Chlorophylle
Les chlorophylles (mot composé en 1816 à partir du grec ancien χλωρός / khlôrós (« vert ») et φύλλον / phúllon (« feuille ») sont les principaux pigments assimilateurs des végétaux photosynthétiques.
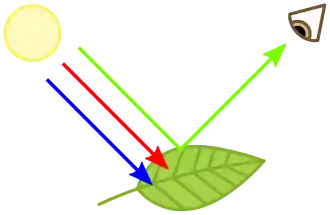



Leur nom est introduit par Joseph Bienaimé Caventou et Joseph Pelletier en 1816[1]. Ces pigments, situés dans les chloroplastes des cellules végétales, interviennent dans la photosynthèse pour intercepter l'énergie lumineuse, première étape dans la conversion de cette énergie en énergie chimique. Leurs spectres d'absorption du rayonnement lumineux sont responsables de la couleur verte des végétaux ; la longueur d'onde la moins absorbée étant le vert, cette couleur est perçue dans la lumière réfléchie vers l'œil par la feuille.
L'assimilation chlorophyllienne est l'assimilation active du dioxyde de carbone par voie de photosynthèse au moyen de la chlorophylle. Elle est un processus biologique de transformation de l’énergie lumineuse en énergie chimique de liaison grâce aux chloroplastes chlorophylliens. La synthèse chlorophyllienne est la forme la plus fréquente de photosynthèse.
Différentes formes
La chlorophylle peut avoir plusieurs structures chimiques, les deux principales étant :
- les chlorophylles a et a' (symbole : « chla » ) sont les pigments photosynthétiques les plus communs du règne végétal ; elles sont présentes chez tous les végétaux aquatiques et terrestres (≈ 3 g/kg de feuilles fraîches). La mesure de leur concentration dans l'eau est utilisée comme indicateur de la quantité de plancton végétal (phytoplancton, base principale du réseau trophique aquatique). Les taux de l'eau en chlorophylle sont donnés en µg chla/L.
- la chlorophylle b (en réalité b et b') se trouve chez les cormophytes (végétaux supérieurs) et les chlorophycées (algues vertes) à des teneurs moindres (≈ 0.75 g/kg MF).
Trois autres formes sont moins communes :
- les chlorophylles c (c1, c2) : identifiée chez les phéophycées (dites algues brunes),
- la chlorophylle d : identifiée en 1943 chez les cyanobactéries,
- la chlorophylle f : identifiée en 2010 dans certains stromatolithes ; elle a pour caractéristique une absorption décalée vers le rouge par rapport aux autres chlorophylles[2].
La chlorophylle se caractérise notamment par sa fluorescence dans le proche infrarouge (autour de 700 nm). Les clichés aériens ou satellitaires en « fausses couleurs » (infrarouge + vert + bleu) permettent aux spécialistes de reconnaître les essences par analyse radiométrique.
Structure chimique et biosynthèse
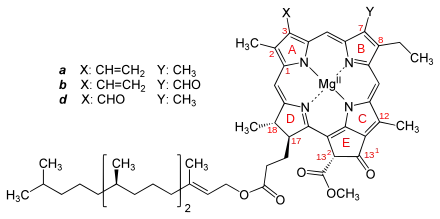
La chlorophylle est une chlorine chélatant un cation de magnésium Mg2+ au centre du macrocycle et estérifiant — hormis les chlorophylles c — un alcool terpénoïde en C20, le phytol, qui est hydrophobe et sert d'ancrage à des protéines des membranes thylacoïdes[3]. Elle présente une structure quasi identique à celle de l'hème des érythrocytes, les globules rouges du sang. C'est la présence, dans sa structure, de nombreuses doubles liaisons conjuguées qui permet une interaction avec le rayonnement lumineux et son absorption. Les chaînes latérales de la chlorine sont variables et ceci entraîne une modification du spectre d'absorption entre les différentes familles de chlorophylles.
Dans le tableau ci-dessus, les cases colorées en jaune soulignent les différences avec la chlorophylle a prise comme référence.
 Structure de la chlorophylle d. |
 Structure de la chlorophylle f. |

Un déficit de magnésium dans le sol ou dans l'eau affecte donc directement la biosynthèse des chlorophylles. La quantité de pigment produite est dans ce cas plus faible et, chez les plantes terrestres, les nouvelles feuilles sont alors vert-pâle, voire jaunes. Une manifestation de la carence, plus ou moins prononcée, en minéraux affectant la teneur finale en chlorophylle est appelée chlorose.
La chlorophylle, faiblement soluble dans l'eau, l'est en revanche bien davantage dans l'éthanol.
Spectre d’absorption et origine de la couleur verte des végétaux
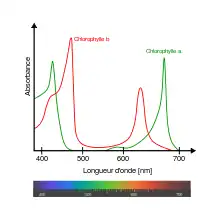
La chlorophylle 'a (comme tous les pigments) est fluorescente. La mesure de la fluorescence de la chlorophylle a (émise par une plante ou un organisme photosynthétique en général) est un puissant outil pour mesurer l'absorption de la lumière et le fonctionnement de la photosynthèse.
Le spectre visible se situe approximativement entre 380 nm à 780 nm bien qu'une gamme de 400 nm à 700 nm soit plus commune. La lumière perçue comme « verte » par l’œil et le cerveau humain a une longueur d'onde, selon les notions de la couleur « verte », approximativement entre 490 et 570 nanomètres.
On remarque sur le graphique que l’absorbance de la chlorophylle est moindre pour cette plage du spectre électromagnétique. La chlorophylle absorbe donc la majeure partie du spectre visible sauf la lumière verte (notion de green window, de « vide vert » ou « fenêtre du vert »). Si cette chlorophylle absorbait de manière optimale toutes les régions du spectre, les végétaux paraîtraient noirs, même en plein jour. Ce défaut d'absorption peut être interprété comme une stratégie optimale d'allocation des ressources : les plantes privilégient la biosynthèse des pigments qui utilisent le spectre de lumière optimal (dans le rouge et le bleu) au lieu d'investir dans un équipement pigmentaire plus complet mais au coût du captage du rayonnement solaire trop élevé[alpha 1] (complexité des assemblages protéiques nécessaires à la synthèse des différentes chlorophylles)[7]. Une autre hypothèse formulée par le biologiste Andrew Goldsworthy[8] fait jouer un rôle important aux bactéries pourpres sulfureuses. Ces micro-organismes ont développé un système primitif de photosynthèse utilisant un pigment de couleur pourpre, la bactériorhodopsine, peu sélectif[9], ce qui leur permettait d'absorber l'énergie solaire, facilitant l'assimilation des éléments nutritifs carbonés de la soupe primitive. Mais ce système n'absorbait pas le dioxyde de carbone, ce qui conduisit au déclin des bactéries pourpres car la prolifération des bactéries primitives à fermentation a progressivement épuisé la matière organique de la soupe primordiale. Des cyanobactéries vivant dans l'ombre des bactéries sulfureuses, dans une lumière filtrée qui ne laissait que les extrémités du spectre de lumière visible disponibles (la lumière rouge et la lumière bleue), auraient sélectionné ces zones d'absorption et délaissé le centre du spectre de la lumière visible, soit le vert[10].
Ce défaut est à l'origine d'un autre phénomène : l'ombre « verte » qui baigne les sous-bois. Les arbres ne laissant filtrer que très peu de lumière solaire (3 % au maximum pour le hêtre), il devrait faire extrêmement sombre sous leurs frondaisons, même en plein jour. Le rayonnement entre 490 et 570 nanomètres n'étant pas filtré, il parvient jusqu'au sol et donne la pénombre « verte », avec un effet apaisant sur les sens, utilisé notamment en sylvothérapie[11].
La lumière rouge a une longueur d'onde de 620-750nm et une fréquence de 400-484THz. La région du rouge atteint un maximum de 660-670 nm pour la chlorophylle a et aux alentours de 635-645 nm pour la chlorophylle b. Les plantes ont un grand besoin des ondes rouges sauf celles beaucoup plus longues que 670 nm.
Le fait que les chlorophylles absorbent très peu la lumière verte laisse supposer à tort que les radiations vertes ont peu d'importance dans la photosynthèse. Or les valeurs d'absorption des feuilles dans le vert (valeurs comprises entre 50 et 90 %) sont bien plus importantes que ne le laisseraient supposer les spectres d’absorption des chlorophylles qu'elles contiennent. Les radiations rouges et bleues sont largement absorbées dans la première couche cellulaire et le chemin qu’elles parcourent dans la feuille est faible, tandis que les radiations vertes sont transmises et surtout réfléchies à la surface des cellules, des chloroplastes. Le résultat est un allongement du chemin qu’elles parcourent dans la feuille, augmentant la probabilité de leur absorption par les chlorophylles[12]. Il semble ainsi que la palette de pigments complémentaires associés à la collecte de la lumière solaire a été sélectionnée au cours de l'évolution des plantes vasculaires afin de maximiser l'utilisation des radiations vertes[13].
Mise en évidence
On peut montrer que la chlorophylle est nécessaire pour la photosynthèse en utilisant une feuille d'une plante panachée (les parties blanches de la feuille sont dépourvues de chlorophylle) où l'on aura préalablement digéré l'amidon. Après une exposition de quelques heures à la lumière et l'utilisation d'une solution iodée, une coloration noire apparaît seulement sur les zones qui étaient vertes et contenaient de la chlorophylle.
Notes et références
Notes
- Les végétaux investissent jusqu'à un tiers de leur photosynthèse nette dans la synthèse du système de captage du rayonnement[6].
Références
- Hervé This, « Parlons des chlorophylles, et pas de la chlorophylle ! », sur Académie d'Agriculture de France (consulté le ).
- Min Chen, Schliep M, Willows RD et Als, A red-shifted chlorophyll, Science, 2010:329;1318-1319
- Biologie Végétale 2e édition
- Anthony W. D. Larkum, Susan E. Douglas et John A. Raven, « Photosynthesis in algae », Kluwer, Londres, 2003. (ISBN 0-7923-6333-7).
- (en) Dennis J. Nürnberg, Jennifer Morton, Stefano Santabarbara, Alison Telfer, Pierre Joliot et al., « Photochemistry beyond the red limit in chlorophyll f–containing photosystems », Science, vol. 360, , p. 1210-1213 (DOI 10.1126/science.aar8313).
- (en) John Albert Raven, « A cost-benefit analysis of photon absorption by photosynthetic unicells », New Phytologist, vol. 98, no 4, , p. 593-625 (DOI 10.1111/j.1469-8137.1984.tb04152.x).
- (en) Marcell A Marosvölgyi & Hans van Gorkom, « Cost and color of photosynthesis », Photosynth Res., vol. 103, no 2, , p. 105–109 (DOI 10.1007/s11120-009-9522-3).
- Chercheur à l'Imperial College London.
- Ce système primitif de photosynthèse absorbe une large proportion du spectre de la lumière visible.
- (en) Andrew Goldsworthy, « Why trees are green », New Scientist, vol. 116, no 1590, , p. 48–51.
- Peter Wohlleben, La Vie secrète des arbres. Ce qu'ils ressentent. Comment ils communiquent, Les Arènes, (lire en ligne), p. 87.
- (en) R. A. Moss & W. E. Loomis, « Absorption Spectra of Leaves. I. The Visible Spectrum », Plant Physiology, vol. 27, no 2, , p. 370-391.
- (en) Jindong Sun, John N. Nishio & Thomas C. Vogelmann, « Green light drives CO2 deep within leaves », Plant Cell Physiol, vol. 39, no 10, , p. 1020-1026.
Voir aussi
Bibliographie
- P. H. Raven, R. F. Evert et S. E. Eichhorn (trad. de l'anglais par Jules Bouharmont, révision scientifique de Carlo Evrardt), Physiologie Végétale, Bruxelles, 2, coll. « Editions De Boeck Université », , 121 p. (ISBN 978-2-8041-5020-4)