Métabolisme acide crassulacéen
Le métabolisme acide crassulacéen (CAM, pour crassulacean acid metabolism) est un type de photosynthèse qui permet à certaines plantes terrestres chlorophylliennes de fixer le carbone. Ce métabolisme d'assimilation chlorophyllienne crassuléenne concerne les plantes grasses[1] présentant des caractères de crassulescence (forme biologique végétale ayant une concentration élevée du suc vacuolaire, conférant à la plante son aspect charnu) comme les cactées ou les euphorbes, ainsi que de nombreux xérophytes et épiphytes vivant dans des environnements qui peuvent être périodiquement très pauvres en eau, comme les déserts[2] - [3]. Le nom du métabolisme vient de la famille de la plante où il a été observé pour la première fois, soit chez les Kalanchoe de la famille des Crassulacées.
Ce métabolisme est à l'origine du goût parfois légèrement acidulé des plantes grasses dû à l'accumulation d'acide citrique, isocitrique et malique synthétisés la nuit et à la désacidification diurne progressive[4]
Anatomie des plantes CAM
Comme le métabolisme CAM implique de faire d'importantes réserves d'acide malique à l'intérieur des cellules végétales, les plantes ayant ce métabolisme possèdent de grandes cellules, à rapport surface/volume faible. Ces grandes cellules peuvent alors contenir de grandes vacuoles[3] remplies d'acides organiques (acide citrique, isoctrique et malique) et aussi, elles peuvent retenir de grandes quantités d'eau. Les plantes CAM ont également des feuilles et des tiges modifiées, qui ne possèdent pas beaucoup de stomates et qui ont une cuticule épaisse afin de réduire au maximum les échanges gazeux de vapeur d'eau avec l'extérieur. C'est surtout dans leurs feuilles charnues que les plantes CAM font leurs réserves d'acide malique, car ce sont les cellules des feuilles qui possèdent de grosses vacuoles[5] - [2].
Mécanismes
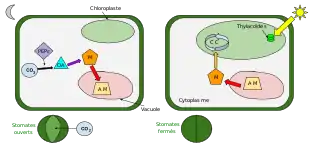
À la différence des plantes C4, qui effectuent une séparation spatiale entre la fixation initiale du CO2 et son utilisation par le cycle de Calvin, les plantes CAM réalisent une séparation temporelle des deux voies, puisque l'une a lieu la nuit et l'autre le jour[3] - [5] - [6]. En revanche, le métabolisme photosynthétique CAM fait intervenir un système à double carboxylation tout comme les plantes C4, puisque le CO2 est fixé à deux reprises dans la cellule végétale, une première fois avec la phosphoénolpyruvate carboxylase dans le cytosol, puis une seconde fois avec la rubisco dans le chloroplaste[2].
De nuit
Les stomates des plantes CAM s'ouvrent uniquement la nuit, car à ce moment, la température est plus basse que durant la journée et le taux d'humidité est plus élevé[3] - [5] - [2]. Cela provoque une faible évapotranspiration et donc de faibles pertes d'eau. C'est pourquoi les plantes CAM se sont spécialisées dans la fixation du CO2 pendant la nuit[2].
Cette fixation est réalisée par la PEP carboxylase sur le phosphoénolpyruvate (PEP), qui provient de la dégradation de l'amidon et du saccharose produit dans le chloroplaste le jour[5]. Cette fixation permet de former de l'oxaloacétate, qui sera immédiatement réduit en malate, puis stocké dans une vacuole sous forme d'acide malique, d'où le nom de plante à métabolisme acide[2]. Autrement dit, durant la nuit, la plante CAM fait "le plein" de CO2 sous forme d'acide malique, mais elle ne peut pas le transformer tout de suite en sucre du fait de l'obscurité. En effet, comme toutes les autres plantes, les plantes CAM ont besoin de l'énergie lumineuse du jour pour compléter le cycle de Calvin et pour ainsi accomplir la photosynthèse en entier.
De jour
Les plantes CAM vivant dans des milieux arides où l'air est très sec, les stomates de leurs feuilles restent fermés durant la journée pour limiter les pertes d'eau par évapotranspiration[2] - [5].
Les plantes CAM fixent donc le CO2 la nuit et le stockent sous forme d'acide malique dans leurs vacuoles pour l'utiliser ensuite le jour lorsque les stomates sont fermés[2]. À la fin de la nuit, l'acide malique est retransformé en malate, qui est transféré hors des vacuoles pour être décarboxylé en CO2 et en pyruvate dans le cytosol. Cette décarboxylation est catalysée par une enzyme malique dépendante NAD+ (ou NADP+) ou une PEP carboxykinase[5]. Après cette étape, le CO2 est fixé à nouveau, mais cette fois grâce à la rubisco, pour ensuite entrer dans le cycle de Calvin[2] dans le chloroplaste[3]. C'est à la suite de ce cycle que des sucres comme l'amidon et le saccharose sont synthétisés[2]. Le pyruvate sert aussi à refaire le stock d'amidon et est converti en sucres par néoglucogénèse.
Le cycle de Calvin a lieu le jour, car il a besoin du NADPH et de l'ATP produits durant la phase claire de la photosynthèse nécessitant de l'énergie lumineuse. Contrairement aux types de plantes C3 et C4, chez les plantes CAM cette étape se fait en absence d'échanges gazeux avec l'atmosphère, car à ce moment, leurs stomates sont fermés[2].
La seule source de CO2 disponible pour effectuer le cycle de Calvin le jour est la réserve d'acide malique produite durant la nuit. La quantité de CO2 disponible diminue rapidement en cours de journée, jusqu'à ce que la vacuole de stockage soit épuisée[2]. La photosynthèse est moins efficace chez les plantes CAM, le CO2 étant le facteur limitant.
Intérêts physiologiques
L'un des avantages des plantes CAM est que leur métabolisme est principalement contrôlé par des facteurs du milieu, plutôt que par une base génétique comme les plantes C4[2] - [5]. En effet, le métabolisme crassulacéen peut être constitutif ou inductible. Pour une espèce inductible comme Mesembryanthemum cristallinum, le métabolisme adopté dépend donc de la quantité d'eau disponible pour la plante. Par exemple, au printemps, lorsque le sol est gorgé d'eau, les plantes CAM peuvent manifester un métabolisme de type C3 le jour[3] - [2]. Par contre, l'été, les sols sont plus secs et le manque d'eau, de même que l'élévation de la teneur en sel des sols provoquent chez les plantes CAM de rapides changements dans l'expression de leurs gènes et donc de leurs enzymes, ce qui leur permet de mettre en place le métabolisme crassulacéen. On peut donc dire que le mécanisme de fixation du carbone des plantes CAM est souple et très adapté aux variations du milieu[2].
Un autre avantage majeur de ce métabolisme est de limiter les pertes d'eau grâce à la fermeture des stomates durant la journée[7], ce qui est hautement important pour les plantes CAM qui vivent dans des conditions de haute intensité lumineuse et de stress hydrique[2] - [5] - [6]. Les pertes d'eau par CO2 fixé sont chez les plantes CAM de 3 à 6 fois moindres que chez les plantes C4 et de 4 à 10 fois moindres que chez les plantes C3. Ainsi, les plantes CAM fixent de 10 à 40 kg de CO2 par kilo d'eau transpiré, alors que les plantes C4 n'en fixent que de 2 à 5 kg et les plantes C3 de 1 à 3 kg[2].
Le métabolisme des plantes CAM permet également d'éviter la photorespiration qui se produit particulièrement chez les plantes C3. En effet, comme le CO2 est fixé en premier lieu par la PEP carboxylase plutôt que par la rubisco, il n'y a pratiquement plus de photorespiration possible du fait de l'absence d'affinité entre la PEP carboxylase et l'oxygène[6]. La PEP carboxylase a une bien meilleure affinité avec le CO2 que la rubisco, elle en fixe donc plus sans gaspiller d'énergie en fixant de l'oxygène inutilement[7]. D'autre part, les plantes CAM contiennent aussi de la rubisco (qui a une grande affinité avec l'oxygène) pour faire le cycle de Calvin, mais rappelons que ce cycle a lieu le jour, lorsque les stomates sont fermés. La rubisco est alors saturée de CO2 provenant de la vacuole d'entreposage et il y a très peu d'oxygène de présent, c'est pourquoi la rubisco ne fait pas de photorespiration et pourquoi elle est si efficace chez les plantes CAM et C4[7]. On dit alors que les plantes CAM ont une efficacité d'utilisation de l'azote élevée, car la rubisco contient beaucoup d'azote[7].
Aussi, comme le CO2 est tout de suite converti en oxaloacétate sitôt qu'il est fixé par la PEP carboxylase, le système n'est jamais saturé en CO2 et il peut en fixer davantage[7]. Ainsi, l'enzyme PEP carboxylase travaille à plein régime en présence de CO2, c'est pourquoi les plantes C4 et CAM sont plus efficaces à fixer le carbone que les C3.
Désavantages physiologiques
Le métabolisme crassulacéen a beaucoup d'avantages, mais comme toute chose, il comporte aussi des coûts :
- Tout comme pour les C4, le métabolisme des plantes CAM est plus coûteux en énergie que celui des plantes C3. Comme il nécessite une double machinerie enzymatique pour une double carboxylation (c'est-à-dire plus de structures, plus d'enzymes, etc.), cela provoque davantage de coûts pour les plantes CAM, qui produisent ces machineries à partir de leurs réserves énergétiques[7].
- D'autre part, les plantes CAM sont les seules à pouvoir entreposer des réserves de CO2 sous forme d'acide malique, ce qui s'avère être un avantage autant qu'un inconvénient, car ces réserves à l'intérieur d'une vacuole prennent beaucoup de place dans la cellule végétale[7]. Cela nécessite des cellules plus grandes et donc d'autres coûts énergétiques.
- Finalement, l'activité photosynthétique globale des plantes CAM est plutôt modeste, car comme il a été mentionné, celles-ci sont limitées par leur réserve de CO2[8] - [9] - [10]. Ainsi, lorsque la vacuole de réserve est vide, il n'y a plus de photosynthèse possible pour les plantes. Cela limite très fortement leur croissance[11], car leurs enzymes et toute leur machinerie photosynthétique ne fonctionnent pas autant qu'elles le peuvent, puisqu'elles n'ont pas accès en permanence à du CO2 , contrairement aux C4 et C3 qui ont leurs stomates ouverts pour faire les échanges gazeux durant la journée. C'est pourquoi les plantes CAM poussent si lentement, comme les cactus par exemple.
Intérêt économique/agronomique
L'intérêt des plantes CAM est plutôt limité au niveau agronomique, sauf en ce qui concerne l'ananas, la vanille ou l'agave. Mentionnons cependant que ces plantes peuvent être cultivées dans des milieux non arides[2].
Distribution[12]
On retrouve généralement les plantes CAM dans des régions extrêmement arides ou riches en sel et très chaudes, où la lumière n'est pas limitante pour la croissance, mais où il fait relativement frais la nuit[5]. Il s'agit là des seules conditions où les plantes CAM peuvent surpasser en efficacité les C3 et les C4. En effet, en dehors de ces conditions, les plantes CAM ont une croissance trop lente et un coût énergétique trop élevé pour être avantagées. Leur distribution est donc relativement limitée à ces endroits.
On retrouve aussi les plantes CAM en tant qu'épiphytes dans la cime des arbres dans certaines régions tropicales, où il y a suffisamment d'humidité atmosphérique pour supporter leur métabolisme[11], malgré un système racinaire limité.
Des plantes CAM submergées ont même été trouvées dans certains lacs où la majorité des plantes présentes utilisent le CO2 dissout le jour pour la photosynthèse, mais où les plantes CAM utilisent le CO2 dissout dans l'eau la nuit. Cela les avantage, car il y a plus de CO2 disponible la nuit que le jour, puisque le jour, il y a davantage de plantes qui entrent en compétition pour son obtention.
Liste de plantes CAM
Ce métabolisme se rencontre généralement chez les plantes vivant sous des conditions arides, par exemple le cactus ou l'ananas. Les plantes CAM représentent environ 6 à 10 % des plantes vasculaires[2] - [13]. Plus spécifiquement, on considère 33 familles d'angiospermes comme étant des plantes CAM[13], parmi lesquelles on retrouve:
- Les Aizoaceae
- Les Cactaceae
- Les Crassulaceae[14]
- Hoya carnosa (Apocynaceae)
- Les sansevières (Liliaceae)
- De nombreuses espèces d'orchidées[15]
- Des Agavaceae et des Bromeliaceae[16], dont l'ananas et la mousse d'Espagne (Tillandsia usneoides)
Notes et références
- Étienne Roth (dir.), Bernard Poty (dir.), Georgette Délibrias et al. (préf. Jean Coulomb), Méthodes de datation par les phénomènes nucléaires naturels, Paris, Éditions Masson, coll. « Collection CEA », , 631 p. (ISBN 2-225-80674-8), chap. 15 (« Le carbone 14 »).
- J.Farineau, J.-F. Morot-Gaudry, La photosynthèse, Processus physiques, moléculaires et physiologiques, Paris, Inra, , 405 p. (ISBN 2-7380-1209-4), p.310-312
- (en) Micheal J Crawley, Plant ecology, Londres, Blackwell Scientific Publications, , 497 p. (ISBN 0-632-01363-X), p. 368
- (en) J. Keeley, G. Busch, « Carbon Assimilation Characteristics of the Aquatic CAM Plant, Isoetes howellii », Plant Physiology, vol. 76, no 2, , p. 525–530 (DOI 10.1104/pp.76.2.525).
- Gerhard Richter, Métabolisme des végétaux, Physiologie et biochimie, Allemagne, Presses polytechniques et universitaires romandes, , 526 p. (ISBN 2-88074-231-5), p. 175-177
- (en) C. Barry Osmond, William W. Adams and Stanley D. Smith, Plant Physiological Ecology : field methods and instrumentation, USA, Chapman and Hall, , 457 p. (ISBN 0-412-40730-2), chapitre 12 p. 255-275
- (en) Jessica Gurevitch, Samuel M. Scheiner, Gordon A. Fox, The Ecology of plants, Massachusetts, USA, Sinauer Associates, inc., , 574 p. (ISBN 978-0-87893-294-8), p.26-30
- (en) Bowyer J.R., Leegood R.C., 2 Photosynthesis, Plant Biochemistry Academic Press, , Chap. 2: 49-110
- (en) Heldt H.W., Plant Biochemistry and Molecular Biology, Oxford University Press, , 522 p.
- (en) Buchanan B.B., Gruissem W., Russell L.J., Biochemistry and Molecular biology of plants, Rockville, Maryland, American Society of Plant Physiologists, , 1367 p.
- (en) O.W. Archibold, Ecology of World Vegetation, Saskatoon, Canada, Chapman and Hall, , 512 p. (ISBN 0-412-44290-6), p.114-115
- (en) Michael J Crawley, Plant ecology, Londres, Blackwell Scientific Publications, , 497 p. (ISBN 0-632-01363-X), p. 371
- (en) Antony N. Dodd, Anne M. Borland, Richard P. Haslam et Howard Griffiths, « Crassulacean acid metabolism: plastic, fantastic », Journal of Experimental Botany, vol. 53, no 369, , p. 569–580 (ISSN 1460-2431 et 0022-0957, DOI 10.1093/jexbot/53.369.569, lire en ligne, consulté le )
- (en) J. Thiede et U. Eggli, « Crassulaceae », dans Flowering Plants · Eudicots, vol. 9, Springer Berlin Heidelberg, (ISBN 978-3-540-32214-6, DOI 10.1007/978-3-540-32219-1_12, lire en ligne), p. 83–118
- http://www.publish.csiro.au/?act=view_file&file_id=FP10084.pdf
- (en) Alejandra Matiz, Paulo Tamaso Mioto, Adriana Yepes Mayorga, Luciano Freschi et Helenice Mercier, Agricultural and Biological Sciences : "Photosynthesis" : Chapitre : CAM Photosynthesis in Bromeliads and Agaves : what can we learn from these plants ?, Zvy Dubinsky, , 382 p. (ISBN 978-953-51-1161-0, lire en ligne).