Homologie (évolution)
En biologie de l'évolution, une homologie désigne un lien évolutif entre deux traits (en général anatomiques) observés chez deux espèces différentes, qui est dû au fait que toutes deux l'ont hérité d'un ancêtre commun. Ces traits sont alors dits homologues. Ce peut être des caractères anatomiques, ou moléculaires (protéines homologues). Ce terme s'étend aussi aux séquences génétiques (de l'ADN).
Le terme homologie est un concept fondamental pour la biologie en ce qui concerne l’organisation d’un organisme. Il est aussi un sujet très flou et complexe car il est expliqué différemment selon la communauté scientifique. L’homologie a été identifiée, par Hall en 2000, comme l’un des cinq éléments le plus important dans le domaine de développement évolutif. Au niveau de la biologie évolutive du développement, le terme homologie est utilisé pour expliquer les structures similaires et répétitives d’un organisme à travers plusieurs générations. Selon Van Valen, l’homologie est la correspondance provoquée par une continuité de l’information pour des structures au sein d’un organisme ou entre des organismes. Le concept d’homologie est peu connu et peu développé par les biologistes, surtout au niveau moléculaire. Malgré, les nombreuses définitions décrivant l’homologie, il n’y en a encore aucune qui est accepté par tous les biologistes. Ne pas confondre avec la définition d’homologie du domaine en biologie comparative et évolutionnaire.
Par opposition, des traits analogues sont des traits similaires en forme mais apparus indépendamment au cours de l'histoire de la vie : par exemple les ailes des oiseaux, des chauves-souris, ou des insectes.
Niveau anatomique
Cas général chez des espèces différentes
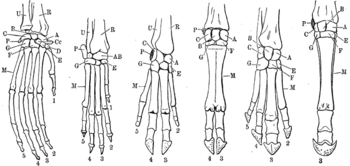
l’orang-outan, le chien, le cochon, les bovidés, le tapir et le cheval.
Certaines structures morphologiques ou anatomiques constitutives d’un organisme ont été acquises très précocement dans l'histoire de l'évolution des espèces et ont donné naissance à des structures modifiées mais dérivant toutes d’une structure initiale commune. À titre d'exemple, les membres des Vertébrés tétrapodes (ensemble des espèces possédant quatre membres) dérivent d’un membre ancestral et ont donné lieu à des variations (ailes des oiseaux, pattes des mammifères...) dans lesquelles il est possible de retrouver un patron initial (le membre chiridien).
En particulier, c'est le cas, par exemple, de la patte antérieure d'un cheval et de la nageoire pectorale d'un dauphin, qui ont comme ancêtre commun un mammifère (qui, il y a 80 millions d'années, ressemblait probablement à un chevrotain[1]), qui possédait comme ces deux animaux quatre membres et c'est le membre antérieur de ce mammifère primitif qui a donné la patte du cheval et la nageoire du dauphin.
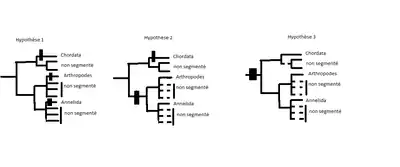
Présentement, en ce qui concerne l’évolution de la segmentation des bilatéraux, trois hypothèses sont postulées par plusieurs biologistes. La première n’a pas rapport à l’homologie, mais les deux dernières hypothèses démontrent une homologie. La deuxième postule qu’il y a une homologie des segmentations parmi les protostomiens et les chordés, mais que plusieurs des embranchements ont perdu cette segmentation. La dernière postule qu’il a une homologie parmi les bilatéraux mais que plus tard, encore plus d’embranchements ont perdu cette segmentation. Malgré les similarités retrouvées dans les divers organismes reliés, il n’y a pas assez de preuves et recherches pour conclure l’homologie des segmentations. Par exemple, dans l’évaluation de la deuxième hypothèse, le gène engrailed, connu pour exprimer les bandes segmentaires chez les arthropodes, a été considéré comme homologue chez les annélides et les arthropodes. Son expression chez les annélides semble jouer un rôle dans la segmentation, mais vu qu’il est exprimé après la séparation du primordium ganglionnaire, son rôle dans le processus de segmentation est douteux. Dans le cas de l’évaluation de la troisième hypothèse, le gène her1 retrouvé chez les poissons-zèbres est présumé d’être l’orthologue du gène pair-rule (pair-rule gene) hairy, responsable du développement des segments embryonnaires chez les insectes, car il imite son patron d’expression. Ceci suggère que l’ancêtre commun des protostomiens et deutérostomiens était segmenté, ainsi les segments de toutes les bilatéraux sont homologues. Malheureusement, il n’est pas encore certain si le gène her1 des poissons-zèbres s’agit actuellement de l’orthologue du gène pair-rule hairy.
Autres exemples :
- l'aile de la chauve-souris est homologue à la patte antérieure de la taupe ;
- malgré la différence de taille, le squelette du colibri est homologue à celui de la baleine[2] ;
- la vessie natatoire des Poissons est homologue au poumon des Vertébrés aériens ;
- les pièces buccales des Insectes, si diverses d'un ordre à l'autre, sont des organes homologues ;
- les homologies se rencontrent entre les éléments reproducteurs des végétaux vasculaires (Ptéridophytes, Gymnospermes, Angiospermes).
Lorsque deux traits apparaissent indépendamment dans deux taxons sans être l'héritage d'un ancêtre commun comme les ailes des insectes et celles des oiseaux, on parle d'analogie ou d'homoplasie.
Cas spécifique chez un même organisme
Lorsqu'un même caractère est présent plusieurs fois chez un même organisme on parle d'homologie sérielle. Par exemple les membres antérieurs et postérieurs d'un tétrapode sont homologues car ils ont la même structure mais ne sont pas issus d'un même organe ancestral commun car présents simultanément sur un même organisme. Le cas de la métamérie aussi en est un exemple. Ce cas de figure est très semblable à une « duplication » de caractères chez un même organisme et donc, comme dans le cas des gènes, cela peut être considéré comme un cas de paralogie. Par exemple, pour les poils de mammifères, des vibrisses de rat et de chat seront plus proches entre elles qu'une vibrisse de chat comparée à un poil du pelage du même chat.
Niveau chromosomique
En génétique des êtres diploïdes, sont homologues chacun des deux chromosomes d'une paire, l'un étant hérité du gamète mâle (« père ») et l'autre du gamète femelle (« mère »). En effet, dans l'œuf fécondé, l'état diploïde résulte de la réunion d'un lot de chromosomes d'origine paternelle et d'un lot de chromosomes d'origine maternelle, qui associés par paires sont homologues deux à deux.
Concernant les relations génétiques au sein d’un organisme unique, la drosophile en est un exemple. Il existe une homologie génétique dans l’hétérochromatine entre tous les chromosomes de Drosophila melanogaster. Plus spécifiquement, cette homologie est située entre le chromosome I et le chromosome IV.
Niveau moléculaire
Des protéines homologues sont des protéines dont les gènes qui les codent ont une origine commune. On reconnaît deux protéines homologues car elles ont des structures spatiales proches et des séquences en acides aminés qui présentent des similarités. Les fonctions de ces protéines peuvent être plus ou moins semblables. On peut quantifier le degré d'homologie entre protéines en réalisant un alignement de séquence optimal, par exemple au moyen de l'algorithme de Smith-Waterman et de matrices de similarité permettant de quantifier la ressemblance entre acides aminés.
On peut trouver des protéines homologues dans des espèces différentes, c'est le cas des globines des vertébrés. Cela traduit l'origine commune de plusieurs espèces et donc l'évolution. On peut trouver des protéines homologues dans une même espèce, c'est le cas des globines alpha et bêta qui composent l'hémoglobine humaine. Cela démontre que l'évolution peut être due à une complexification des génomes.
En biologie moléculaire, c’est principalement les gènes et protéines qui sont considérés homologues. Par exemple, les histones, l’actine, la tubuline, et les protéines avec des fonctions uniques dans les cellules eucaryotes, sont homologues chez les animaux, plantes et fungi. C’est une relation génétique résultant d’une modification héréditaire d’une structure qui est survenue seulement une fois. Elle peut être entre organismes ou au sein d’un organisme unique. Cette définition diffère de celle de Van Valen car elle spécifie les structures comme étant génétiques, et la continuité d’information comme étant l’hérédité. L’homologie moléculaire peut varier sur plusieurs niveaux d’organisation, tous interdépendants. Ainsi, la reconnaissance d’homologie au niveau de l’ADN implique une homologie au niveau des séquences d’acides aminés, et vice versa. Les différents niveaux d’organisation comprennent également la position des nucléotides individuels des séquences d'ADN ainsi que l’interaction des séquences pendant l’expression du gène. L'expression d’un système génétique est considéré homologue lorsqu’elle a été héritée d’un organisme portant le système génétique commun ancestral. Pour reconnaître des homologies de l’ADN, seule l’information de la séquence est utilisée. Cela est fait en comparant les brins parallèles de deux éléments géniques. Ainsi, la reconnaissance d’homologie moléculaire dépend de la recherche de similarités pertinentes à travers les séquences.
Prenons l'exemple de l’étude d’homologie entre la souris et la drosophile. Une homologie a été retrouvée entre l’oncogène int-1 des glandes mammaires des souris et du gène wingless des drosophiles. Le gène int-1 est le gène d’un virus que l'on retrouve dans les tumeurs mammaires d’une souris. Le gène wingless quant à lui intervient dans la morphogenèse et la polarité segmentaire chez la drosophile. Lors de l’étude, l’homologue du gène int-1 a été retrouvé dans le génome de la drosophile, situé à la même position cytologique que le gène wingless. Ainsi, l’homologue du gène int-1 et le gène wingless ont les mêmes séquences donc ils codent des protéines homologues. Lorsque la séquence d’int-1 est exprimée dans la drosophile, celui-ci exprime le phénotype du gène wingless.
Une autre étude s'est interessée aux souches E.coli et Salmonella typhimurium résistantes à apramycine et gentamicine prises sur des animaux et des patients d’hôpitaux. Afin de comparer les plasmides de ces bactéries venant des animaux et des humains, on a étudié la relation moléculaire entre apramycine et gentamicine. Plusieurs souches des animaux et des humains ont été identifiés comme E.coli et Salmonella typhimurium, et étaient tous résistant à gentamicine et apramycine. Ainsi, une homologie génétique a été découverte c’est-à-dire, les plasmides d’animaux et d’humains codent la même protéine, aminoglycosidase 3-N-acetyltransferase.
Plusieurs homologue du gène T-box Brachyury (en) ont récemment été isolés chez divers organismes, des cnidaires jusqu’aux vertébrés et insectes. Il a un rôle dans le processus de différenciation du mésoderme postérieur et la formation de la notochorde, puis de l’élongation de l’axis postérieur. L’homologue de Brachyury a été notamment analysé chez un membre des lophotrochozoaires, Platynereis. Le gène est exprimé le long de l’ancien blastopore en forme de fente et continue d’être exprimé dans l’intestin antérieur et postérieur larvaire après la gastrulation. Cette expression est similaire à celui de la larve basal des deutérostomiens, comme l’étoile de mer Asterina, indiquant qu’il a une homologie entre les deutérostomiens et protostomiens par rapport au développement de l’intestin antérieur larvaire.
Un autre exemple concernant les deutérostomiens et protostomiens montre une expression homologue des gènes codant les facteurs de transcriptions et les molécules de signalisation. Cette homologie permet de conclure que certaines parties du corps des insectes et des vertébrés sont homologues. En particulier, les gènes Wnt/TCF, Dpp or TGFβ/MADS, TollR/Rel Factor, Hedgehog/Ci, Nuclear Receptor, Jack/Stat sont tous similaires chez les deutérostomiens et les ecdysozoaires. En somme, les voies de transduction de signalisation comme les récepteurs de surface membranaire et les facteurs de transcription sont tous orthologues à travers les bilatéraux.
De plus, les taxons cnidaires et bilatéraux démontrent une symétrie bilatérale causée par les expressions Hox homologues le long de l’axe primaire du corps et les expressions dpp homologues asymétriques le long de l’axe secondaire du corps. Le gène Hox détermine le patron de l’axe antério-postérieur, et le gène dpp est impliqué dans modelage dorsale-ventrale chez les bilatériens
Il peut y avoir homologie segmentaire au sein du groupe des arthropodes, c’est le cas des homologues de chacun des huit gènes de la drosophile[3].
Orthologue
Deux séquences homologues de deux espèces différentes sont orthologues si elles descendent d'une séquence unique présente dans le dernier ancêtre commun aux deux espèces. Le terme orthologue a été proposé par Walter Fitch en 1970.
Par exemple, le gène caudal de la drosophile et son homologue, le gène Cdx chez le coléoptère Tribolium castaneum, le criquet Shistocerca gregaria et le crustacé Artemiafranciscana. Le rôle principal du gène caudal est le développement de la région postérieur du corps. Lorsque le gène a été enlevé par un (ARN interférent) chez les coléoptères et les crustacés, ils ont été dépourvus des segments thoraciques et abdominaux, et des segments thoraciques postérieurs, génitaux et post-génitaux respectivement. Cela confirme l’implication du gène caudal dans la caractérisation de la région postérieure.
Paralogue
Deux séquences homologues au sein d'une espèce (ou d'espèces différentes[4]) sont paralogues si elles résultent d'une duplication génique.
Inparalogue
Deux séquences paralogues au sein d'une espèce sont inparalogues si l'événement de duplication a eu lieu après la spéciation[5].
Outparalogue
Deux séquences paralogues au sein d'une espèce sont outparalogues si l'événement de duplication a eu lieu avant la spéciation.
Ohnologue
Deux séquences paralogues sont ohnologues si elles résultent d'un événement de duplication complète du génome. Le terme ohnologue a été proposé par Ken Wolfe en hommage à Susumu Ohno[6].
Xénologue
Deux séquences homologues sont xénologues lorsqu'elles résultent d'un transfert horizontal de gène entre deux organismes.
Notes et références
- (en) Placental mammal diversification and the Cretaceous-Tertiary boundary. Springer MS, Murphy WJ, Eizirik E, O'Brien SJ. Proc Natl Acad Sci U S A. 2003 Feb 4;100(3):1056-61.
- Cyril Langlois, "Faits et arguments paléontologiques en faveur de l’évolution".
- Cornec, J. P., & Gilles, A., « Urbilateria, un être évolué? », M/S: médecine sciences, no 22(5), , p. 493-501
- Fitch WM. Homology: a personal view on some of the problems. Trends Genet. 2000 May;16(5):227-31
- Sonnhammer EL, Koonin EV. Orthology, paralogy and proposed classification for paralog subtypes. Trends Genet. 2002 Dec;18(12):619-20
- Ken Wolfe (2000) Robustness—it's not where you think it is. Nature Genetics May;25(1):3-4.
Voir aussi
Bibliographie
- John V. Freudenstein, Characters, States, and homology, Systematic biology, 2005.
- Jacob, F. (1990). L'unité du vivant.
- Hall, B. K. (2003). Evo-Devo: evolutionary developmental mechanisms.International Journal of Developmental Biology, 47(7/8), 491-496.
- Davis, G. K., & Patel, N. H. (1999). The origin and evolution of segmentation.Trends in Genetics, 15(12), M68-M72.
- Technau, U. (2001). Brachyury, the blastopore and the evolution of the mesoderm. Bioessays, 23(9), 788-794.
- Bledsoe, A. H., & Sheldon, F. H. (1990). Molecular homology and DNA hybridization. Journal of molecular evolution, 30(5), 425-433.
- Erwin, D. H., & Davidson, E. H. (2002). The last common bilaterian ancestor.Development, 129(13), 3021-3032.
- Chaslus-Dancla, E., Pohl, P., Meurisse, M., Marin, M., & Lafont, J. P. (1991). High genetic homology between plasmids of human and animal origins conferring resistance to the aminoglycosides gentamicin and apramycin.Antimicrobial agents and chemotherapy, 35(3), 590-593.
- Falkow, S., Rownd, R., & Baron, L. S. (1962). Genetic homology between Escherichia coli K-12 and Salmonella. Journal of bacteriology, 84(6), 1303-1312.
- Newman, S. A. (2006). The developmental-genetic toolkit and the molecular homology-analogy paradox. Biological Theory, 1(1), 12.
- Finnerty, J. R., Pang, K., Burton, P., Paulson, D., & Martindale, M. Q. (2004). Origins of bilateral symmetry: Hox and dpp expression in a sea anemone.Science, 304(5675), 1335-1337.
- Sandler, L., & Novitski, E. (1956). Evidence for genetic homology between chromosomes I and IV in Drosophila melanogaster, with a proposed explanation for the crowding effect in triploids. Genetics, 41(2), 189.