Communication chez les plantes
En botanique, la communication chez les plantes, appelée aussi communication des plantes ou communication végétale n'est pas un acte réfléchi comme la communication humaine mais implique chez la plante émettrice une émission plastique et conditionnelle du signal (composé sémiochimique, signal électrique, sonore, hydraulique ou lumineux) en fonction de stimuli environnementaux, associée à une réponse rapide de l'organisme récepteur (micro-organismes, champignons, animaux, plante de la même espèce ou d'une autre espèce)[1].
Une des modalités est la communication inter-plantes ou signalétique des plantes, domaine de recherche prolifique qui a commencé en 1983 en Amérique du Nord.
Modes de communication
Communication souterraine versus communication aérienne
- Communication souterraine par les systèmes racinaires qui peuvent échanger des signaux via des anastomoses inter-individu[2] ou le réseau mycorhizien, établissant un système de « relation sociale ».
- Communication aérienne par des composés organiques volatils biosynthétisés au niveau de cellules spécialisées dans la sécrétion. Ces cellules sont situées soit au niveau des fleurs, et on parle alors de plante à parfum, soit au niveau de l’appareil végétatif, et on parle alors de plante aromatique. Ces composés sont principalement des terpénoïdes. Ce sont également des dérivés d’acides gras (principalement des substances volatiles des feuilles vertes de type hexénal (en)[3]), des composés aromatiques (famille des benzénoïdes, des phénylpropanoïdes auxquels appartiennent les coumarines et les flavonoïdes qui sont les substances les plus communes dans le parfum de fleurs[4]), des composés azotés et des composés soufrés[5]. Parmi les 200 000 métabolites secondaires biosynthétisés par les plantes, plusieurs dizaines de milliers de ces substances allélochimiques ont été identifiés, la majorité consacrée à leur défense contre les herbivores[6]. Le profil chimique de différentes plantes est généralement différent et est souvent complètement unique pour cette plante individuellement[7].
Communication intra-plante versus communication inter-plantes
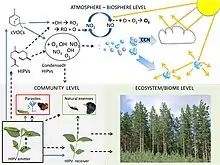
La communication végétale fait appel à la perception des plantes et à différents systèmes de transduction des signaux et de signalisation cellulaire qui impliquent généralement des kinases, des canaux ioniques calcium, potassium… et des facteurs de transcription[9].
Dans la communication intra-plante (ou communication interne des plantes), la perception des signaux de communication débute dans plusieurs voies. La principale, hormonale, peut utiliser les systèmes vasculaires (les signaux chimiques sont essentiellement des phytohormones de défense mais aussi impliquées dans la fermeture des stomates, avec par exemple le dessèchement des racines qui induit la production d'acide abscissique envoyé, via le xylème, en direction des feuilles)[10]. Les signaux électriques à propagation rapide et mettent en jeu des pompes dépendantes de l'ATP et une dépolarisation membranaire transitoire locale du potentiel électrique[11]. Des signaux hydrauliques induits par des blessures utilisent le système hydraulique (xylème et phloème)[12].
La communication inter-plantes ou signalétique des plante est un domaine de recherche qui a commencé en 1983 en Amérique du Nord avec les travaux pionniers[13] du zoologue Rhoades[alpha 1] puis des écologues Baldwin (en) & Schultz[alpha 2]. Bien que ces études initiales soient critiquées dans le monde scientifique[1], notamment[16] celle du koudou et l'acacia[alpha 3], leurs conclusions se répandent dans la culture populaire à travers les métaphores et expressions anthropomorphisantes[alpha 4] d'« arbres parlants », de « langage des plantes », de « sensibilité des plantes » et d'« intelligence des plantes » (« débat qui nécessiterait une solide clarification épistémologique du concept d'intelligence »)[21], relayées non seulement dans les pages du magazine Science, mais aussi dans la presse d'information générale du monde entier[22]. Depuis, les preuves de communication inter-plantes dans un contexte de défense ont été étayées surtout dans des laboratoires ventilés et plus rarement sur le terrain (avec notamment l'aulne glutineux, la sauge et le tabac qui comptent parmi les végétaux les plus étudiés)[23]. Cette communication utilise parfois des signaux sonores (perception des vibrations acoustiques émises par un prédateur)[24] et lumineux[13].
Ces preuves restent encore limitées, si bien que les scientifiques ne s'aventurent pas à généraliser le phénomène[25]. En effet, si la plante réceptrice tire bénéfice de cette communication (induction de défense contre l'herbivorie), l'avantage d'une valeur adaptative pour la plante émettrice reste à élucider car la plante réceptrice non affectée a plus de risque de la priver de lumière et des éléments nutritifs du sol. Dans ces conditions, le scénario le plus probable est que le signal chimique s'adresse à l'herbivore plus qu'à la plante voisine qui intercepte les composés volatils lui signalant la présence d’un ravageur. Il s'agirait plus d'une « écoute clandestine » que d'une véritable communication[26]. Les études sur le terrain confirment ce scénario : dans un contexte de défense, les composés organiques volatils émis par une plante ne sont perçus qu'à quelques dizaines de centimètres d'elle, cette signalisation ne bénéficiant à des plantes voisines que par chance[27]. Ainsi, la communication végétale relèverait plutôt de la soliloquie[28] et la communication aérienne serait avant tout une communication intra-plante, une défense optimale combinant une phase rapide de « priming[alpha 5] » qui met en œuvre l'élicitation de la phase suivante plus lente, un déploiement de la défense via des signaux relais vasculaires (émission d'hormones de défense se comptant en jours ou en semaines) qui régulent la résistance systémique induite[30]. Ces fonctions de défense via la communication racinaire est mise en doute par certains scientifiques dans la revue nature [31] - [32].
Fonctions
Les signaux chimiques des plantes (arbres, plantes à fleur, etc.) servent principalement de moyen de défense chimique contre les herbivores, fonction primordiale chez ces organismes sessiles. Ils participent aussi à leur lutte contre les microbes pathogènes par le phénomène de résistance systémique acquise[33]. Ils apportent à la fois une protection directe, en inhibant ou en repoussant les agressions ou agresseurs (renforcement des parois, synthèse de toxines) et une protection indirecte, en attirant les prédateurs des herbivores eux-mêmes (parasitoïdes, oiseaux insectivores). Ils peuvent aussi avoir un impact différentiel (attractif ou répulsif) sur les organismes qui interagissent avec elles (symbiontes, parasites, pollinisateurs)[34].
Les composés chimiques ont également un rôle dans les mécanismes d'assemblage de communautés[35].
Les signaux électriques sont produits en réponse à de nombreux stimuli (pluies acides, irradiation, choc froid, stress osmotique, attaque d'agents pathogènes)[11].
Applications
Une meilleure compréhension de cette communication peut être utilisée pour protéger les cultures contre les pathogènes biotrophes, nécrotrophes et saprotrophes[36].
Notes et références
Notes
- Les feuilles de saule sitka parasitées par des « chenilles à tente » synthétisent des tanins et composés phénoliques toxiques qui réduisent l'herbivorie. Rhoades suggère que les arbres attaqués envoient, par voie aérienne un message phéromonal d'avertissement aux arbres sains, via des composés organiques volatils[14].
- De jeunes plants de peupliers et d'érable à sucre dont le feuillage est partiellement endommagé augmentent la teneur en tanins et composés phénoliques toxiques de toutes leurs feuilles et, selon les auteurs de l'étude, induiraient à distance, via un signal gazeux la même réaction chez des plants non affectés[15].
- La communication entre des acacias sous la pression de l'herbivorie des koudous, a fait l'actualité, à grand renfort de communication assurée depuis la fin des années 1980 par Jean-Marie Pelt via notamment L'aventure des plantes, les livres de vulgarisation de Francis Hallé et La Vie secrète des arbres de Peter Wohlleben, avec l'anecdote sensationnaliste du koudou et l'acacia[17], histoire sujette à caution car non validée par une revue à comité de lecture[16] : dans les années 1980, des milliers de grands koudous élevés dans des game ranches du Transvaal pour les soustraire à une chasse excessive, meurent. En autopsiant ces animaux, le zoologiste Wouter van Hoven de l'université de Pretoria considère que leur mort est due à leur principale source d'alimentation dans ces ranches lors de la sécheresse hivernale, une espèce d'acacia qui, en réponse à l'herbivorie, augmente la concentration en tanins dans ses feuilles jusqu'à des niveaux létaux[18] - [19].
- Pour le botaniste Lucien Baillaud, l'emploi d'expressions anthropomorphisantes lors de la médiatisation de la vulgarisation scientifique contrainte pour retenir le public de sacrifier aux recettes de la simplification et du sensationnalisme, peut être utile pour faciliter la compréhension : « Ne méprisons pas l'anthropomorphisme s'il nous aide à nous exprimer[20] ».
- « Priming » ou amorçage, préparation des défenses des végétaux à la suite de leurs interactions avec des microorganismes de l’environnement. Cette phase rapide aérienne (quelques heures) via des composés organiques volatils leur permet de résister à des attaques ultérieures d'agents pathogènes[29].
Références
- (en) Richard Karban, « Plant Behaviour and Communication », Ecology Letters, vol. 11, no 7, , p. 727‐739 (DOI 10.1111/j.1461-0248.2008.01183.x)
- Les anastomoses entre individus de la même espèce permettent aux racines des arbres abattus en éclaircie de survivre et de s'intégrer au système racinaire d'arbres voisins vivants.
- (en) S. Allmann et I. T. Baldwin, « Insects Betray Themselves in Nature to Predators by Rapid Isomerization of Green Leaf Volatiles », Science, vol. 329, no 5995, , p. 1075-1078 (DOI 10.1126/science.1191634)
- (en) F.P. Schiestl et S. Dötterl, « The evolution of floral scent and insect chemical communication », Ecology Letters, vol. 13, no 5, , p. 643-656 (DOI 10.1111/j.1461-0248.2010.01451.x).
- Jacques Tassin, À quoi pensent les plantes ?, Odile Jacob, (lire en ligne), p. 4
- (en) Maarten J. Chrispeels, Sadava E. David, Plants, Genes, and Agriculture, Jones and Bartlett Publishers, , p. 343
- (en) Sarah Y. Dewhirst & John A. Pickett, « Production of semiochemical and allelobiotic agents as a consequence of aphid feeding », Chemoecology, vol. 20, , p. 89–96 (DOI 10.1007/s00049-009-0032-8)
- (en) Jarmo K. Holopainen & James D. Blande, « Where do herbivore-induced plant volatiles go? », Front. Plant, vol. 4, no 185, (DOI 10.3389/fpls.2013.00185)
- Lydie Suty, Les végétaux. Des symbioses pour mieux vivre, éditions Quæ, (lire en ligne), p. 14-15
- (en) Martin R. McAinsh, C. Brownlee & Alistair M. Hetherington, « Abscisic acid-induced elevation of guard cell cytosolic Ca2+ precedes stomatal closure », Nature, vol. 343, no 6254), , p. 186-188 (DOI 10.1038/343186a0)
- François Bouteau et Patrick Laurenti, « La neurobiologie végétale, une idée folle ? », Pour la science, no 101, , p. 38
- (en) Stefano Mancuso, « Hydraulic and electrical transmission of wound-induced signals in Vitis vinifera », Australian journal of plant physiology, vol. 26, no 1, , p. 55-61 (DOI 10.1071/PP98098)
- (en) Sudhir P. Singh, Santosh Kumar Upadhyay, Ashutosh Pandey, Sunil Kumar, Molecular Approaches in Plant Biology and Environmental Challenges, Springer Nature, (lire en ligne), p. 128.
- (en) Rhoades, David F. 1983. « Responses of Alder and Willow to Attack by Tent Caterpillars and Webworms: Evidence for Pheromonal Sensitivity of Willows ». In Plant Resistance to Insects, Symposium Series 208, édité par Paul A. Hedin, p. 55‐68. Washington
- (en) Ian T. Baldwin et Jack C. Schultz, « Rapid Changes in Tree Leaf Chemistry Induced by Damage: Evidence for Communication Between Plants », Science, vol. 221, no 4607, , p. 277‐279 (DOI 10.1126/science.221.4607.277)
- Adrien Delattre, « Le koudou et l’acacia : histoire et analyse critique d’une anecdote », sur tela-botanica.org, (consulté le )
- (en) Edward E. Farmer, Leaf Defence, OUP Oxford, , p. 87
- (en) Wouter van Hoven, « Trees’ secret warning system against browsers », Custos, vol. 13, no 5, , p. 11‐16
- (en) Wouter van Hoven, « Mortalities in Kudu (Tragelaphus Strepsiceros) Populations Related to Chemical Defence in Trees », Journal of African Zoology, vol. 105, no 2, , p. 141‐145
- Lucien Baillaud, « La végétalité: un réseau de déterminismes reliant structure, biologie et milieu de vie dans le deuxième royaume de la nature », Acta Botanica Gallica, vol. 154, no 2, , p. 153-201
- François Bouteau et Patrick Laurenti, « La neurobiologie végétale, une idée folle ? », Pour la science, no 101, , p. 37
- Daniel Chamovitz, La Plante et ses sens, Buchet/Chastel, (lire en ligne), p. 47.
- (en) Richard Karban, Ian T. Baldwin, Induced Responses to Herbivory, University of Chicago Press, , 330 p. (lire en ligne)
- (en) Body MJA, Neer WC, Vore C1, Lin CH, Vu DC, Schultz JC, Cocroft RB, Appel HM, « Caterpillar Chewing Vibrations Cause Changes in Plant Hormones and Volatile Emissions in Arabidopsis thaliana », Front Plant Sci., vol. 10, no 810, (DOI 10.3389/fpls.2019.00810)
- (en) Edward E. Farmer, Leaf Defence, OUP Oxford, (lire en ligne), p. 84
- (en) M. Heil et R. Karban, « Explaining Evolution of Plant Communication by Airborne Signals », Trends in Ecology and Evolution, vol. 25, no 3, , p. 137 (DOI 10.1016/j.tree.2009.09.010)
- (en) Guillaume Becard, How Plants Communicate with their Biotic Environment, Academic Press, (lire en ligne), p. 7
- (en) Martin Heil, Rosa M. Ádame Alvarez, « Short signalling distances make plant communication a soliloquy », Biology letters, vol. 6, no 6, , p. 843-845 (DOI 10.1098/rsbl.2010.044)
- (en) Deepak G. Panpatte, Yogeshvari K. Jhala, Rajababu V. Vyas, Harsha N. Shelat, Microorganisms for Green Revolution, Springer, (lire en ligne), p. 316
- (en) M. Heil, J. Ton, « Long-distance signalling in plant defence », Trends Plant Sci., vol. 13, no 6, , p. 264-272 (DOI 10.1016/j.tplants.2008.03.005)
- « Controverse sur la communication souterraine entre les arbres », Le Monde.fr, (lire en ligne, consulté le )
- (en) Justine Karst, Melanie D. Jones et Jason D. Hoeksema, « Positive citation bias and overinterpreted results lead to misinformation on common mycorrhizal networks in forests », Nature Ecology & Evolution, (ISSN 2397-334X, DOI 10.1038/s41559-023-01986-1, lire en ligne, consulté le )
- (en) W.E. Durrant & X. Dong, « Systemic acquired resistance », Annu. Rev. Phytopathol, vol. 42, , p. 185–209 (DOI 10.1146/annurev.phyto.42.040803.140421)
- Anne-Geneviève Bagnères, Martine Hossaert-Mckey, Ecologie chimique, ISTE Group, (lire en ligne), p. 48
- Christophe Drénou, « Uni par les racines », Forêt-Entreprise, vol. 153, , p. 34-38
- Sudhir P. Singh, op. cit., p.127
Voir aussi
Bibliographie
- (en) František Baluška, Stefano Mancuso, Dieter Volkmann, Communication in Plants: Neuronal Aspects of Plant Life, Springer, , 438 p. (lire en ligne)
- (en) František Baluška, Velemir Ninkovic, Plant Communication from an Ecological Perspective, Springer Science & Business Media, , 252 p. (lire en ligne)
- (en) Richard Karban, Plant Sensing and Communication, University of Chicago Press, , 240 p. (lire en ligne)