Conflit parents-progéniture
Le terme de conflit parents-progéniture fut introduit en 1974 par Robert Trivers. Il est utilisé en écologie évolutive pour décrire les différences d'intérêts entre les parents et leur progéniture dans l'investissement parental optimal. Ce concept découle d'une vision de la sélection naturelle au niveau du gène et de l'individu.
La théorie de Trivers[1]
Historique de la notion
Sir Ronald Fisher avait déjà évoqué le concept de dépense parentale (c'est-à-dire la part de ressource parentale, temps ou énergie, investie dans les soins parentaux apportés à la progéniture) lorsque Trivers introduisit en 1972 le terme d’investissement parental comme unité fondamentale pour mesurer toutes les ressources qu’un parent investit sur sa progéniture.
En 1974 Trivers publia son article "Parent-offspring conflict".
Le conflit parents-progéniture
En écologie évolutive, les relations entre les parents et leur progéniture peuvent être observées du point de vue des parents mais également des enfants. L'investissement parental est défini comme étant tout ce qui est fait par le parent afin d'augmenter les chances de survie de son enfant, mais qui diminue la capacité du parent à investir dans d'autres enfants. Il est donc présumé que les parents auraient tendance à fournir un investissement qui permettra de maximiser le nombre de petits qui survivront, tandis que la progéniture aurait tendance à maximiser son succès reproductif en demandant davantage d'investissement de la part des parents.
Le conflit qui est donc établi entre parents et progéniture, au contraire de celui observé entre des individus sans lien de parenté, est caractérisé par les relations génétiques très proches entre parents et enfants.
Un enfant aurait tendance à exiger plus d'investissement parental que celui que le parent lui donne (comme résultat de la sélection naturelle). Mais ceci aura pour conséquence de diminuer le nombre de futurs enfants de ce parent qui pourront survivre. Afin de définir le coût maximal qu'un enfant pourrait infliger à son parent, sans que son comportement soit contre-sélectionné, c'est-à-dire sans que le coût en mortalité de ces frères et sœurs soit supérieur à son bénéfice en termes de succès reproductif, on doit prendre en compte son degré d'apparentement (corrélation génétique) avec ses frères et sœurs.
Selon les travaux de William Donald Hamilton (1964), un comportement altruiste, c’est-à-dire un comportement qui nuit à l'individu qui l'accomplit et profite à un autre individu bénéficiaire, en termes de succès reproductif, peut être sélectionné uniquement si le bénéficiaire de ce comportement est suffisamment apparenté génétiquement à l'altruiste. Plus concrètement, le rapport bénéfice/coût (B/C) de ce comportement doit être supérieur à l'inverse du degré d'apparentement (1/r) entre les deux individus. Le degré d'apparentement est de 0.5 entre parents et enfants, 0.5 entre vrais frères et sœurs et 0.25 entre demi-frères et sœurs.
Le comportement altruiste sera sélectionné si :
| rB > C ≥ B/C > 1/r |
On peut donc appliquer la théorie de Hamilton aux relations parents–progéniture, en considérant des comportements altruistes de la part des parents envers leurs enfants, mais aussi entre les différents enfants.
Le conflit pour la continuation de l'investissement parental
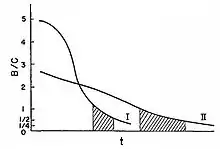
Afin d'illustrer ce type de conflit on peut considérer l'exemple d'un mammifère nouveau-né allaité par sa mère.
Le bénéfice de l'allaitement sur le nouveau-né en termes de chances de survie est énorme, tandis que le coût pour la mère en termes de capacité à produire d'autres nouveau-nés est très faible. Au fil du temps et comme le petit devient de plus en plus capable de se nourrir par lui-même, le bénéfice que l'allaitement lui fournit diminue et le coût pour la mère augmente (en fonction par exemple de la taille du petit). À un certain point, ce coût pour la mère va être supérieur au bénéfice du petit.
À partir de ce moment, la mère devrait arrêter l'allaitement pour ne pas diminuer le succès reproductif de sa future progéniture. Comme le petit est partiellement apparenté à ses futurs frères et sœurs (degré de 0.5 si ceux-là sont issus du même père et de la même mère) mais complètement apparenté à lui-même (degré de 1), il devrait vouloir être allaité jusqu'à ce que le rapport bénéfice/coût soit de 1/2, c'est-à-dire jusqu'à ce que le coût pour sa mère soit double par rapport au bénéfice pour lui. Le petit est donc sélectionné pour demander deux fois plus de nourriture que n'en recevront ses futurs frères et sœurs, puisqu'il est apparenté à lui-même deux fois plus qu'à eux. Au-delà de cette valeur (1/2), la sélection naturelle s'oppose à l'allaitement autant pour la mère que pour le petit.
On peut donc imaginer une période de temps (pour un rapport bénéfice/coût compris entre 1/2 < B/C < 1) durant laquelle on aura un conflit entre la mère et le petit concernant l'arrêt ou la continuation de l'allaitement. La sélection naturelle agissant sur la mère est favorable à l'arrêt de l'investissement parental (le rapport étant de 1, son comportement altruiste ne peut plus être sélectionné), tandis que la sélection agissant sur le petit est favorable à la continuation de cet investissement.
Ce type de conflit est notamment observé chez de nombreuses espèces d'oiseaux. Chez le Busard cendré par exemple, (Circus pygargus)[2] la période de conflit parents-progéniture commence après le premier envol des petits, lorsque ceux-ci ne sont pas encore complètement autonomes et ne peuvent subvenir à leurs besoins. Lors de cette phase qui annonce la fin de la dépendance des petits, l'apport de proies par les parents diminue graduellement, et notamment dès que les petits commencent à suffisamment bien voler pour tenter de chasser. Pourtant, les enfants ont des succès de capture encore modérés. Les actes agressifs des jeunes vers leurs parents deviennent alors fréquents et augmentent avec la diminution d'apport de nourriture par les parents. Cela suggère que cette diminution n'est pas due à un désintérêt des jeunes pour les proies mais à une modification du comportement des parents. Ceux-ci semblent donc capables de contrôler la fin du sevrage. Les petits de leur côté, essaient d'obtenir un maximum de nourriture de la part de leurs parents, mais réussissent uniquement lors des périodes où la nourriture est abondante dans le milieu. Par contre, quand la nourriture n'est pas abondante, la durée de la période d'investissement parental est quasiment sous le contrôle absolu des parents. Ceux-ci tendent quand même à la prolonger durant ces périodes, puisque les petits ont été relativement mal nourris.
Le conflit sur la quantité d'investissement parental
Le conflit pour la continuation de l'investissement parental peut être considéré comme étant simplement un cas particulier du conflit qui existe pendant toute la durée de cet investissement, concernant la quantité de celui-ci, entre le parent et son enfant. Et, lors du sevrage, ce type de conflit va se transformer en conflit concernant l'arrêt ou pas de cet investissement.
À tout moment pendant la durée de l'investissement parental, il est attendu que mère et petit soient en désaccord concernant la quantité de l'investissement fourni: la femelle est sélectionnée pour fournir une quantité d'investissement inférieure à celle que l'enfant est sélectionné pour demander.
Types de conflits prénataux
Sexe-ratio:
Selon Trivers, un enfant potentiel peut être, sous certaines conditions, en désaccord avec ses parents concernant son futur sexe. Trivers considère qu'on ne peut pas affirmer de façon certaine qu'un enfant n'est pas capable d'influencer son propre sexe. Il fait donc l'hypothèse que les sexe-ratios observés dans la nature reflètent potentiellement le sexe préférentiel de chaque enfant, mais également celui désiré par le parent considéré. Le parent serait donc sélectionné pour produire préférentiellement le sexe qui lui demanderait le moins d'investissement (sexe non limitant), tandis que l'enfant serait sélectionné pour devenir le sexe qui recevrait le plus d'investissement parental (sexe limitant) et donc qui aurait un meilleur succès reproductif (car ayant reçu plus de ressources).
Autres traits d'histoire de vie:
Certains auteurs ont testé des hypothèses de conflits concernant différents traits d'histoire de vie. Notamment chez les mammifères on peut supposer que le fœtus est capable potentiellement d'influencer, via divers signaux, des caractères comme la taille de la portée ou son poids à la naissance.

Par exemple, des études faites sur le Mouton de Soay (Ovis aries)[3] ont permis d'analyser la taille des portées, ainsi que le poids des petits à la naissance. Les conclusions montrent la présence d'un conflit entre mère et enfants concernant ces deux traits d'histoire de vie. La sélection agissant sur la mère favorise la naissance de plusieurs petits de relativement petite taille. Au contraire, la sélection agissant sur l'enfant favorise la naissance d'un seul petit de relativement grande taille. La taille optimale des portées et le poids optimal des nouveau-nés diffèrent entre mère et petits. Cela suggère l'existence probable d'un compromis, ou trade-off, entre taille de portée et poids à la naissance.
On peut penser que ce genre de conflit serait moins probable chez des groupes tels que les sauropsidés, où les petits se développent dans des œufs. Cependant, il existe des études sur ce sujet et au terme desquelles ces hypothèses ont été traitées avec des résultats positifs.
L'enfant en tant que manipulateur psychologique
La présence de conflits entre un enfant et son parent force l'enfant à entrer en conflit avec celui-ci. Mais l'enfant n'est pas en mesure d'exiger physiquement plus d'investissement de la part de son parent. Étant plus petit et moins expérimenté, il se retrouve en désavantage envers son parent qui contrôle les ressources d'intérêt. Il va donc mettre en place des techniques de manipulation psychologique pour l'influencer et induire davantage d'investissement de sa part.
L'enfant possédant une meilleure connaissance de ses propres besoins que son parent, la sélection naturelle favorise la mise en place d'un système de signaux entre eux qui permet d'informer le parent de sa condition physique. Si, par exemple, l'enfant a faim, ou s'il se trouve en danger, il émet un signal sonore ou chimique, et le parent agit de façon appropriée. Les deux parties profitent de ce système, mais une fois qu'il est mis en place, l'enfant peut commencer à l'utiliser hors contexte (faim, danger ou autre). Il peut, par exemple, commencer à émettre un signal lorsqu'il n'a pas vraiment faim mais qu'il veut juste plus de nourriture que celle que le parent lui donne. Plus généralement, il peut utiliser ce système de signaux lors du conflit avec son parent, afin d'exiger davantage d'investissement parental que celui que son parent fournit.
Il est bien sûr également attendu que la sélection naturelle favorisera une capacité parentale à différencier les deux utilisations du système de signaux établi, et à contrôler son investissement. Mais même dans ce cas, l'enfant peut toujours l'utiliser afin d'induire plus d'investissement. Par exemple, dans une espèce pour laquelle le silence est important afin d'éviter les prédateurs, les signaux sonores de l'enfant obligent le parent à répondre, même s'il sait que son besoin n'est pas réel.
L'approche de Richard Dawkins[4]
Conflit du point de vue des parents (en particulier de la mère)
Chez les mammifères et plus généralement chez les vertébrés, une mère n’a génétiquement aucune raison de faire de préférence sur l’un ou l’autre de ses petits puisqu'elle partage le même degré d'apparentement avec chacun d’eux : 0.5 (à noter que le degré d'apparentement maximum est de 1, celui entre un individu et lui-même ou son clone ou encore son vrai jumeau). Or on observe, dans la nature, des cas où la mère a des préférences, c'est-à-dire qu'elle opère une répartition inégale de ses ressources (nourriture, protection, éducation, ...) entre ses différents petits.
Cas du petit le plus faible :
À un instant donné, le plus jeune petit d’une femelle, qui est a priori aussi le plus faible, nécessite plus de ressources pour arriver à maturité que ses frères et sœurs plus grands et plus forts. Or c’est la maturité des petits qui importe aux parents car ils pourront alors assurer la survie et la dispersion de leurs gènes. Il y aura donc un compromis sur les ressources allouées par la mère sur ses différents enfants en fonction de leur âge, face à une situation de plus ou moins grande gravité :
- S’il est question de vie ou de mort, la mère préfèrera protéger le (ou les) plus grand(s) car elle aura moins de ressources à fournir pour l’(les) amener à maturité. Un exemple particulier observé chez de nombreux oiseaux (avec la théorie de Lack, de 1954, sur la taille de la couvée) : une jeune femelle inexpérimentée va pondre un œuf de plus pour tester son optimal de ponte. S’il s’avère qu'elle ne peut pas assumer un petit de plus, elle ne s’occupera pas du plus faible de la couvée et le laissera mourir pour ne pas dépenser plus d’énergie avec lui que ce qu'elle a déjà dépensé à la constitution du jaune d’œuf (cas particulier de vitellus), voire, dans certains cas, le laissera se faire manger par ses frères et sœurs et/ou par elle-même afin de ne pas perdre cette énergie.
Ceci a, par exemple, été observé dans des études faites sur le Perce-oreille (Forficula auricularia)[5] - [6] démontrant l'existence de signaux chimiques dépendants de la condition physique des nymphes de cette espèce et ayant des effets sur l'investissement maternel. Plus concrètement, des signaux de bonne condition physique de la part des jeunes induisent la continuation de l'investissement maternel, tandis que des signaux de mauvaise condition physique induisent une agressivité de la part de la mère. On peut donc penser que la mère agit en fonction de l'intérêt de sa future progéniture ; si ses nymphes actuelles présentent des indices d'un futur succès reproductif important, elle prolonge son investissement, mais au contraire, si elles présentent des indices d'un faible succès reproductif, elle arrête l'investissement. Ces signaux chimiques sont probablement une stratégie qui a évolué afin de résoudre le conflit entre mère et nymphes concernant l'arrêt de l'investissement maternel.
- Si l'enjeu est moindre, elle préfèrera se concentrer de façon plus importante sur le plus petit de la couvée car les plus grands sont déjà à même de se débrouiller seuls, et sont beaucoup plus proches de la maturité.
Cette deuxième situation amène chez les mammifères un comportement de sevrage et de conflit tel que Trivers l'a décrit : le petit tout juste mature veut continuer de profiter des ressources de la mère alors que celle-ci a plus d'intérêts à se séparer de lui et à investir sur une nouvelle portée ou un nouveau petit. Le conflit (qui est également un trade-off) s’opère entre mère, enfant présent et enfant(s) à naître, pour le sevrage de l'enfant présent ainsi que pour la date de ce sevrage.
Autre cas de conflit :
Quand la mère ne peut plus avoir d’enfants, il peut être plus profitable pour sa fitness (valeur sélective) relative d’allouer ses ressources à un ou plusieurs autres parents moins proches (petits enfants, nièces, neveux, ...) qu'à son propre enfant déjà mature. Ce mécanisme évolutif aurait abouti au phénomène de ménopause chez la femme, conjointement au fait que, selon la théorie de Peter Medawar sur le vieillissement, une femme devient avec l’âge de moins en moins efficace pour élever des enfants. Donc l’espérance de vie d’un petit issu d’une mère âgée est inférieure à celle d'un petit issu d’une mère jeune.
Traduction en terme génique : « Lorsqu'une femme atteint l’âge où la probabilité moyenne pour que chacun de ses enfants atteigne l’âge adulte est à peine inférieure à celle qu'ont ses petits-enfants d’atteindre le même âge, tout gène [induisant le comportement] d’investir sur les petits-enfants plutôt que sur ses propres enfants, prospère ». (Le gène envahit le système : le pool génique) - Richard Dawkins.
Conflit du point de vue des enfants
Hypothèse de chantage du petit sur les parents, énoncé par A.Zahavi, selon laquelle les petits pourraient pousser des cris bien qu’ils aient reçu leur part de ressources par les parents. Ceci aurait pour effet d’attirer les prédateurs en signalant la position du nid, et donc forcerait les parents à donner plus que sa part au petit afin qu’il cesse ses cris.
Exemple : Le conflit lors du sevrage chez les babouins[7] est caractérisé par une compétition quotidienne qui peut durer pendant plusieurs mois. La mère résiste de plus en plus aux sollicitations du petit et réagit souvent en le rejetant violemment. Le petit de son côté intensifie ses efforts pour s’accrocher à sa mère et réclamer davantage d'allaitement. Ce rejet de la part de la mère se traduit chez le petit par des cris. Ces cris présentent un coût très important pour le parent et son enfant car cette espèce possède des comportements très silencieux qui sont adaptés à l'évitement des prédateurs. Ainsi les cris sont contre-sélectifs.
Traduction en terme génique : Les gènes induisant chez le bébé babouin des cris forts en réaction au comportement de la mère se sont répandus dans la population de babouins – dans le pool génique – car ils augmentaient la probabilité que le bébé reçoive une quantité d'investissement parental (nourriture ou autres) supplémentaire. Mais pourquoi, d’un point de vue évolutif, les mères de cette espèce répondent à ce comportement (cris) ? Parce que les gènes induisant chez les femelles le comportement de nourrir plus leur(s) petit(s) babouin(s) en réponse à leur(s) cris se sont répandus dans le pool génique. Et ceci car les individus possédant ces gènes auraient une meilleure fitness que ceux ne les possédant pas et qui ne nourrissaient donc pas plus le petit babouin en réponse à ses cris. Ce qui aurait pour conséquence d’attirer les prédateurs qui pourraient manger la progéniture et/ou la mère. Leurs gènes ne pouvant se transmettre à la génération suivante, ces individus auraient alors décliné dans le pool génique laissant la place aux gènes induisant une réponse positive aux cris.
Conflit sur la taille de la couvée :
On a vu précédemment le comportement possible d’une jeune mère cherchant à déterminer son optimum de taille de ponte. Mais le choix qu'elle va faire peut ne pas être en accord avec la taille optimale de ponte du point de vue de sa progéniture. En effet, lorsqu'on se place du point de vue d’un petit, l’optimum de ponte est très probablement inférieur à celui que détermine la mère car le petit veut plus que la part des soins qui devrait lui être attribuée. Mais comment fait un petit, pour modifier la taille de ponte dont il est issu ? Il peut développer un comportement fratricide (c'est-à-dire avoir un gène qui confère un comportement fratricide à un jeune âge). Si nous prenons l’exemple d’une couvée de 5 petits, et que les ressources sont distribuées équitablement aux petits par la mère : si vous êtes l'un des petits, vous obtiendrez 1/5 des ressources de votre mère. Si vous tuez un de vos frères ou sœurs, vous recevrez 1/4 des ressources, si vous en tuez un deuxième vous en obtiendrez 1/3,... Car même si la moitié de vos gènes se trouve chez vos frères et sœurs apparentés à vous à 50 % (ce qui conduit habituellement à l’altruisme), vous êtes apparenté avec vous-même à 100 %. Mais ce comportement semblerait ne pas pouvoir aboutir à une ESS (stratégie évolutivement stable) car il ne s’observe quasiment pas dans la nature à un point aussi extrême.
Remarque

Richard Dawkins utilise à plusieurs reprises l'exemple de parasitisme de couvée avec le coucou. Il est important de garder à l'esprit que cet exemple vise à illustrer les différentes stratégies, exagérées jusqu'à un point extrême, pour bien les saisir. Il s'agit en réalité, dans le cas du coucou, de stratégies de parasitisme et non de réels conflits de génération. Pour cette raison, ne sont donnés ci-dessus que des exemples tirés d'études plus récentes et illustrant cette vision.
De plus, le point le plus important à retenir n'est pas tant l'ensemble des cas particuliers énoncés et illustrés ici, mais plutôt la démarche réflexive à effectuer en amont: le fait de ramener chacun des traits comportementaux individuels à une sélection naturelle au niveau du gène ou de la combinaison de gènes qui contrôle ledit comportement.
Théorie du "poussin appétissant"[8] - [9]
.jpg.webp)
La théorie du poussin appétissant ou "tasty chick hypothesis", est une théorie élaborée dans le domaine de l'évolution des interactions hôtes-parasites. Elle rend compte de l'effet de dilution de la charge parasitaire entre des individus qui cherchent le contact avec d'autres individus moins (ecto)parasités afin qu'une partie de leurs ectoparasites passe de leur corps à un autre. Ici la mère alloue moins de ressources (énergétiques ou immunitaires) à l'un des œufs de sa ponte ou bien pond un œuf supplémentaire, mais retardé par rapport au reste de la ponte. Ceci aura pour corollaire de rendre ce dernier petit plus jeune et donc plus fragile vis-à-vis de ses frères et sœurs plus forts, déjà âgés de quelques jours. Ainsi, alors que ce petit veut sa part des ressources comme les autres membres de sa fratrie, la mère le maintient dans un état de faiblesse vis-à-vis des ectoparasites qui migreront préférentiellement de ses frères et sœurs vers lui plutôt que l'inverse. Ceci assure donc une meilleure santé aux plus grands en leur assurant une dilution de leur quantité d'ectoparasites, et augmente ainsi indirectement la fitness de la mère. Il découle donc de cette stratégie évolutive de la mère, un conflit cruel avec l'un de ses petits.
Conflits parents-progéniture chez les végétaux
On pourrait parler de conflit parent-progéniture chez certaines plantes[10] - [11]. Effectivement, l'allocation des ressources d'une plante dans les différentes graines qu'elle produit, ainsi que la taille de celles-ci, peuvent être le sujet d'un conflit entre parent et progéniture. La meilleure façon d'économiser les ressources maternelles (et donc de maximiser la fitness maternelle) est de produire un maximum de graines par fruit, de relativement petite taille. Au contraire, pour maximiser la fitness de la progéniture, les fruits devraient être plus gros qu'ils ne le sont et contenir moins de graines. De cette façon, la compétition entre les individus se minimise, autant avant qu'après la dissémination. Ce conflit pour les ressources se manifeste par le fratricide observé dans certaines espèces de plantes. Les embryons dominants, formés tôt, tuent les autres avec une substance chimique abortive. Chez le chêne, les ovules fécondés tôt vont empêcher la fécondation des autres en empêchant la formation du tube pollinique. Également, le degré d'apparentement entre les embryons joue un rôle important dans ce type de conflit, car ces phénomènes d'avortement d'embryons sont plus souvent observés chez des espèces qui présentent une fécondation croisée que chez celles qui pratiquent l'autofécondation.
Notes et références
- Trivers, R.L. 1974. Parent-offspring conflict. American Zoologist 14:247-262.
- Arroyo B.E., De Cornulier T. et Bretagnolle V. 2002. Parental investment and parent-offspring conflicts during the postfledging period in Montagu's harriers. Animal Behaviour 63:235-244.
- Wilson A.J. et al. 2005. Selection on mothers and offspring: whose phenotype is it and does it matter ?. Evolution 59:451-463.
- Dawkins R. 1976. Le gène égoïste. Chapitre VIII:171-192.
- Mas F. and Kölliker M. 2011. Differential effects of offspring condition-dependent signals on maternal care regulation in the European earwig. Behavioral Ecology and Sociobiology 65:341-349.
- Mas F., Haynes K.F.and Kölliker M. 2009. A chemical signal of offspring quality affects maternal care in a social insect. Proceedings of The Royal Society B: Biological Sciences 276(1668):2847-2853.
- Dario Maestripieri, August 2002, Parent–Offspring Conflict in Primates. International Journal of Primatology, Vol. 23, No. 4.
- Philippe Christe, Anders Pape Møller and Florentino de Lope, Immunocompetence and Nestling Survival in the House Martin: The Tasty Chick Hypothesis. Oikos Vol. 83, No. 1 (Oct., 1998), pp. 175-179
- Alexandre Roulin, Martin W. G. Brinkhof, Pierre Bize, Heinz Richner, Thomas W. Jungi, Christian Bavoux, Nicolas Boileau and Guy Burneleau, 2003. Which chick is tasty to parasites? The importance of host immunology vs. parasite life history. Journal of Animal Ecology 72, 75–81
- Shaanker, R.U., Ganeshaiah, K.N. and Bawa, K.S. 1988. Parent-offspring conflict, sibling rivalry, and brood size patterns in plants. Annual Review of Ecology and Systematics 19:177-205.
- Sadras V.O. and Denison R.F. 2009. Do plant parts compete for resources? An evolutionary viewpoint. New Phytologist 183(3):565-574.