Rubisco
La Rubisco[alpha 1], de son nom complet ribulose-1,5-bisphosphate carboxylase/oxygénase, est l'enzyme-clé de la photosynthèse. C'est elle qui permet la fixation du dioxyde de carbone CO2 dans la biomasse végétale en initiant le cycle de Calvin, grâce à l'énergie solaire captée par la chlorophylle. Elle catalyse aussi bien la carboxylation que l'oxydation du ribulose-1,5-bisphosphate.
| IUBMB | Entrée IUBMB |
|---|---|
| IntEnz | Vue IntEnz |
| BRENDA | Entrée BRENDA |
| KEGG | Entrée KEGG |
| MetaCyc | Voie métabolique |
| PRIAM | Profil |
| PDB | RCSB PDB PDBe PDBj PDBsum |
| GO | AmiGO / EGO |
Sous ses différentes formes elle est probablement la protéine la plus abondante sur Terre[2] - [3], où elle représente environ 50 % des protéines solubles dans les feuilles des plantes en C3 et 30 % dans celles en C4 (respectivement 20 à 30 % et 5 à 9 % de l'azote contenu dans ces feuilles)[3]. En raison de sa prépondérance, cette enzyme joue un rôle essentiel dans le cycle du carbone de notre planète : de nombreuses archées et bactéries fixent en effet le carbone par le cycle du 3-hydroxypropionate ou par le cycle de Krebs inverse, mais ne comptent que pour une faible part dans le total ; la phosphoénolpyruvate carboxylase est également capable de fixer le carbone dans le métabolisme acide crassulacéen et dans celui des plantes en C4, mais en permettant son transport pour fixation définitive par la Rubisco.
Structure
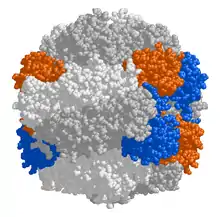
Chez les plantes, les algues, les cyanobactéries et les protéobactéries phototrophes et chimiotrophes, la Rubisco est généralement constituée de deux types de sous-unités : de grandes sous-unités (dites L pour l'anglais large) de 55 kDa et de petites sous-unités (dites S pour small) de 13 kDa[4]. D'une manière générale, les petites sous-unités sont codées par le matériel génétique du noyau et sont importées dans le stroma des chloroplastes à travers les membranes externe et interne de cet organite[2] - [5]. Une Rubisco complète compte typiquement huit sous-unités L et huit sous-unités S, formant un complexe protéique d'environ 540 kDa.
Les sites actifs de l'enzyme se trouvent sur des dimères de sous-unités L, chacune des deux sous-unités du dimère contribuant à la formation du site actif. Certains dinoflagellés et certaines protéobactéries ont d'ailleurs une Rubisco constituée d'un simple dimère de sous-unités L[6].
Des cations Mg2+ sont nécessaires pour assurer l'activité enzymatique de la protéine. Le positionnement correct de ces cations nécessite également une molécule de dioxyde de carbone « d'activation », qui n'est pas métabolisée mais forme un carbamate avec un résidu de lysine du site actif ; la formation de carbamate est favorisée à pH basique. La concentration en Mg2+ et le pH augmentent dans le stroma des chloroplastes sous l'effet de la lumière.
Activité
Activité carboxylase
La carboxylation catalysée par la Rubisco peut s'écrire de façon simplifiée (et approchée compte tenu du fait que tous ces composés sont ionisés dans le cytosol) comme suit :
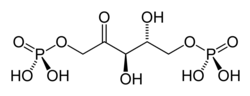
Le mécanisme réactionnel fait en réalité intervenir un énol, carboxylé en 3-céto-2-carboxyarabinitol-1,5-bisphosphate (3-KCABP) aussitôt hydrolysé en deux trioses-phosphate :
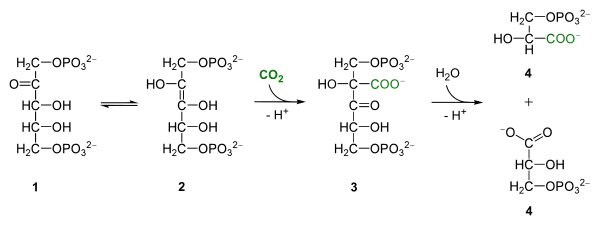 Addition de CO2 sur du ribulose-1,5-bisphosphate (1) donnant deux molécules de 3-phosphoglycérate (4) via un énol (2) carboxylé en 3-céto-2-carboxyarabinitol-1,5-bisphosphate (3), clivé par hydrolyse.
Addition de CO2 sur du ribulose-1,5-bisphosphate (1) donnant deux molécules de 3-phosphoglycérate (4) via un énol (2) carboxylé en 3-céto-2-carboxyarabinitol-1,5-bisphosphate (3), clivé par hydrolyse.
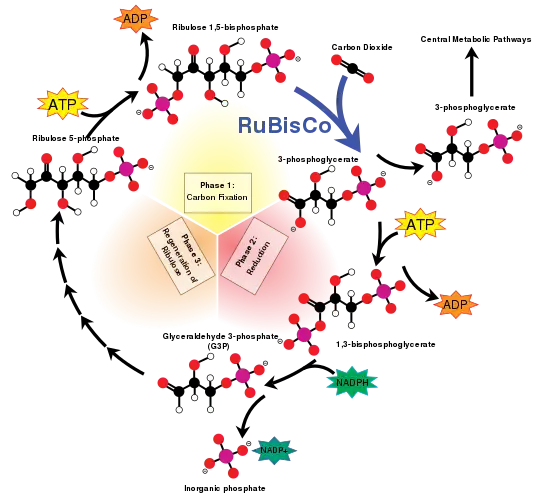
Activité oxygénase
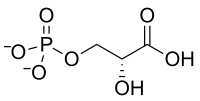
L'oxygénation catalysée par la Rubisco peut s'écrire de façon simplifiée comme suit :
Le 2-phosphoglycolate produit par cette réaction est recyclé à travers la photorespiration, qui fait intervenir des enzymes situées dans les mitochondries et les peroxysomes : deux molécules de phosphoglycolate sont converties en CO2 et en 3-phosphoglycérate, qui peut être à son tour recyclé dans le cycle de Calvin. Une partie du phosphoglycolate peut être également convertie en glycine.
Importance respective des activités carboxylase et oxygénase
Aux concentrations normales de dioxyde de carbone et d'oxygène dans l'atmosphère terrestre, le rapport entre l'activité carboxylase et l'activité oxygénase de la Rubisco est d'environ 4:1. Ce rapport diminue lorsque la température augmente, ce qui a favorisé des adaptations métaboliques particulières chez certaines plantes tropicales (plantes dites « en C4 ») et certaines plantes grasses (métabolisme acide crassulacéen, ou « CAM ») pour concentrer le CO2 autour de la Rubisco :
- les « plantes en C4 » fixent le CO2 sur le phosphoénolpyruvate dans le mésophylle par l'action successive de l’anhydrase carbonique et de la phosphoénolpyruvate carboxylase pour donner de l'oxaloacétate, ultérieurement converti en malate, lequel migre ensuite vers les cellules de la gaine périvasculaire pour y libérer le CO2 afin d'être métabolisé par la Rubisco sans être trop concurrencé par l'oxygène.
- les « plantes CAM » disposent du même matériel enzymatique mais accumulent le malate pendant la nuit pour le métaboliser par la Rubisco pendant la journée, lorsque les stomates sont fermés pour limiter les pertes d'eau par transpiration.
La Rubisco n'est pas une enzyme très rapide : chacune d'elles ne fixe qu'entre trois et dix molécules de CO2 par seconde[7]. En raison de cette lenteur relative, la Rubisco est l'enzyme limitante du cycle de Calvin pendant la journée. Cependant, dans la plupart des cas, et lorsque la lumière ne limite pas par ailleurs l'efficacité de la photosynthèse, la vitesse de la Rubisco répond positivement à l'augmentation de la concentration en CO2. Dans la mesure où la Rubisco réagit d'abord avec le ribulose-1,5-bisphosphate pour produire un ènediol puis avec le CO2 après quelques changements conformationnels pour produire le 3-phosphoglycérate, il est possible d'élaborer des modèles biochimiques rendant compte de la cinétique de cette enzyme sur la base de ces réactions[8].

| Les enzymes suivantes sont représentées :
PGP : phosphoglycolate phosphatase ; GOX : glycolate oxydase ; GGT : glycine transaminase ; GDC : système de clivage de la glycine ; SHMT : sérine hydroxyméthyltransférase ; SGT : sérine-glyoxylate transaminase ; HPR : hydroxypyruvate réductase ; GLYK : glycérate kinase ; |
Les numéros indiquent les métabolites suivants :
1 ribulose-1,5-bisphosphate, 2 3-phosphoglycérate, 3 2-phosphoglycolate, 4 glycolate, 5 glyoxylate, 6 glycine, 7 sérine, 8 hydroxypyruvate, 9 glycérate. |
Régulation
La Rubisco n'est normalement active que pendant la journée, car c'est dans la journée seulement qu'est produit le ribulose-1,5-bisphosphate en raison de la régulation de plusieurs autres enzymes du cycle de Calvin. Plusieurs autres facteurs contribuent à coordonner l'activité de la Rubisco avec celle des autres enzymes de ce cycle.
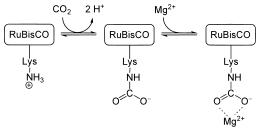
Régulation ionique
Le pH du stroma des chloroplastes passe de 7,0 à 8,0 pendant l'illumination diurne sous l'effet du gradient de protons généré à travers les membranes des thylakoïdes. Ce processus a pour effet de concentrer les protons à l'intérieur des thylakoïdes, et donc d'en diminuer la concentration dans le stroma. Parallèlement, les cations de magnésium Mg2+ sont pompés hors des thylakoïdes et s'accumulent dans le stroma. La Rubisco, dont le pH optimal peut dépasser 9,0 selon la concentration en Mg2+, est par conséquent activée par fixation de CO2 avec l'aide de ce cation sur un résidu de lysine du site actif de l'enzyme pour y former un carbamate indispensable à la réaction.
Régulation par la Rubisco activase
Chez les plantes et certaines algues, une enzyme spécifique, la Rubisco activase, est nécessaire pour catalyser la formation du carbamate avant que la Rubisco soit bloquée par la liaison du substrat[9] - [10], lequel se lie plus fortement à l'enzyme inactive (sans carbamate) qu'à l'enzyme carbamylée : la Rubisco activase libère le ribulose-1,5-bisphosphate lié à la Rubisco inactive, permettant l'activation de cette dernière par le dioxyde de carbone. Cette enzyme est également nécessaire à certaines plantes (notamment Nicotiana tabacum et certains haricots) dont le métabolisme produit un inhibiteur compétitif, le 2-carboxy-D-arabinitol-1-phosphate (CA1P), lequel se lie à la Rubisco carbamylée en bloquant son site actif[11] ; la Rubisco activase libère le CA1P du site actif de la Rubisco, et une autre enzyme, la 2-carboxyarabinitol-1-phosphatase, déphosphoryle le CA1P en 2-carboxy-D-arabinitol. Cette opération doit être renouvelée régulièrement — environ à chaque minute — lorsqu'une nouvelle molécule de CA1P vient bloquer la Rubisco.
Le CA1P pourrait stabiliser la Rubisco dans une conformation limitant les risques de protéolyse[12]. Les propriétés de la Rubisco activase tendent à limiter le potentiel photosynthétique des plantes à température et concentration de CO2 élevées[13]. Par ailleurs, à température élevée, la Rubisco activase tend à former des agrégats inactifs qui ne sont plus en mesure de maintenir l'activité de la Rubisco, ce qui explique le ralentissement de l'activité carboxylase en cas de stress thermique[14].
Régulation de l'activase par le ratio ATP/ADP et par l'état redox de la thiorédoxine du stroma
L'élimination du 2-carboxy-D-arabinitol-1-phosphate (CA1P) par la Rubisco activase consomme de l'ATP et est inhibée par l'ADP. L'activité de l'activase est donc déterminée par le ratio ATP/ADP, mais la sensibilité de cette enzyme à ce ratio est elle-même conditionnée, dans la plupart des plantes, par l'état rédox moyen des molécules de thiorédoxine dans le stroma des chloroplastes. Le niveau d'activité de l'activase, et donc de la Rubisco, dépend ainsi de l'intensité lumineuse, ce qui permet de la corréler à la formation du ribulose-1,5-bisphosphate[15].
Régulation par le phosphate inorganique
Chez les cyanobactéries, le phosphate inorganique se lie au site actif de la Rubisco ainsi qu'à un autre site sur les grandes sous-unités, modulant ainsi les transitions entre états actif et inactif de la Rubisco. Le phosphate inorganique pourrait jouer, chez ces bactéries, un rôle semblable à celui de la Rubisco activase des plantes supérieures[16].
Régulation par le dioxyde de carbone
Le dioxyde de carbone CO2 et l'oxygène O2 étant en compétition dans le site actif de la Rubisco, l'efficacité de l'enzyme quant à la fixation du CO2 peut être améliorée par accroissement de la concentration en CO2 dans le stroma des chloroplastes, où se trouve la Rubisco. Cela a fait l'objet de certaines adaptations métaboliques à l'œuvre dans les plantes dites « en C4 » et « CAM ». Le fait que l'oxygène soit en concurrence avec le dioxyde de carbone peut surprendre dans la mesure où la photorespiration aboutit à perdre de l'énergie fixée par photosynthèse et peut même annuler le flux net de CO2 fixé par la Rubisco dans des feuilles saines en plein lumière.
La vitesse relative des activités carboxylase et oxygénase de la Rubisco dépendent en premier lieu de la température, car une température élevée réduit le taux de CO2 dissous dans les tissus et favorise donc l'activité oxygénase, c'est-à-dire la photorespiration. Or la température dépend de l'hydratation de la plante, car c'est la transpiration qui permet de refroidir les feuilles.
Les plantes en C4 utilisent la phosphoénolpyruvate carboxylase, une décarboxylase qui a une meilleure affinité pour le CO2 que la Rubisco, afin de catalyser l'addition d'un anion bicarbonate HCO3− sur du phosphoénolpyruvate pour donner de l'oxaloacétate, lequel est ensuite converti en malate et transporté pour être décarboxylé en libérant du CO2 là où se déroule le cycle de Calvin, dans un environnement moins riche en oxygène et donc favorable à l'activité carboxylase de la Rubisco.
Histoire évolutive
La Rubisco est un catalyseur relativement peu efficace en raison de sa lenteur (1 à 9 cycles catalytiques par seconde), de sa faible affinité pour CO2 et de sa faible spécificité pour CO2 par rapport à O2. La baisse de concentration de CO2 et l'augmentation concomitante de celle de O2 dans l'atmosphère terrestre ancienne ont conduit au développement, indépendamment dans diverses lignées d'organismes photosynthétiques, de mécanismes de concentration du dioxyde de carbone au voisinage du site actif et de raréfaction de l'oxygène[17].
Les isoformes de type I de la superfamille des Rubisco sont présentes dans les plantes terrestres, les algues et les bactéries. En raison des grandes lacunes dans le registre fossile ancien, l'histoire évolutive de ces isoformes est difficile à reconstituer. En 2022, la comparaison des formes actuelles et des expérimentations sur Escherichia coli ont néanmoins permis de montrer que la Rubisco ancestrale ne comportait que l'octamère L8 et que le recrutement des sous-unités S a quadruplé l'efficacité catalytique et doublé la spécificité pour CO2 vis-à-vis de O2. Cet événement a dû intervenir avant ou au début du grand événement d'oxygénation il y a 2,4 milliards d'années[17] - [18].
Optimisation de la Rubisco par génie génétique
Le « rendement photosynthétique » d'une cellule se définit comme la fraction d'énergie lumineuse convertie par cette cellule en énergie chimique. Plus il est élevé, et plus la plante est en mesure d'absorber du CO2 pour fabriquer des glucides et, in fine, toute une gamme de biomolécules constituant notamment sa physiologie. La Rubisco est le facteur limitant du rendement photosynthétique global des organismes qui reposent sur cette enzyme ; en accroître l'efficacité catalytique — en augmentant celle de l'activité carboxylase et/ou en réduisant celle de l'activité oxygénase — pourrait donc favoriser la séquestration écologique ou naturelle du CO2 par voie biologique et constituer une option valable dans le contrôle du taux atmosphérique des gaz à effet de serre en partie responsables du réchauffement climatique. Des généticiens estiment que le rendement de certaines plantes pourrait ainsi être dopé de 40 %.
Des expériences ont d'abord porté sur l'expression de gènes de Rubisco d'une espèce transférés par transgenèse dans une plante d'une autre espèce. La stimulation de l'expression de certaines sous-unités de la Rubisco a été testée, de même que l'expression des gènes des sous-unités S directement depuis les plastomes (c'est-à-dire le matériel génétique chloroplastique), voire la modification des gènes codant les sous-unités de la Rubisco pour en augmenter la spécificité au CO2 ou en accroître la vitesse de fixation du carbone. Une voie prometteuse serait d'introduire, dans des plantes cultivées, des gènes de Rubisco plus performante (ex : ceux de rhodophytes telles que Galdieria partita) ; La Rubisco du tabac a ainsi été remplacée par celle d'une protéobactérie, Rhodospirillum rubrum. Ces OGM ont poussé plus haut et produit environ 40 % de biomasse supplémentaire ; d'autres tests devraient porter sur des plantes alimentaires riches en protéines et/ou amidon (soja, cornille (niébé), riz ou pomme de terre)[19].
Il reste à montrer que les changements induits n'affectent pas négativement la plante, car cette amélioration de la Rubisco est basée sur une réduction de la photorespiration, or cette dernière, si elle présente l'inconvénient de faire consommer de l'énergie à la plante, semble aussi avoir un rôle dans la signalisation cellulaire végétale, dans le processus de production de peroxyde d'hydrogène, molécule impliquée dans la signalisation rédox, impliquée dans les mécanismes de croissance mais aussi de réponse à certains stress (attaques de microbes ou parasites)[20].
Utilisation alimentaire
Parce que cette protéine Rubisco est très abondante dans la nature et qu'elle contient tous les acides aminés essentiels, elle est un aliment potentiellement intéressant. Une méthode a été développée pour l'extraire et la concentrer afin d'en faire des "steaks végétaux"[21].
Notes et références
Notes
- Le terme « Rubisco » a été forgé en 1979 par le biochimiste américain David S. Eisenberg lors d'un séminaire marquant le départ à la retraite de Samuel G. Wildman, éminent spécialiste de cette enzyme. L'abréviation dérive de l'appellation anglophone « ribulose-1,5-bisphosphate carboxylase/oxygenase, » avec un clin d'œil aux snacks Nabisco en souvenir de la tentative malheureuse de Wildman de créer des feuilles de tabac comestibles[1].
Références
- (en) Samuel G. Wildman, « Along the trail from Fraction I protein to Rubisco (ribulose bisphosphate carboxylase-oxygenase) », Photosynthesis Research, vol. 73, nos 1-3, , p. 243-250 (PMID 16245127, DOI 10.1023/A:1020467601966)
- (en) Amit Dhingra, Archie R. Portis Jr. et Henry Daniell, « Enhanced translation of a chloroplast-expressed RbcS gene restores small subunit levels and photosynthesis in nuclear RbcS antisense plants », Proceedings of the National Academy of Sciences of the United States of America, vol. 101, no 16, , p. 6315-6320 (PMID 15067115, PMCID 395966, DOI 10.1073/pnas.0400981101, Bibcode 2004PNAS..101.6315D, lire en ligne)
- (en) Urs Feller, Iwona Anders et Tadahiko Mae, « Rubiscolytics: fate of Rubisco after its enzymatic function in a cell is terminated », Journal of Experimental Botany, vol. 59, no 7, , p. 1615-1624 (PMID 17975207, DOI 10.1093/jxb/erm242, lire en ligne)
- (en) Le gène des sous-unités L fait partie du génome des chloroplastes chez les plantes (Entrez GeneID: )
- (en) M. Yoon, J. J. Putterill, G. S. Ross, W. A. Laing, « Determination of the relative expression levels of rubisco small subunit genes in Arabidopsis by rapid amplification of cDNA ends », Anal. Biochem., vol. 291, no 2, , p. 237–244 (DOI 10.1006/abio.2001.5042, lire en ligne)
- (en) T. Lundqvist et G. Schneider, « Crystal structure of activated ribulose-1,5-bisphosphate carboxylase complexed with its substrate, ribulose-1,5-bisphosphate », Journal of Biological Chemistry, , p. 12604-12611 (PMID 1905726, lire en ligne)
- (en) R. John Ellis, « Biochemistry: Tackling unintelligent design », Nature, vol. 463, no 7278, , p. 164-165 (PMID 20075906, DOI 10.1038/463164a, lire en ligne)
- (en) Hadi Farazdaghi, « The single-process biochemical reaction of Rubisco: A unified theory and model with the effects of irradiance, CO2 and rate-limiting step on the kinetics of C3 and C4 photosynthesis from gas exchange », Biosystems, vol. 103, no 2, , p. 265-284 (PMID 21093535, DOI 10.1016/j.biosystems.2010.11.004, lire en ligne)
- (en) Archie R. Portis Jr., « Rubisco activase – Rubisco's catalytic chaperone », Photosynthesis Research, vol. 75, no 1, , p. 11-27 (PMID 16245090, DOI 10.1023/A:1022458108678, lire en ligne)
- (en) JIN Song-heng, JIANG De-an, LI Xue-qin et SUN Jun-wei, « Characteristics of photosynthesis in rice plants transformed with an antisense Rubisco activase gene », Journal of Zhejiang University SCIENCE, vol. 5, no 8, , p. 897-899 (PMID 15236471, DOI 10.1631/jzus.2004.0897, lire en ligne)
- (en) P. John Andralojc, Glenn W. Dawson, Martin A. J. Parry et Alfred J. Keys, « Incorporation of carbon from photosynthetic products into 2-carboxyarabinitol-1-phosphate and 2-carboxyarabinitol », Biochemical Journal, vol. 304, no Pt 3, , p. 781-786 (PMID 7818481, PMCID 1137402, lire en ligne)
- (en) Shahnaz Khan, P. John Andralojc, Peter J. Lea et Martin A. J. Parry, « 2′-Carboxy-D-arabitinol 1-phosphate protects ribulose 1,5-bisphosphate carboxylase/oxygenase against proteolytic breakdown », European Journal of Biochemistry / FEBS, vol. 266, no 3, , p. 840-847 (PMID 10583377, DOI 10.1046/j.1432-1327.1999.00913.x, lire en ligne)
- (en) Steven J. Crafts-Brandner et Michael E. Salvucci, « Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2 », Proceedings of the National Academy of Sciences of the United States of America, vol. 97, no 24, , p. 13430-13435 (PMID 11069297, PMCID 27241, DOI 10.1073/pnas.230451497, Bibcode 2000PNAS...9713430C, lire en ligne)
- (en) Michael E. Salvucci, Katherine W. Osteryoung, Steven J. Crafts-Brandner et Elizabeth Vierling, « Exceptional Sensitivity of Rubisco Activase to Thermal Denaturation in Vitro and in Vivo », Plant Physiology, vol. 127, no 3, , p. 1053-1064 (PMID 11706186, PMCID 129275, DOI 10.1104/pp.010357, lire en ligne)
- (en) Ning Zhang, Russell P. Kallis, Robert G. Ewy et Archie R. Portis Jr, « Light modulation of Rubisco in Arabidopsis requires a capacity for redox regulation of the larger Rubisco activase isoform », Proceedings of the National Academy of Sciences of the United States of America, vol. 99, no 5, , p. 3330-3334 (PMID 11854454, PMCID 122518, DOI 10.1073/pnas.042529999, Bibcode 2002PNAS...99.3330Z, lire en ligne)
- (en) Yehouda Marcus et Michael Gurevitz, « Activation of cyanobacterial RuBP-carboxylase/oxygenase is facilitated by inorganic phosphate via two independent mechanisms », European Journal of Biochemistry, vol. 267, no 19, , p. 5995-6003 (PMID 10998060, DOI 10.1046/j.1432-1327.2000.01674.x, lire en ligne)
- (en) Robert E. Sharwood, « Reconstructing CO2 fixation from the past », Science, vol. 378, no 6616, , p. 137-138 (DOI 10.1126/science.ade6522).
- (en) Luca Schulz, Zhijun Guo, Jan Zarzycki, Wieland Steinchen, Jan M. Schuller et al., « Evolution of increased complexity and specificity at the dawn of form I Rubiscos », Science, vol. 378, no 6616, , p. 155-160 (DOI 10.1126/science.abq1416).
- Scientists engineer shortcut for photosynthetic glitch, boost crop growth by 40 percent ; publié le 03 janvier 2019, Université de l'Illinois (Urbana-Champaign). phys.org/news ; URL https://phys.org/news/2019-01-scientists-shortcut-photosynthetic-glitch-boost.html#jCp
- Christine H. Foyer, Arnold J. Bloom, Guillaume Queval et Graham Noctor, « Photorespiratory Metabolism: Genes, Mutants, Energetics, and Redox Signaling », Annual Review of Plant Biology, vol. 60, no 1, , p. 455–484 (ISSN 1543-5008, DOI 10.1146/annurev.arplant.043008.091948, lire en ligne, consulté le )
- René Didde, « Nutrition. Vous reprendrez bien un peu de steak d’herbe ? », Courrier International, (lire en ligne)