Homoplasie
L’homoplasie est la similitude d’un état de caractère chez différents taxons qui, contrairement à l’homologie, ne provient pas d’un ancêtre commun.
Il existe différents types d’homoplasie : la convergence, le parallélisme et la réversion.
- la convergence : ressemblance apparue indépendamment chez des taxons distants phylogénétiquement.
- le parallélisme : ressemblance apparue chez des taxons relativement proches.
- la réversion : un état dérivé d’un caractère revient à un état ancestral (antérieur).
L'homoplasie de fonction est une analogie, c'est-à-dire une ressemblance de caractères remplissant les mêmes fonctions biologiques.
Une homoplasie peut être simple, telle qu’un changement de monomères d’ADN, ou bien complexe, comme une réorganisation indépendante de systèmes multiples composés de plusieurs gènes et membres qui convergent vers une même fonction. Dans tous les cas, ces similitudes sont survenues par des événements indépendants sur des lignages séparés, puisque ces caractéristiques ne correspondent pas à des évènements uniques chez un ancêtre commun.
Niveau moléculaire
Il n’y a pas de preuve à ce jour que les données moléculaires soient moins homoplasiques que les données morphologiques. La convergence est peut-être moins fréquente parmi certains caractères moléculaires tels que l’ADN ou les sites de restriction. Cependant, ils sont globalement moins exposés à la sélection naturelle que les caractères morphologiques.
Remplacement d'un nucléotide
Les organismes qui occupent un rôle écologique similaire évoluent souvent de manière convergente avec des phénotypes similaires qui reflètent des pressions de sélection similaires. Cependant, l’homoplasie peut être engendrée en l'absence de sélection via des mutations récurrentes[1]. Lorsqu’un organisme n’est pas sous sélection, les mutations, ne diminuant pas l'adaptation, ont tendance à s’accumuler. Ainsi, des mutations analogues peuvent s’accumuler par chance dans les espèces. Les mêmes remplacements de nucléotides (mutations analogues) peuvent se produire dans différentes lignées de manière indépendante par hasard et s’accumuler. Ce phénomène ne reflète donc pas un ancêtre commun mais plutôt un changement parallèle, inverse ou convergent[2].
Prenons l’exemple de deux espèces différentes, 1 et 2. Une mutation peut surgir sur le lignage conduisant à l’espèce 1, changeant ainsi le T en troisième position pour un A. Indépendamment, une mutation peut aussi modifier le nucléotide G en troisième position pour un A dans le lignage qui conduit à l’espèce 2. Finalement, les séquences des deux espèces sont identiques bien que le nucleotide A ne soit pas hérité d’un ancêtre commun. Il ne peut être alors considéré comme homologue. Il s’agit ainsi d’une homoplasie, c'est-à-dire une similitude qui résulte d’évènements indépendants sur deux lignages différents. Le nombre d’homoplasies récurrentes, comme celle de l’exemple précédent, devrait généralement augmenter avec le nombre de taxons dans une lignée[1]. En fait, plus on compare d’espèces divergentes, plus on a de chances que deux ou plusieurs de celles-ci partagent une mutation basée sur le hasard et non sur un ancêtre commun.
Transfert de gènes horizontal
Le transfert de gènes horizontal peut aussi générer de l’homoplasie. Il s’agit d’une manière de propagation d’un trait dans un taxon qui diffère de la forme de transmission habituelle soit verticale et biparentale. C’est ainsi, par exemple, que de nombreux microbes infectieux ont acquis la résistance aux mêmes antibiotiques par la propagation et l’incorporation de plasmides contenant la séquence génétique pour une telle résistance[1]. Un autre exemple d’homoplasie grâce au transfert horizontal de gènes sont les syncytines. Ces protéines jouent un rôle important dans le développement du placenta. Elles provoquent la fusion des cellules et permettent le développement du syncytiotrophoblaste, un des tissus qui composent le placenta. Celui-ci possède des propriétés immunodépressives, ce qui évite au système immunitaire de la mère de rejeter le fœtus. Les gènes qui codent les syncytines proviennent de rétrovirus ayant infecté de nombreuses fois les mammifères de manière indépendante. En effet, on retrouve ces protéines dans plusieurs mammifères tels que les souris, les écureuils et les humains. Chez l’être humain, il existe deux de ces gènes intacts d’enveloppe issus d’infections par des rétrovirus endogènes exprimés de manière spécifique dans le placenta : Syncytin-1 et Syncytin-2. Pour faire fusionner les cellules, les gènes de ces protéines utilisent le même mode d’action que les rétrovirus. En fait, tout comme les protéines ancestrales de rétrovirus, les syncytines reconnaissent un récepteur cellulaire spécifique et ont une activité immunosuppressive. Ainsi, il y a de nombreuses années, un virus aurait infecté la cellule, copié son ADN pour ensuite l’intégrer au génome de la cellule. La cellule aurait exprimé ses nouveaux gènes et commencé à synthétiser de la syncytine. Ce processus se serait produit dans plusieurs groupes différents de mammifères placentaires de manière indépendante grâce au phénomène de convergence évolutive.
Triade catalytique
On désigne triade catalytique les trois résidus d’acides aminés qui interviennent ensemble dans le site actif de certaines hydrolases et transférases. Les protéases de sérine jouent un rôle essentiel dans de nombreuses fonctions cellulaires et extracellulaires telles que le processus de coagulation sanguine, la digestion des protéines, la signalisation cellulaire, l’inflammation et la transformation des protéines[3]. Ceux-ci contiennent tous la triade Ser/His/Asp. Deux des protéases-sérine les plus connues qui utilisent cette triade sont chrymotrypsine et la trypsine. La subtilisine utilise elle aussi la triade catalytique mais elle n’a aucune similarité de séquence avec les deux protéases précédentes. Elle adopte la structure de feuille ouverte α/β- torsadée plutôt que la structure en double tonneau-β[4]. La subtilisine et la chymotrypsine sont de bons exemples d’homoplasie. Bien que les plis protéiques de ces protéases soient complètement différents, les deux ont convergé vers un mécanisme similaire de Ser/His/Asp. De plus, la trypsine et la chymotrypsine sont des protéases digestives tandis que la subtilisine est une protéase bactérienne produite par Bacillus subtilis. Les contraintes chimiques et physiques sur la catalyse des enzymes ont ainsi engendré des arrangements identiques de triade. En fait, la nature même des réactions chimiques catalysées ont conduit de façon indépendante aux mêmes types de sites actifs d’au moins 23 superfamilles distinctes.
Voie de signalisation des EGFR
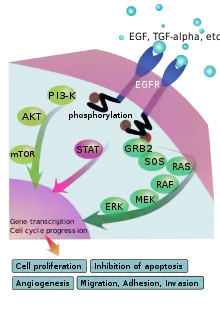
La voie de signalisation des EGFR (Epidermal Growth Factor Receptor) joue un rôle essentiel dans le développement des vertébrés et des invertébrés. Cette voie s’est développée chez les deux groupes de manière indépendante. Chez les vertébrés, elle détient un rôle important dans l’embryogenèse et le développement. EGFR présente un rôle physiologique préférentiel dans le développement des tissus d’origine épithéliale. L’activation du récepteur a pour conséquence d’augmenter la prolifération cellulaire et la motilité cellulaire ainsi que de diminuer l’apoptose. Tout comme chez les vertébrés, la voie des EGFR est nécessaire à un grand nombre d’étapes dans le développement des invertébrés, plus particulièrement de la Drosophile. Cette voie permet d’établir l’axe antéro-postérieur de l’embryon, de contrôler le destin cellulaire pendant le développement des disques embryonnaires et imaginaux et pendant l’oogenèse.
Écholocation
L’homoplasie au niveau moléculaire engendre parfois de l’homoplasie au niveau morphologique. En effet, la comparaison des génomes d’écholocation des chauves-souris et des dauphins identifie plusieurs substitutions d’acides aminés convergentes dans les gènes impliqués dans l’audition et la vision. Il a été déterminé que les deux types d’écholocation soit chez la chauve-souris et le dauphin avaient les mêmes mutations dans une protéine particulière appelée prestine qui affecte la sensibilité de l’ouïe[5]. De plus, l’analyse d’autres gènes connus pour être impliqués dans l’audition a démontré que plusieurs protéines avaient aussi été modifiées de manière similaire chez ces deux mammifères. En fait, pas moins de 200 gènes ont changé indépendamment de la même façon. La plupart de ces gènes sont impliqués dans l’audition mais d’autres ne sont pas directement liés à l’écholocation jusqu’à présent mais plusieurs seraient importants pour la vision. Ainsi, la convergence n’est pas qu’un processus restreint à plusieurs loci, mais peut engendrer des homoplasies moléculaires répandues dans le génome. L’écholocation est un mode de repérage et de visualisation utilisé par quelques mammifères tels que les chauves-souris et les dauphins lors de leurs déplacements pour localiser leurs proies et les obstacles dans leur environnement. Ceux-ci émettent des ondes sonores qui sont renvoyées lorsqu’elles se heurtent à une proie ou un obstacle. Elles sont ensuite enregistrées et visualisées par le cerveau de l’animal ce qui lui fournit ainsi une image en trois dimensions de son environnement. Puisque cette caractéristique leur procure un net avantage, il n’est donc pas difficile de comprendre pourquoi la sélection naturelle conserva ces mutations chez ces mammifères très différents.
Niveau morphologique
De nombreuses similarités phénotypiques ont évolué de manière indépendante dans différentes lignées. Ces homoplasies sont convergentes soit au niveau de la structure, soit au niveau de la fonction ou même les deux. Ces convergences morphologiques sont des réponses évolutionnaires à des patrons de sélection naturelle similaires dans les habitats des espèces. Il existe de nombreux exemples de ces homoplasies physiologiques dont quelques-uns sont décrits ci-dessous.
Forme hydrodynamique du corps
La forme du corps des vertébrés marins comme les poissons, les ichtyosaures, les dauphins et les phoques est un cas intéressant d’homoplasie. Prenons comme exemple les requins et les dauphins qui possèdent la même forme hydrodynamique. Puisque les dauphins ont évolué depuis un ancêtre tétrapode, ce caractère ne peut être dû par la présence d’un ancêtre commun. En effet, les fossiles conservés prouvent bel et bien que certains mammifères terrestres ont modifié leur mode de vie passant de la terre à l’eau et ont évolué des formes hydrodynamiques du corps tout comme les espèces marines déjà présentes à cette époque[6]. Ainsi, bien que ces deux vertébrés partagent des ancêtres communs, leurs formes de corps n’est pas dérivé de ceux-ci. Ces deux espèces ont été confrontées aux mêmes pressions de sélection des milieux aquatiques. Leurs formes de corps hydrodynamiques est ainsi une adaptation indépendante et convergente à la vie aquatique afin d’améliorer leurs performances et leurs vitesses sous l’eau.
Aile et vol actif

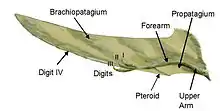
L’exemple d’homoplasie le plus souvent énoncé est sans contredit le vol actif. Il s’agit d’une convergence morphologique de structure et de fonction. Trois groupes séparés de tétrapodes ont modifié leurs membres antérieurs en ailes soit les ptérosaures, les oiseaux et les chauves-souris. Bien que leurs ailes soient superficiellement semblables, elles sont construites très différemment. Les ailes de chauves-souris sont composées en grande partie de leur main dont les doigts sont très allongés et entre lesquels est tendu une membrane de peau palmée[7]. Les ailes des ptérosaures, aujourd’hui éteints, sont similaires. Cependant, elles sont constituées d’un seul doigt, au lieu de 4, d’où s’étire une membrane de peau jusqu’au côté de l’animal. Les ailes des oiseaux sont encore plus différentes puisqu’elles sont constituées de plumes allongées attachées aux os s’étendant sur tout le long de leurs membres antérieurs. Ces différences structurales suggèrent que leurs ailes, bien que d’apparences similaires, ont évolué de manière indépendante et ne seraient pas héritées d’un ancêtre commun avec des ailes. Les trois lignées distinctes auraient, en fait, répondu de la même façon aux mêmes pressions de sélection. Finalement, les ailes de ces trois groupes sont des modifications convergentes des membres antérieurs, structure originellement utilisée pour la marche et la course. Sans peau membraneuse ou plumes sur leurs membres antérieurs, ces espèces seraient confinées à une vie au sol et ne pourrait éviter la forte prédation qui s’y trouve.
En plus des oiseaux, des chauves-souris et des ptérodactyles, le vol actif a aussi évolué indépendamment chez les insectes. Leurs ailes sont, quant à elles, radicalement différentes. Ceux-ci n’ont pas modifié leurs membres antérieurs comme les tétrapodes. En fait, l’origine des ailes des insectes seraient une modification des branchies originellement présentes seulement lors du stade larvaire[7]. Présentes sur de nombreux segments sur l’abdomen des larves aquatiques, ces structures branchiales se sont développées sur certains segments alors qu’ils ont été supprimés sur d’autres. Cette restructuration a alors mené aux premiers insectes volants à 4 ailes. Il est possible d’observer cette condition primitive chez certains insectes aquatiques adultes comme l’éphémère. Encore une fois, ce sont pour des raisons fonctionnelles et non héréditaires que les insectes auraient développé la capacité de vol. La sélection naturelle aurait alors avantagé les insectes dotés d’ailes passant du milieu aquatique à l’atmosphère.
Yeux camérulaires
L’évolution répétée des yeux de type « chambre noire » ou camérulaire est l’un des exemples d’homoplasie des plus impressionnants. On appelle yeux camérulaires les yeux dont la rétine est concave et qu’une image de l’espace visuel peut être réalisée par l’adjonction d’une lentille convergente devant cette surface[8]. Un compartiment liquidien ou humeur vitrée sépare le cristallin de la rétine, ce qui a pour conséquence la formation de l’image d’un objet dans le plan focal rétinien[9]. Ce type d’œil a évolué indépendamment chez les cnidaires (certaines méduses), les céphalopodes (comme les calmars et les pieuvres) et les vertébrés (oiseaux, mammifères). Il existe cependant quelques différences morphologiques entre les yeux des céphalopodes et des vertébrés. Ces derniers possèdent une rétine en position inversé comparativement à la rétine directe des céphalopodes. Par exemple, les photopigments sont directement touchés par les rayons lumineux chez la seiche par rapport aux vertébrés où la lumière doit traverser un grand nombre de couches cellulaires avant d’atteindre les cellules photoréceptrices[9].
Mammifères placentaires et mammifères marsupiaux

_(6008352459).jpg.webp)
Certains animaux très éloignés géographiquement mais vivants dans des habitats similaires et se nourrissant d’aliments similaires ont des ressemblances physiologiques étonnantes. C’est le cas des mammifères placentaires d’Amérique du Nord et des mammifères marsupiaux d’Australie regroupant plusieurs espèces partageant de nombreuses homoplasies dues à l’évolution convergente. Les Marsupiaux[10] se caractérisent par un développement embryonnaire qui commence dans l’utérus maternel et se poursuit dans une poche ventrale. Ils se distinguent des Placentaires[11], dont le développement embryonnaire se déroule entièrement dans l’utérus[12]. Certains Marsupiaux d’Australie ressemblent étonnamment à plusieurs Placentaires de divers continents.
Prenons comme exemple les écureuils volants (Glaucomys volans) et les phalangers du sucre (Petaurus breviceps). Ce dernier est un marsupial arboricole tandis que l’écureuil est un placentaire qui saute d’arbre en arbre dans les forêts d’Amérique du Nord. Les deux présentent des adaptations similaires puisqu’ils sont dotés les uns et les autres de membranes qui leur permettent de planer entre les arbres et ralentir leur chute. On ne peut expliquer ces ressemblances par la présence d’un ancêtre commun puisque ces deux lignées distinctes de mammifères ont divergé après que la Pangée se soit déjà séparée en continents du nord et du sud[13]. La parenté entre ses espèces est très lointaine et leur ancêtre commun ne détenait pas la capacité de planer. On parle alors ici d’homoplasie et non d’homologie. Ainsi, bien qu’ils aient évolué indépendamment à partir d’ancêtres différents, ces deux petits mammifères se sont adaptés de manière semblable à des environnements similaires.
Végétaux en milieu désertique
L’homoplasie peut être retrouvé aussi bien chez les animaux et les insectes que chez les végétaux. Comparons, par exemple, les cactus retrouvés en Amérique du Nord et du Sud tels que Echinocereus avec les euphorbes des déserts du sud de l’Afrique et de Madagascar. Leurs apparences générales sont extraordinairement similaires. En effet, les euphorbes ont évolué au fil du temps des caractéristiques physiques semblables aux cactus de sorte qu’elles sont très souvent appelées « cactus » à tort. Ces deux plantes occupent des habitats secs et désertiques ainsi ils ont développé des caractéristiques similaires pour minimiser leurs pertes en eau. Ils ont développé des tiges épaisses capables de stocker ainsi que des structures épineuses pour décourager les animaux à se nourrir d’eux en plus de réduire les surfaces d’évaporation[13]. Ces végétaux pratiquent aussi la photosynthèse CAM, métabolisme acide crassulacéen, et leurs stomates ne s’ouvrent que pendant la nuit pour absorber le dioxyde de carbone. Ainsi, bien que le cactus d’Amérique et l’euphorbe d’Afrique ne partagent pas la présence d’un ancêtre commun avec ces caractéristiques dans leur lignée, les deux ont convergé vers les mêmes solutions afin de lutter contre les périodes de sécheresse.
Ces exemple d’homoplasie sont tous formés par l’environnement physique des organismes en question soit la possibilité de voler dans les airs pour les ailes ou de se déplacer efficacement dans l’eau pour la forme du corps hydrodynamique, la perception de la lumière pour les yeux camérulaires, etc. On pourrait s’attendre à ce que la présence fixe de ces caractéristiques environnementales conduise à l’adaptation de nombreuses espèces.
Notes et références
- (en) Givnish, T., & Sytsma, K., Molecular Evolution and Adaptative Radiation, Cambridge University Press,
- (en) Rogozin, I., Thomson, K., Csuros, M., Carmel, L., & Koonin, E., « Homoplasy in genome-wide analysis of rare amino acid replacements : the molecular-evolutionary basis for Vavilov's law of homologous series », Biology Direct, 2008, mar 17, p. 3 :7
- (en) Buller, A & Townsend, C., « Intrinsic evolutionary constraints on protease structure, enzyme acylation, and the identity of the catalytic triad », Proceedings of the National Academy of Sciences of the United States of America, 2013, feb 4, p. 110(8) : 653-661
- (en) Ekici, O., Paetzel, M., & Dalbey, R., « Unconventional serine proteases : Variations on the catalytic Ser/His/Asp triad configuration », Protein Science, , p. 17 (12) : 2023-2037
- (en) Joe Parker, Georgia Tsagkogeorga, James A. Cotton, Yuan Liu, Paolo Provero, Elia Stupka & Stephen J. Rossiter, « Genome-wide signatures of convergent evolution in echolocating mammals », Nature, 2013, oct 10, p. 502(7470) : 228-231
- (en) Kampourakis, K, Understanding Evolution, New York, Cambridge University Press,
- (en) McGhee, G., Convergent Evolution : Limited Forms Most Beautiful, Cambridge, MIT Press,
- Yves Le Men, La vision dans le monde animal, AlterPublishing, (présentation en ligne)
- Tanzarella, S, Perception et communication chez les animaux, Bruxelles, De Boeck & Larcier,
- Marsupiaux
- Placentaires
- Campbell, N., Reece, J., Urry, L., Cain, M., Wasserman, S., Minorsky, P. & Jackson, R., Biologie de Campbell (4e édition), ERPI,
- (en) Russell, P., Hertz, P., & McMillan, B., Biology : The Dynamic Science (4th edition), Boston, Kindle Edition,
Voir aussi
Articles connexes
Liens externes
- http://extraordinairemondeduplacenta.weebly.com/la-syncytine.html
- http://docnum.univ-lorraine.fr/public/SCD_T_2011_0050_CHRETIEN.pdf
- http://www.universalis.fr/encyclopedie/echolocation/
- http://www.futura-sciences.com/sante/dossiers/medecine-oeil-vision-dela-vision-667/page/2/
- http://www.australia-australie.com/articles/les-marsupiaux
- http://www.universalis.fr/encyclopedie/mammiferes-placentaires-origine-des/