertébrés).
- Élaboration de la réponse au signal (décision) sur la base de l'interprétation du signal par le système nerveux central. Des alternatives à une réponse comportementale directe existent, telle l'attente de la réception de signaux complémentaires. Mise en jeu possible de la mémoire à court et long terme.
- En conclusion : l'individu récepteur du signal change son comportement. Exemple : si le signal a fonction de prévenir le danger, le récepteur va changer de comportement et adopter un comportement méfiant et agressif.
Le codage de l'information
L'information est codée, et le code dépend de chaque espèce, voire parfois de chaque communauté. Par exemple les lucioles modulent leur fréquence d'allumage et la « forme » du signal. Les abeilles dansent devant leurs congénères de retour à la ruche pour signifier la distance à la nourriture, la direction par rapport au soleil, la nature de la nourriture.
Le code peut être discret (des signaux de structure différente, chacun codant une ou plusieurs informations différentes) ou gradé ou gradué (des signaux dont les caractéristiques sont modifiées graduellement pour coder une information différente ; ex. aboiement du chien).
Les codes utilisés dans les communications animales sont utilisés pour signifier une identité (espèce, groupe, sexe, individu), la description de l'environnement (ex. présence et localisation de la nourriture, de prédateurs), l'état physiologique et cognitif de l'animal (parades nuptiales, signaux montrant l'état de réceptivité sexuelle, âge…).
Les fonctions de la communication
Le choix du partenaire et la sélection sexuelle


- Signaux informant sur l'identité spécifique et les caractéristiques individuelles de l'émetteur (état physiologique, réceptivité sexuelle, niveau d'agressivité, statut social dominant, dominé, son histoire de vie, patrimoine génétique).
- Souvent utilisation de plusieurs canaux de communication concomitamment (signaux visuels, acoustiques et chimiques ; ex. lors des parades nuptiales des cervidés).
- En règle générale, la présence d'un fort risque de prédation entraîne la diminution de la préférence pour un signal sexuel visible.
La communication entre génération
Elle est développée chez les animaux pratiquant les soins aux jeunes (nombreuses espèces de poissons, crocodiles, oiseaux, mammifères).
- Les signaux permettent
- la reconnaissance parents-jeunes (processus particulièrement développé lorsque le risque de confusion est élevé, tel dans les colonies d'oiseaux marins et de pinnipèdes);
- la quémande alimentaire par les jeunes est utilisée par les jeunes pour solliciter le nourrissage par leurs parents et il s'agit d'une démonstration extravagante qui peut être coûteuse pour la fitness des jeunes. La quémande alimentaire est un signal informatif en partie honnête puisque l'intensité de la quémande est en principe corrélée à l'état de satiété.
Cependant elle peut aussi être un signal malhonnête résultant de la compétition entre les jeunes d'une même nichée. Des expériences ont été menées sur la quémande alimentaire chez le Moineau domestique (Passer domesticus), afin de tester les deux hypothèses suivantes :
- Les parents sont-ils sensibles à l'intensité de la quémande alimentaire des oisillons ?
Être un parent sensible à la quémande de ses oisillons permet de diminuer deux risques opposés : le risque de perdre trop de temps en revenant trop tôt au nid (gain de temps pour les parents donc augmentation de leur fitness) et le risque de négliger certains oisillons au hasard à plusieurs reprises.
- L'intensité de la quémande alimentaire a-t-elle une origine génétique ?
L'intensité de la quémande des oisillons a une origine génétique relativement faible. En revanche, de nombreux facteurs sociaux et environnementaux induisent des variations de l'intensité de la quémande (environnement éducatif commun aux oisillons et compétition frères-sœurs).
La compétition territoriale
- Caractéristiques des signaux territoriaux : en principe grande portée (ex. chants des oiseaux) et/ou rémanence temporelle (ex : marquage par les carnivores).
- La structure du signal peut être corrélée aux capacités physiques de l'émetteur (permet l'évitement des affrontements physiques directs).
- Les signaux peuvent parfois permettre la reconnaissance entre compétiteurs voisins (et ainsi permettre d'éviter des combats = principe du « cher ennemi »).
La reconnaissance de parentèle et/ou du groupe social
Il s'agit ici d'un rôle tout à fait primordial des communications animales.
Souvent observée lors des interactions parents-jeunes, la reconnaissance de parentèle peut s'étendre aux autres membres du groupe familial. Ex. sociétés d'insectes eusociaux (certains hyménoptères, isoptères).
La mise en place de la reconnaissance de parentèle peut s'effectuer selon deux grandes modalités :
- par reconnaissance phénotypique (ex. chez la souris, les individus préfèrent des congénères à odeur semblable ; or les molécules odorantes dépendent du CMH des individus)
- par apprentissage (ex. la jeune otarie apprend à reconnaître la voix de sa mère dans les 2-5 jours qui suivent sa naissance).
La reconnaissance du groupe social est importante pour permettre des relations de réciprocité entre individus (entre-aide, altruisme…) Elle est l'apanage des groupes possédant une forte structure et hiérarchie. Les singes (babouins) en sont un bon exemple, mais également les hyènes car ces deux espèces possèdent une construction sociale assez similaire. Cette reconnaissance est, entre autres, acoustique et visuelle.
Tout d'abord, singes et hyènes savent tous deux reconnaître les individus qui leur sont apparentés :
- Reconnaissance mère/petit : chez la hyène, lorsque le cri d'un petit est passé au haut-parleur, la mère s'approche de celui-ci et cherche même son petit. Il en est de même pour les singes (vervets.)
- Reconnaissance de la « famille » : lorsque le cri d'un petit est passé au haut-parleur, les individus qui lui sont apparentés (frères et sœurs, tante, cousins…) cherchent activement le petit, d'autant plus s'ils sont fortement apparentés.
Ces deux espèces à fort réseau social sont donc capables de reconnaître le cri d'un de leur proche. Mais sont-elles capables de reconnaître les relations d'affiliation qui existent entre différents individus ? Sont-elles capables de dire que tel individu est parent de tel autre ?
Les babouins en seraient tout à fait capables ! En effet, les femelles babouins peuvent reconnaître des liens qui unissent des individus non apparentés, et même adapter leur comportement en fonction des individus impliqués dans une relation. Par exemple, lorsqu'une femelle entend une dispute entre un de ses parents et un autre individu, elle se montrera ensuite plus distante si elle doit interagir avec un apparenté de cet individu.
De même, d'autres exemples existent chez différentes espèces de singes : en effet, chez les singes vervets, lorsque le cri d'un petit est passé au haut-parleur, toutes les autres femelles regardent la mère, ce qui signifie qu'elles associent bien un petit avec sa mère. Chez les macaques, une femelle entraînée serait capable d'associer des photos de petit avec celles de leurs mères, ce qui montre encore une fois que ces singes sont capables de reconnaître visuellement des individus affiliés.
Chez les hyènes, en revanche, un tel processus serait pour l'instant impossible. En effet, lorsque le cri d'un petit donné est passé au haut-parleur, les autres individus ne se tournent pas plus vers la mère que vers un autre individu. Cependant, les hyènes évoluant dans des groupes de plus d'une vingtaine d'individus et reconnaissant très bien leur position hiérarchique au sein de celui-ci, il est difficile de croire qu'elles sont incapables de reconnaître les affiliations entre individus. D'autres investigations sont nécessaires pour le prouver.
Le degré de reconnaissance sociale semble donc lié à la complexité de l'organisation du groupe social, mais aussi à l'évolution. Certains animaux semblent donc plus aptes que d'autres, les singes étant les maîtres en la matière.
La marque de statut social

Dans un groupe social (ex. chez insectes, poissons, oiseaux, mammifères), le signalement du statut de l'individu est une donnée importante permettant la structuration du groupe (dominant/dominé, reproducteur/non reproducteur, sexe, âge, signatures individuelles et de groupe, …). Ces signaux sont souvent corrélés à l'état physiologique du moment (ex. protrusion sexuelle des singes femelles), mais aussi à l'histoire passée de l'individu (notion de « badges de statut »).
La coordination sociale
- Les stratégies de communication antiprédatrices
- Signaux d'alarme et de détresse, dont la structure est adaptée à la fonction (voir l'exemple des singes vervets cités plus haut ; il y a également des exemples chez les oiseaux – mésanges qui modifient la longueur de leur cri d'alarme en fonction de la taille du prédateur repéré). Ce sont souvent des signaux que leur structure rend difficiles à localiser par le prédateur.
- Les collaborations au sein du groupe social
- Coopération pour la recherche de nourriture (ex. danse des abeilles ; chasse en groupe des grands carnivores).
- Utilisation de signaux lors des alliances sociales (ex. au sein des groupes de singes)
La communication est le processus fondateur de toute interaction sociale. Il serait judicieux dans cette conclusion d'aborder les notions d'altruisme et de bénéfice réciproque (amenée par les cris d'alarme, les quémandes alimentaires des jeunes). Les rapports coûts/bénéfices et un élargissement à la théorie des jeux sont aussi envisageables.
Communication et évolution
Deux aspects peuvent être différenciés :
- l'évolution des communications animales (en lien les contraintes pesant sur les mécanismes de communication et leurs adaptations associées)
- le rôle des communications animales dans l'évolution des espèces (et en particulier leur intervention dans les phénomènes de spéciation).
Adaptations structure-fonction
La taille de l'animal, sa température interne, son organisation anatomomorphologique sont autant de facteurs qui contraignent les possibilités d'émission et de réception des signaux.
Ces facteurs sont souvent d'abord liés à des contraintes phylogénétiques. Par exemple, les oiseaux chanteurs ont une syrinx et un contrôle nerveux moteur associé qui leur permet d'émettre des signaux acoustiques potentiellement complexes ; les crocodiles n'ont pas d'organe émetteur d'onde sonore spécialisé. Ils peuvent également être modulés par des facteurs ontogénétiques (maturité sexuelle) ou dépendants de l'environnement (température externe chez hétérothermes par exemple). En conséquence, chaque animal dispose d'une gamme de signaux de communication potentiels. Cette gamme diffère entre les espèces, les populations, voire les individus (même au sein d'une seule famille : voir protéine Forkhead-P2).
« Coévolution » émetteur-récepteur
- Correspondance (pas toujours parfaite) entre caractéristiques du signal émis par l'émetteur et possibilités de réception par l'animal récepteur (idée de coévolution organes émetteurs – organes récepteurs ; ex. correspondance entre les fréquences des sons émis et les fréquences pouvant être perçues par l'oreille
- Idem pour électrorécepteurs des poissons communiquant par ondes électriques).
Sélection sexuelle
- Notions d'emballement dit « de Fisher » et conséquence sur les signaux de communication;
- Théorie du Handicap et question de « l'honnêteté du signal » (hypothèse = un signal porte une information fiable quant aux caractéristiques intrinsèques de l'émetteur);
- Théorie de l'exploitation sensorielle des signaux par le récepteur & biais sensoriel.
Relations parents-jeunes et interactions inter-jeunes
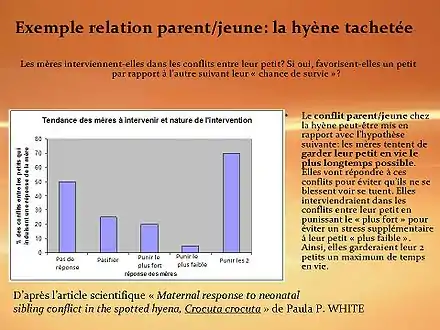

Chez les espèces présentant des soins aux jeunes, les parents et les jeunes peuvent avoir des intérêts divergents (théorie du conflit parents-progéniture).
De plus, les jeunes peuvent être en compétition pour l'accès aux ressources parentales. Se mettent donc en place des stratégies de signalisation lors de la quémande alimentaire (exagération des signaux, signalisation concurrente des jeunes ou au contraire coopération lors de l'émission…).
Corrélation système de communication / système social
La « complexité » (attention à ce terme, pris ici sans jugement de valeur !) d'un système de communication va dépendre a minima du répertoire de signaux (nombre de signaux différents), de la variété des informations transmises, de la plasticité du système (possibilités d'apprentissage). Elle sera donc corrélée à la « complexité » du système social de l'espèce : plus les individus vivent des interactions sociales variées, complexes (compétition, collaboration, réconciliation – chez les primates par exemple) et changeantes au cours du temps, plus le système de communication est élaboré.
Les récepteurs non désirés
La présence de prédateurs, de parasites potentiels ou d'individus concurrents peut représenter une contrainte importante sur les signaux (risque d'être repéré). On constate l'utilisation de signaux discrets et/ou difficilement localisables (ex. cris d'alarme aigus des passereaux, cris de copulation à faible intensité de nombreuses espèces de vertébrés).
Contraintes environnementales et adaptation des communications
Certains environnement (ex sous la terre, dans l'eau, dans un feuillage dense, en bord de mer ou près d'une cascade bruyante, etc.) atténuent et dégradent les signaux visuels, biochimiques et/ou sonores (« bruitage » du signal).
Divers types d'adaptations permettent à des animaux de néanmoins communiquer en présence d'un bruit de fond (la notion de bruit de fond s'applique à tous les canaux de communication et pas seulement aux signaux acoustiques ; par exemple le feuillage agité par le vent gêne la perception visuelle de signaux visuels – tels que saccades de la tête - envoyés par un lézard à ses congénères). Le bruitage est plus ou moins important selon les caractéristiques du signal émis (ex : des sons basse fréquence et peu modulés sont en forêt moins sensibles à l'absorption par la végétation). Des signaux chimiques rémanents ; des signaux présentant un fort rapport signal/bruit, un codage de l'information dans des paramètres résistants, une redondance de l'information sont des adaptations. Le choix des lieux, moments et modalités d'émission (ex. utilisation d'un terrier ou d'un trou comme caisse de résonance chez la courtilière ou certains amphibiens ; postes de chant chez oiseaux) sont d'autres adaptations possibles. Côté réception du signal, les capacités du système neurophysiologique à extraire un signal du bruit de fond entrent aussi en jeu. Certaines espèces jouent sur les modifications imposées par l'environnement à la structure des signaux pour cibler tel ou tel récepteur et éviter le « parasitage » de l'information (ex. les signaux pré-copulatoires se propagent souvent mal, attirant moins les compétiteurs ou prédateurs ; exemple = différence entre le chant territorial du grillon –très fort et portant loin- et le chant de cour –d'intensité faible et réserver au partenaire sexuel). Des exemples d'adaptations sont :
- l'adaptation des organes émetteurs et récepteurs aux contraintes physiques de l'environnement (ex. chaîne des osselets de l'oreille moyenne qui traite de la différence d'impédance air-eau).
- l'utilisation de signaux chimiques plus ou moins volatils, rémanents ou non, adaptée à leur rôle biologique.
- l'utilisation optimisée des caractéristiques de résonance d'un terrier ou d'un gîte pour en faire une caisse de résonance pour un chant ; ainsi le mâle d'une petite grenouille forestière de Bornéo (Metaphrynella sundana) est capable d'adapter précisément (après quelques essais) sa position, la force et le ton de son champ, selon le niveau de l'eau dans son trou pour utiliser au mieux la résonance de la cavité et mieux faire entendre son chant dans le brouhaha de la jungle (distinctement jusqu'à une cinquantaine de mètres dans ce cas)[4] - [5]. De même certains oiseaux forestiers adaptent leur chant à la densité du feuillage de leur environnement, selon la hauteur où ils sont actifs[6]
Communication et évolution des espèces
La seconde question liant communication et évolution est de savoir en quoi les communications jouent un rôle dans l'évolution des espèces. De ce point de vue, il n'y a communication que si le processus d'échange d'information représente un bénéfice sur le plan de la survie de l'individu, du groupe, de l'espèce.
Dire que le premier rôle des signaux de communication est de permettre la reconnaissance de l'espèce lors de la rencontre des deux sexes est une hypothèse raisonnable (s'il n'y pas de reconnaissance du signal, l'appariement mâle-femelle n'a pas lieu). Les signaux de communication peuvent alors facilement jouer le rôle de barrière prézygotique.
L'existence de populations allopatriques entraîne la dérive des signaux de communication (apparition de « dialectes », soit liés à une dérive génétique, ex. drosophile des îles Hawaï ; soit culturelle, ex. oiseaux chanteurs, à la suite d'« erreurs » de copie du chant du père). Cette dérive, si elle est assez importante, peut amener l'apparition d'une barrière prézygotique plus ou moins étanche (les signaux mâles d'une population n'ont plus – ou moins - d'effet attracteur sur les femelles de l'autre population). Il est à noter qu'une dérive « culturelle » des signaux de communication peut être très rapide, beaucoup plus rapide qu'une dérive génétique.
Une autre possibilité est que la modification des signaux de communication soit un effet secondaire d'une différenciation adaptative entre populations. Ex. cas des pinsons de Darwin : l'évolution du bec (liée aux contraintes alimentaires) entraîne des modifications du tractus vocal avec pour conséquence des modifications des signaux sonores produits, pouvant ainsi amener une barrière prézygotique entre des populations de la même espèce (Rq. par ailleurs, les pinsons de Darwin ont connu et connaissent encore le premier cas : dérive des signaux à la suite d'une allopatrie).
Cas des primates non-humains
La « plasticité vocale » des primates non-humains (la capacité d'apprendre des sons nouveaux ou de modifier la structure des appels en réponse à des variables sociales ou environnementales) a d'abord été considérée comme limitée bien que les oiseaux chanteurs, les baleines, les dauphins et quelques autres mammifères aient cette capacité[7]. Certaines études ont conclu à l'absence de « trame vocale » permettant de produire les sons de la parole humaine chez les primates non humains et que les primates ne peuvent se relayer (aspect de la conversation humaine)[7]. Mais ces trois points ont été récemment contestés par des auteurs qui considèrent que les primates non humains restent des modèles importants pour comprendre l'évolution de la parole et du langage[7].
Communication animale et applications
On envisage ici deux domaines où les connaissances acquises concernant les communications animales permettent d'envisager une action sur les processus naturels, soit sur le plan de la protection de la nature, soit sur le plan de la lutte contre les nuisances provoquées par certaines espèces.
Biologie de la conservation
Le fait que les signaux de communication soient signés, c'est-à-dire qu'ils puissent porter l'identité de l'espèce, d'une population, voire des individus, permet d'envisager l'identification — et donc le suivi — des animaux sur la base de ces signaux.
- Suivi acoustique (méthode non invasive ; utilisée pour suivi des migrations d'oiseaux, d'espèces menacées). Ces méthodes sont employées de façon routinière en mer pour le suivi des cétacés (espèces non facilement observables, et chaque espèce de baleines émet un chant qui lui est propre).
- Mesure de la biodiversité (forêt tropicale, océan). Un enregistrement sonore en forêt tropicale ou ailleurs permet de recenser les espèces présentes. Si cette méthode est depuis longtemps employée pour recenser les oiseaux par exemple (méthode des « points d'écoute »), des approches automatisées sont en développement afin d'être applicables sur le long terme et dans des environnements difficilement accessibles.
Gestion des risques biotiques
On détourne le rôle biologique de certains signaux (cris de détresse, phéromones d'attraction sexuelles) au profit des activités humaines.
Un premier exemple est celui des effaroucheurs acoustiques positionnés sur la plupart des aéroports européens pour faire face au « péril aviaire » (le danger que représentent les oiseaux lors des décollages et atterrissage des avions). Ils sont également employés par certaines piscicultures et au sein d'urbanisations lorsqu'elles subissent des invasions d'oiseaux (étourneaux par exemple). Les effaroucheurs peuvent envoyer des cris de synthèse, imitant les caractéristiques de cris de détresse d'oiseaux (on utilise le fait que ces cris ont une valeur interspécifique).
Un second exemple est celui des pièges phéromonaux pour le contrôle des ravageurs de culture.
Question du langage animal
La question de la différence entre communication animale et langage humain est âprement discutée. Aujourd'hui, il se dégage un consensus pour dire que le langage humain est plus complexe que n'importe quelle forme de communication animale.
Afin d'étudier cette question, on a d'abord cherché à savoir quelles étaient les propriétés fondamentales de la langue. On peut notamment dégager :
- L'arbitraire du signe : il n'y a pas de lien entre un son et sa signification (cet arbitraire est réduit dans le cas des onomatopées).
- La transmission culturelle : l'utilisation d'une langue est transmise de génération en génération spontanément.
- La discrétivité : le langage est composé d'unités discrètes qui, combinées, créent la signification.
- Le déplacement : le langage peut être utilisé pour véhiculer des idées sur des choses qui ne sont pas présentes spatialement ou temporellement[8].
- La dualité.
- La métalinguistique : nous sommes capables de parler du langage lui-même.
- La productivité : à partir d'un nombre fini de mots, nous sommes capables de créer une infinité de phrases.
En 1952, le linguiste Émile Benveniste publie un article dans Diogène, intitulé « Communication animale et langage humain »[9]. Cet article est une réaction aux découvertes fondamentales (et très modernes pour l'époque) du zoologue Karl von Frisch sur la communication des abeilles. Il montrait que les abeilles ouvrières pouvaient indiquer à leurs congénères la direction, la distance mais aussi la qualité d'une source de nourriture par rapport à la ruche. Émile Benveniste analyse les différences entre le « langage » des abeilles et celui des humains. L'enjeu est considérable, puisqu'il s'agit de savoir si l'être humain a, d'un point de vue linguistique, un semblable sur terre : « [...] pour la première fois nous pouvons nous représenter le fonctionnement d'un « langage » animal. Il peut être utile de marquer brièvement en quoi il est ou il n'est pas un langage, et comment ces observations sur les abeilles aident à définir, par ressemblance ou par contraste, le langage humain. »
Les différences relevées par Benveniste sont les suivantes :
- « Le message des abeilles consiste entièrement dans la danse, sans intervention d'un appareil « vocal », alors qu'il n'y a pas de message sans voix » ;
- « Le message des abeilles n'appelle aucune réponse de l'entourage, sinon une certaine conduite, qui n'est pas une réponse » : le dialogue (et l'intersubjectivité qui lui est essentielle) est une condition du langage ;
- « Le message d'une abeille ne peut être reproduit par d'autres » : la fonction métalinguistique du langage est ignorée des abeilles ;
- « Dans le langage humain, le symbole en général ne configure pas les données de l'expérience » : le signe chez les abeilles n'est pas arbitraire puisqu'il y a correspondance entre la quantité de danses et la distance de la fleur ;
- « Le message des abeilles ne se laisse pas analyser » : il n'y a pas de morphologie du « langage » des abeilles. Dans le langage humain, « un nombre assez réduit de morphèmes permet un nombre considérable de combinaisons, d'où naît la variété du langage humain, qui est capacité de tout dire ».
Selon ces critères, la danse des abeilles n'est donc pas un langage, mais seulement une capacité à communiquer. Ces conditions nécessaires sont néanmoins critiquables, en particulier la première, le système vocal n'étant qu'un système de transmission d'information comme un autre. La langue des signes, bien que n'utilisant pas la voix humaine, est bel et bien un langage doté d'une syntaxe et d'une sémantique. Benveniste conclut l'article en remarquant : « Ce n'est pas le moindre intérêt des découvertes de Karl von Frisch, outre les révélations qu'elles nous apportent sur le monde des insectes, que d'éclairer indirectement les conditions du langage humain et du symbolisme qu'il suppose ».
Les exemples les plus souvent étudiés sont :
- La danse des abeilles
- La vocalisation des oiseaux
- Le chant des baleines
- La communication des singes
- Les sons du dauphin et des autres cétacés (encore mal compris)
- Les cris, grondements et infrasons des éléphants
- Les phéromones, chez la plupart des animaux terrestres
Exemples de tentatives d'apprentissage du langage humain chez des animaux
- Washoe : première tentative d'apprentissage de l'American Sign Language chez un chimpanzé ;
- Nim Chimpsky : deuxième tentative ;
- Kanzi : bonobo ayant des aptitudes au langage ;
- Koko : gorille parlant la langue des signes ;
- Alex : perroquet ayant un vocabulaire étendu et comprenant la notion de zéro ;
- Hans le Malin : cheval dont on a cru qu'il savait compter (ce qui s'est révélé faux) ;
- Batyr : éléphant.
Cet article a envisagé la communication intraspécifique à diverses échelles d'explication et d'observation, tant du point de vue proximal (production des signaux, codage de l'information, transmission et réception des signaux, rôles biologiques de la communication), qu'évolutif (adaptations des communications aux contraintes biologiques et environnementales, rôle des communications dans l'évolution des espèces).
L'environnement physique détermine considérablement la nature et la structure des signaux employés, avec en conséquence une relative homogénéité dans le Règne animal. Les signaux sonores et visuels sont employés par des groupes nombreux et phylogénétiquement très éloignés ; et les signaux chimiques semblent employés par tous. Si l'on excepte les signaux électriques utilisés par certains poissons, aucun groupe animal ne semble avoir développé un canal de communication particulièrement original par rapport aux autres ; l'homme fournit bien sûr une exception notable avec des signaux de communication nouveaux (ex. utilisation d'ondes électromagnétiques) développés dans un cadre technologique (notion de phénotype étendu). Par ailleurs, la communication animale est un exemple de la non-linéarité de l'évolution phylogénétique. Les vertébrés par exemple ont développé des systèmes particulièrement complexes de communication (signalisation multi-modale, apprentissage, …) chez des lignées aussi différentes que les oiseaux et les primates. La diversité des échanges d'information, confirme l'importance de la communication surtout chez les animaux à socialité élaborée. La communication animale est un processus dynamique, connaissant une évolution liée à des contraintes et des bénéfices. Ainsi, des bases théoriques comme les observations montrent que les animaux tentent de résoudre les conflits avec le plus faible coût énergétique possible : les signaux de communication participent de cette stratégie.
Les questions posées par Tinbergen restent d'actualité. Sur le plan proximal, la comparaison des systèmes de communication, des modalités de codage de l'information, des rôles des signaux dans la vie quotidienne des animaux n'ont pas fini de livrer leurs secrets. En particulier, le fonctionnement des réseaux de communication où les protagonistes sont tour à tour émetteurs, récepteurs et parasites de l'information, commence tout juste d'être compris. Par ailleurs, l'étude du comportement animal ne saurait être dissociée de ses bases neurophysiologiques. Le fonctionnement cérébral in vivo et sur des animaux non-anesthésiés (électrophysiologiques notamment) commence à pouvoir être étudié en situation, quand l'animal échange des informations avec des congénères, dont grâce à l'imagerie fonctionnelle (IRMf, approches biophotoniques). Sur le plan des principes évolutifs, la théorisation sur la communication animale a récemment avancé, dont en adoptant des concepts issus de l'économie et de la théorie des jeux. Mais la théorie doit être confrontée aux réalités expérimentales. Et le rôle des communications animales dans les processus d'évolution des êtres vivants n'a été qu'effleuré. Concernant notre espèce, une interrogation majeure demeure celle du langage. Non qu'il faille le placer en haut d'une hypothétique échelle de valeur –chaque système de communication possède ses propres spécificités-, mais l'envisager en tant que processus intrinsèquement lié aux liens sociaux qui unissent les humains permettrait d'apporter un éclairage sur ses modalités. La récursivité (le fait de pouvoir construire une infinité de phrases avec un nombre limité de mots) est-elle propre à notre espèce ? Décèle-t-on des prérequis à cette aptitude chez d'autres Primates ? Enfin, les concepts élaborés dans le cadre de l'étude des communications animales et de la théorie de l'information ont une portée générale, pouvant éclairer l'analyse d'autres systèmes naturels (ex. relations plantes-pollinisateurs, hôte-parasite, voire aux échelles intraorganisme et intracellulaire).
Notes et références
- Cet article est partiellement ou en totalité issu de l'article intitulé « Zoosémiotique » (voir la liste des auteurs).
- « Cours de philosophie sur le langage. », sur mapage.noos.fr (consulté le )
- Liz Langley, « Comment fonctionne l’écholocalisation ? », sur National Geographic, (consulté le )
- Jean-François Dortier (dir.), La Communication. Des relations interpersonnelles aux réseaux sociaux, Éditions Sciences Humaines, coll. « Synthèse », , 398 p. (lire en ligne), « La communication animale »
- Clarke t (2016) Frogs tune call to hole ; Good vibrations aid the amphibian mating game.; News, Field Museum of Natural History, Chicago
- Lardner, B. & bin Lakim, M. Tree-hole frogs exploit resonance effects. Nature, 420, 475, (2002).
- Whitfield J (2001), News : Perch affects birds' pitch Birdsong makes best use of forest acoustics ; mis en ligne le 28 Décembre 2001 | Nature | doi:10.1038/news011227-9
- Charles T. Snowdon (2017), Learning from monkey “talk” ; 17 Mar 2017: Vol.355, N°6330, pp. ; 1120-1122 ; DOI: 10.1126/science.aam7443 (résumé)
- Anne Reboul, Similarités et différences entre la causalité humaine et non humaine, 2005
- Article qui fait partie de son ouvrage Problèmes de linguistique générale, Gallimard, Paris, 1966, (ISBN 2 07 029338 6).
Annexes
Articles connexes
Liens externes
- Équipes de recherche sur la communication animale
- The BioAcoustics Team (Université Paris XI & Université de Saint-Etienne)
- Ethologie Animale et humaine (Université Rennes I)
- Ethologie et Cognition Comparée (Université Paris X)
- Société savante
- Revue
- Exposé
- [vidéo] UniversiteDeLyon, Langage humain et communication animale : Quelles similitudes, quelles différences ? sur YouTube