Réseau trophique
Un réseau trophique est un ensemble de chaînes alimentaires reliées entre elles au sein d'un écosystème et par lesquelles l'énergie et la biomasse circulent (échanges d'éléments tels que le flux de carbone et d'azote entre les différents niveaux de la chaîne alimentaire, échange de carbone entre les végétaux autotrophes et les hétérotrophes).
Le terme trophique se rapporte à tout ce qui est relatif à la nutrition d'un tissu vivant ou d'un organe. Par exemple, une relation trophique est le lien qui unit le prédateur et sa proie dans un écosystème.
Dans un écosystème, la structure des réseaux trophiques (les types et réseaux de relations alimentaires entre organismes) influence fortement la quantité, la diversité, la stabilité et la qualité de la biomasse et de la matière organique résiduelle (excrétions, nécromasse) produites par les écosystèmes[1]. La qualité d'un réseau trophique et de ses interactions a un lien direct avec la stabilité et la résilience écologique des populations qui en font partie[2].
Les réseaux trophiques, parfois très complexes, sont étudiés par l'« écologie trophique ». Ils sont affectés par les changements globaux, dont ceux liés au réchauffement climatique[3] - [4], dont au niveau des espèces clé de voûte[5].
Description
La notion de réseau trophique désigne l'ensemble des relations trophiques existant à l'intérieur d'une biocénose entre les diverses catégories écologiques d'êtres vivants constituant cette dernière (producteurs, consommateurs, décomposeurs). Il peut aussi se définir comme étant la résultante de l'ensemble des chaînes alimentaires unissant les diverses populations d'espèces que comporte une biocénose.
Aux échelles humaines de temps, la structure de la communauté peut sembler en équilibre. Il s'agit en fait d'un équilibre instable maintenu en équilibre dynamique, par le jeu de deux grandes chaines de rétrocontrôles ; les relations « top-down » (régulation des ressources par leurs consommateurs) et « bottom-up » (rétroactions des ressources sur leurs consommateurs).
Mieux comprendre ces interactions est un des enjeux de l'écologie, car elles sont fortement affectées par les impacts en cascade des activités humaines (chasse, pêche, agriculture), qui diminuent fortement le nombre et la biomasse des espèces de haut niveau trophique, tout en augmentant ou perturbant les apports et les cycles de nutriments[6], conduisant notamment à des phénomènes d'appauvrissement en amont et de dystrophisation en aval des bassins versants, ou à des phénomènes d'eutrophisation et d'anoxie en mer[7].
Chaque réseau est caractérisé par des boucles de rétroactions complexes et des équilibres dynamiques, fortement influencées par les facteurs biotiques et abiotiques (température, pH, disponibilité en lumière, oxygène et nutriments notamment).
Ainsi, à titre d'exemple pour les milieux aquatiques, l'augmentation du nombre d'individus d'une espèce ou du nombre d'espèces herbivores tend à diminuer le nombre d'algues fixées ou en suspension. Mais dans le même temps cette pression herbivore favorise l'arrivée de lumière et rend disponible des nutriments, ce qui favorise la croissance d'autres algues ou plantes[8] - [9].
Inversement, un déséquilibre écologique, tel que la pullulation en surface ou en pleine eau d'une espèce envahissante introduite (n'ayant pas ou n'ayant plus de prédateur local) peut priver le milieu de lumière et le conduire à l'anoxie[10] voire à un phénomène de zone morte, ou une espèce envahissante, comme la moule zébrée (Dreissena polymorpha), en filtrant des masses considérables d'eau va induire un changement de la composition algale[11].
Selon les espèces herbivores (éventuellement exotiques et envahissantes) qui dominent le milieu, la pression herbivore peut être très sélective et alors induire d'importants changements dans le réseau trophique (changements de dynamiques démographiques et de colonisation, changement dans les tailles[12] ou dans le nombre d'espèces, etc.)[13].
Les relations inter-spécifiques, les structures en taille et biomasse de la communauté, les relations intraspécifiques des assemblages de plantes ou microalgues en seront affectées et modifient en retour la structure et la dynamique des herbivores et de leurs prédateurs[14].
Chaîne alimentaire

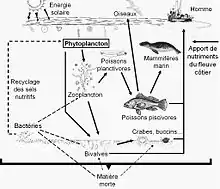
Une chaîne alimentaire est une suite d'êtres vivants de différents niveaux trophiques dans laquelle chacun mange des organismes de niveau trophique inférieur dans le but d'acquérir de l'énergie. Le premier maillon d'une chaîne est toujours un organisme autotrophe. Dans les mers et les océans, le phytoplancton assure ce rôle. Dans les profondeurs abyssales où les rayons du soleil ne parviennent pas, les bactéries thermophiles sont les premiers maillons de la chaîne. Cependant, la chaîne photosynthétique y existe tout de même ; en effet les organismes pélagiques meurent et coulent.
Selon une étude de 2013[15] - [16], le régime alimentaire de l'espèce humaine ne la situe pas au sommet de la chaîne alimentaire, mais au même niveau que les anchois ou les cochons. L'humain est un cas spécifique, il ne possède pas de prédateur sans pour autant être au sommet de la chaîne alimentaire. Les populations humaines avec un régime carné se rapprochent cependant des superprédateurs comme l'ours polaire.
Quoi qu'il en soit, essayer de classer l'humain dans la chaîne alimentaire semble être une impasse, car une chaîne alimentaire indique forcément une relation de dépendance avec le bas (les prédateurs étant démographiquement régulés par le nombre de proies présentes dans leur milieu naturel). Or, en dépit des extinctions massives qui touchent aujourd'hui la planète Terre, la démographie humaine n'en demeure aucunement affectée pour le moment. Plutôt qu'être au sommet de la chaîne alimentaire, l'espèce humaine a donc, en réalité, un rôle omniscient sur une chaîne alimentaire dont elle s'est extraite, cela depuis l'apparition de l'agriculture et de l'élevage au début du néolithique pour remodeler la chaîne alimentaire à son propre service.
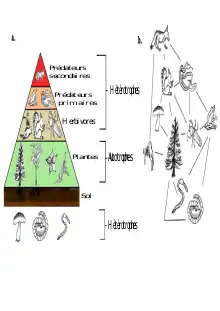
Dans un écosystème, les liens qui unissent les espèces sont le plus souvent d'ordre alimentaire. Ces relations forment des séquences où chaque individu mange le précédent et est mangé par celui qui le suit ; on parle de « chaîne alimentaire ». Chaque maillon est un niveau trophique.
Structure des réseaux trophiques
L’intérêt envers la structure des réseaux trophiques est né lors des recherches effectuées pour comprendre la relation entre la diversité et la stabilité des réseaux. Depuis, les recherches pour comprendre comment sont structurés ces réseaux — nombre de nœuds (espèces), nombre de liens trophiques, densité et organisation des liens trophiques — et quels en sont les processus à l'origine ont été nombreuses, notamment car la structure joue un rôle important dans la dynamique et la stabilité des écosystèmes. De plus, dans un contexte de perte de biodiversité et de changements globaux, comprendre comment sont structurées les communautés d'espèces sur la base des relations trophiques est devenu un enjeu important pour la conservation.
Composition
Les réseaux trophiques sont composés de plusieurs niveaux trophiques en commençant par les espèces basales, qui constituent le niveau le plus bas. Il contient tous les producteurs primaires que sont les espèces autotrophes. Viennent ensuite les herbivores, considérés comme les premiers consommateurs, se nourrissant des espèces basales. On retrouve ensuite les carnivores primaires (seconds consommateurs) et les carnivores secondaires (consommateurs tertiaires). Les taxons se trouvant à différents niveaux trophiques sont liés par des interactions plus ou moins fortes de type consommateur/consommé. Les liens d'un réseau trophique sont alors unidirectionnels. Ensemble, ils forment des chaînes trophiques reliées entre elles. Chaque écosystème a un réseau trophique dont la structure lui est propre. Les réseaux trophiques peuvent être plus ou moins compartimentés, par exemple, en milieu marin, on peut retrouver le compartiment benthique et le compartiment pélagique. Ces compartiments interagissent entre eux par des interactions plus faibles que les interactions au sein même du compartiment. Les réseaux peuvent aussi être plus ou moins complexes, avec un nombre de nœuds plus ou moins important, on parle alors de la diversité du réseau. De plus, le nombre de liens entre les nœuds peut aussi varier. Afin d'expliquer et de prédire les propriétés structurales des interactions trophiques entre ces espèces, de nombreux modèles universels ont été étudiés et décrits.
Réseaux et modèles
Un large éventail de réseaux, sociaux, économiques, technologiques ou encore biologiques, ont été décrits comme étant des réseaux « petit monde » (réseaux avec beaucoup de nœuds faiblement connectés et un petit nombre de nœuds très connectés) et « sans échelle » (la distribution des degrés de nœuds suit une loi de puissance). Cependant, l’appartenance des réseaux trophiques à ces classes de réseaux a été remise en cause par Dunne et al. (2006)[17], grâce à l’étude d’un panel de réseaux trophiques empiriques provenant de divers écosystèmes aquatiques et terrestres.
Les réseaux « petit monde » affichent un coefficient de regroupement plus élevé que les réseaux aléatoires, ce qui signifie que les voisins d’un nœud sont susceptibles d’être beaucoup plus connectés les uns aux autres dans un réseau « petit monde » que dans un graphe aléatoire. Ils possèdent également une plus petite longueur de trajet que dans un réseau régulier, semblable à ce qui est vu dans un réseau aléatoire, ce qui signifie que la longueur de trajet caractéristique (ou longueur de trajet la plus courte moyenne) augmente de manière logarithmique avec le nombre de nœuds. Or, la majorité des réseaux trophiques ont peu de nœuds par rapport à d’autres réseaux non – biologiques et leur coefficient de regroupement est alors similaire à celui d’un graphe aléatoire. La plupart des réseaux trophiques ne possèdent donc pas la topologie typique d’un réseau « petit monde ».
Les distributions de degrés de nœuds des réseaux trophiques, qui sont les distributions probabilistes des degrés de chaque sommet du réseau, différent souvent de celles d’un réseau aléatoire. Ainsi, les réseaux trophiques n’affichent pas toujours des distributions de degrés « sans échelle » mais présentent une variété de formes fonctionnelles et ne peuvent donc pas être exprimés sous une forme fonctionnelle universelle. Cependant, il y a tout de même une relation systématique entre la forme de la distribution des degrés de nœuds et la complexité du réseau trophique (mesurée en termes de connectance). Les réseaux trophiques à connectance élevée affichent généralement des distributions uniformes, les réseaux à connectance moyenne ont souvent des distributions exponentielles et les réseaux à faible connectance affichent des distributions de loi de puissance ou de loi de puissance partielle. La taille des réseaux trophiques joue aussi un rôle dans la forme que prend la distribution des degrés de nœuds.
Les réseaux trophiques sont plus ou moins complexes (la complexité étant le produit du nombre d'espèces et de la connectance); pour comprendre l'évolution des diverses propriétés structurales, l'utilisation de modèles est nécessaire. Sur la base de 113 réseaux trophiques tirés du monde réel, le traité Community Food Webs de Cohen, Briand, Newman (1990)[18] propose un modèle aléatoire dont les prévisions expliquent bon nombre des régularités structurales et phénomélogiques découvertes empiriquement, consolidant le modèle en cascade précédent (1985) développé sur la base des données de 62 réseaux[19]. D'autres modèles ont été avancés par la suite, à commencer par le « modèle de niche » (R.J Williams & N.D Martinez 2000) où les espèces sont rangées en fonction de leur valeur de niche : si leur valeur de niche est supérieure à celles des autres espèces alors elles se nourrissent de ces espèces. On a alors une continuité dans le régime alimentaire. De nombreux autres modèles ont été proposés au fil de la recherche afin d'optimiser au mieux la description des réseaux trophiques.
Une limite de ces modèles, même s’ils aboutissent à des descriptions satisfaisantes de la structure des réseaux trophiques, est d'utiliser des propriétés émergentes des réseaux trophiques (diversité, connectance…) comme paramètres en vue de déterminer d'autres descripteurs et de considérer que ces réseaux sont binaires (les espèces et les liens trophiques sont présents ou absents mais ne sont pas quantifiés). Les processus de niveaux inférieurs, liés à la dynamique des populations et aux phénomènes d’adaptation, ne sont pas étudiés[20].
Une autre approche, proposée par Lœuille et Loreau (2005)[20], élabore un modèle basé sur une approche simple, dans laquelle la taille corporelle est le seul trait qui prédise l’émergence de la structure des réseaux trophiques. Il apparaît alors que la structure des réseaux trophique est principalement déterminée par la largeur de niche, mais aussi par l’intensité de la compétition par interférence. Si la largeur de la niche et l'intensité de la compétition sont faibles, les espèces sont distribuées en groupe de tailles différentes correspondant à des niveaux trophiques distincts. Si elles sont fortes, la structure du réseau trophique est floue, et les espèces sont réparties de manière homogène le long de l’axe de la taille corporelle.
Taille corporelle et régime alimentaire
L'emboîtement ou « nestedness » en anglais, est un terme qui désigne un type d'organisation retrouvée dans de nombreux réseaux trophiques naturels. Il peut se définir comme un modèle d'interactions dans lequel des espèces spécialistes créent des liens trophiques avec des espèces qui forment un sous-groupe des espèces avec lesquelles des espèces plus généralistes interagissent. C'est-à-dire que les proies d'une espèce spécialiste sont un sous ensemble des proies d'une espèce plus généraliste, dont les proies sont elles aussi un sous ensemble des proies d'une espèce encore plus généraliste, d'où cette idée d'emboîtement. La taille corporelle semble jouer un rôle dans cette organisation car une espèce de petite taille doit prédater sur des proies qui appartiennent à l'ensemble des proies d'un prédateur de plus grande taille. De plus la stratégie optimale de recherche de nourriture ou « optimal foraging theory » prédit que le régime d'une espèce doit être celui qui maximise l'apport en énergie, et donc la fitness des individus. Par conséquent la taille des proies détermine l'énergie dépensée lors de la capture. Un modèle prenant en compte cette théorie et la taille corporelle a été proposé par Petchey et al. (2008)[21], ce modèle prédit assez bien la structure de différents réseaux trophiques.
Cependant, la taille peut varier de façon importante au sein d’un même groupe trophique et elle peut aussi se chevaucher entre différents niveaux trophiques, c’est le cas des herbivores et des carnivores dans les savanes africaines. Cela indique que la taille corporelle serait d’une importance moindre que d'autres facteurs pour expliquer la structure des réseaux trophiques[22].
Contraintes phylogénétiques et adaptation
Un autre modèle théorique pour expliquer la structure des interactions trophiques, en particulier cet emboitement, a été formulé par Cattin et al. (2004)[22]. Il s’agit du modèle en « hiérarchie imbriquée » (nested-hierarchy model). Ce modèle prend en compte la phylogénie qui doit contraindre le régime d’une espèce (par exemple une espèce d’oiseau peut appartenir à un genre caractérisé par un bec fait pour attraper des insectes, donc elle doit se nourrir d’insectes). Ce modèle est basé sur l'observation d'une relation significative entre le régime (les interactions trophiques) et l’origine phylogénétique. Mais il existe une variabilité des relations trophiques entre espèces qui ne s’explique pas seulement par la phylogénie. Il faut également prendre en compte l’adaptation, puisque deux espèces proches peuvent diverger s’il y a un changement de comportement ou de nourriture exploitée pour s’adapter à son environnement. Ce modèle, bien qu'il prenne en compte le concept d'adaptation, ne prend pas en compte les concepts de dynamique écologique et évolutive. Lœuille et Loreau (2005)[20] proposent un modèle décrivant bien la structure des réseaux et comment celle-ci émerge. Leur modèle qui est basé sur l'évolution d'un trait, la taille corporelle, montre que la structure des réseaux trophique est principalement déterminée par la largeur de niche et l’intensité de la compétition par interférence.
Effet de la qualité de la ressource
La qualité des ressources, c’est-à-dire des producteurs primaires à la base du réseau trophique, affecte les interactions entre espèces en contraignant la quantité d’énergie transmise entre les niveaux du réseau. D’après la théorie de l’optimal foraging, la structure des réseaux repose en partie sur la taille des prédateurs et des proies. Mais il semblerait que la variation dans les traits de certaines espèces puisse influencer la structure du réseau, notamment si les traits affectent la qualité de la ressource dans le réseau.
Cela a été mis en évidence par Bukovinszky et al. (2008)[23], qui ont étudié la structure de deux réseaux trophiques constitués des mêmes espèces trophiques (groupe fonctionnel d’espèces qui partagent les mêmes proies et les mêmes prédateurs) mais avec des producteurs primaires de qualité différente. Ils ont montré que la variation de qualité du producteur primaire a des effets en cascade sur tous les niveaux trophiques. Elle détermine en premier lieu l’abondance et la taille des herbivores qui contrôlent la diversité des liens trophiques, la densité et la diversité des consommateurs secondaires qui eux influencent les consommateurs tertiaires (le sommet de la chaine) de la même manière. Les traits du producteur primaire ont des conséquences sur la diversité et la complexité du réseau trophique, via des effets directs et indirects de la densité et de la taille des espèces dans le réseau.
On parle généralement de cascade trophique lorsqu’un prédateur affecte indirectement l’abondance des espèces des niveaux trophiques inférieurs, cas de contrôle top-down, ces espèces prédatrices sont qualifiées de clés de voute si leur impact sur l'abondance des autres espèces est très important par rapport à leur propre abondance. Mais l’exemple précédent montre que les producteurs primaires peuvent aussi avoir un contrôle indirect ascendant, contrôle bottom-up, on parle alors d’espèce « pierre fondatrice ».
Stabilité des réseaux
Le lien entre la structure des réseaux trophiques, la stabilité et le fonctionnement des écosystèmes a fait l’objet de nombreuses études. Dans le contexte de perte de biodiversité actuelle, l’impact de cette perte de biodiversité sur la stabilité des communautés est devenu un enjeu important[24]. L'effet de la complexité sur la stabilité des réseaux trophiques a souvent été pris en compte, néanmoins le lien entre complexité et stabilité n’est pas toujours évident. La complexité est définie comme le produit de la diversité spécifique et de la connectance. La diversité ou richesse spécifique de la communauté est déterminée par son passé évolutif, et par les processus d'émigration, immigration et extinction qui résultent d’interactions directes et indirectes entre les populations et au sein des populations. La connectance (ou densité de liaison), notée C dans un réseau trophique, est décrite comme le rapport de l'ensemble des liens trophiques réalisés, L, sur l'ensemble des liens trophiques possibles Lp, elle est aussi exprimée avec la diversité spécifique notée S (nombre de nœuds dans le réseau, équivaut donc aux liens possibles si exprimée au carré). On a alors : .

Connectance
Robert MacArthur formule en 1955 l'hypothèse que la stabilité de la communauté augmente lorsque le nombre de liens dans le réseau trophique augmente[25]. Cette hypothèse indique donc qu'il existerait un lien entre la connectance et la stabilité des réseaux, puisque la connectance, pour une même diversité, doit augmenter avec le nombre de liens trophiques dans le réseau.
Cette relation a été mise en évidence par Dunne et al. (2002)[26] qui se sont intéressés au rôle de la structure des réseaux trophiques dans les cascades d’extinctions engendrées par la perte de certaines espèces. À partir de réseaux trophiques naturels, ils ont modélisé des séquences d’extinctions d’espèces afin de déterminer la robustesse des réseaux.
Ils ont mis en évidence que lorsque les espèces les plus connectées sont retirées, que l’on peut qualifier d’espèces clés de voute, le nombre d’extinctions secondaires engendrées est plus important. Cela indique que ces espèces, de par leur densité de liens élevée, ont en effet un rôle important dans la stabilité de la communauté car de nombreuses espèces seront directement affectées si elles disparaissent.
La connectance a un effet important sur la robustesse des réseaux, lorsque la connectance augmente le fait de retirer des espèces clés n’a pas beaucoup d’effet jusqu’à un certain seuil à partir duquel la sensibilité à la perte d’une espèce très connectée augmente rapidement. Et ce seuil augmente avec la connectance.
Parfois, le fait de retirer les espèces les moins connectées engendre le plus d’extinctions d’espèces dans le réseau. Le fait que ces espèces aient un effet important sur la structure du réseau peut avoir différentes explications.
Il peut s’agir du cas où les espèces les moins connectées sont à la base du réseau, donc si elles sont retirées tous les niveaux supérieurs sont affectés. Une autre possibilité est qu’une espèce peut être prédatée juste par une seule espèce, mais ce prédateur a lui-même beaucoup de prédateurs donc ça engendre de nombreuses extinctions par cascade. Biologiquement ça peut s’interpréter également dans un cadre de dynamique de population avec les « weak-interactor species » qui peuvent avoir un rôle très important sur l’abondance de plusieurs espèces dans la communauté[27].
Force des interactions
Pour comprendre la stabilité du système, on utilise donc sa complexité (connectance*diversité) mais aussi la force des interactions entre les nœuds. C'est une mesure qui représente le taux de changement, par individu, causé par une espèce B sur une espèce A, ceci de manière instantanée. Elle peut aussi être considérée comme le flux de biomasse entre deux espèces (N. Rooney & K. MacCann 2012)[24]. Lorsque ces forces sont importantes dans une chaîne du réseau, on dit que cette chaîne est rapide, sinon elle est lente. Il a été montré que ces forces jouaient un rôle sur la stabilité du système. Effectivement, les faibles forces d'interactions inter-spécifiques (entre deux individus d'espèces différentes) sont considérées comme stabilisantes. À l'inverse, de grandes forces d'interactions inter-spécifiques sont plus potentiellement instables. C'est le couplage entre les forces importantes et faibles, permis par la structure du système qui aurait un effet stabilisant. Pour ce qui est des forces d'interactions intraspécifiques (entre deux individus de même espèce), elles seraient stabilisantes (P. Yodzis 1981)[28].
Ainsi pour étudier la stabilité des réseaux trophique via sa structure, les principales variables utilisées sont la force des interactions (i), la connectance (C) et la diversité (S). Robert May (1972)[29] a élaboré un critère de stabilité se basant sur ces trois mesures. Aux alentours de l'équilibre, si i*(SC)1/2<1 alors, la stabilité de la structure est assurée. Cette information a donné un élan à la recherche sur la structure des chaînes trophiques et le lien entre elle et la stabilité. Cependant, le modèle de May comportait uniquement des structures aléatoires. Yodzis (1981) [28] a réanalysé ce critère via des structures de réseaux trophiques existant. Il en a résulté que le critère de May était trop sévère car ses réseaux trophiques étaient plus stables que ceux créés aléatoirement. Ainsi, il a prouvé que l'organisation de la diversité dans le monde du vivant ne se faisait pas de manière aléatoire et que la structure du réseau était bel et bien importante.
Omnivorie
L'impact de l'omnivorie sur la stabilité des réseaux trophiques est source de débat. Selon Pimm (1979)[30] les systèmes contenant des espèces omnivores sont plus susceptibles d'être instables, le niveau d’instabilité dépendrait donc en partie du nombre d’omnivores et de leur position au sein du réseau. D'après ces résultats, la probabilité d’avoir un équilibre instable est plus élevée avec un nombre important d'omnivores, de plus les effets sur la stabilité du réseau diffèrent selon que les liens omnivores se trouvent à l’intérieur ou entre des chaînes. E K. McCann, publie en 1997 un article stipulant qu'au contraire des boucles omnivores dans un système simple et hors équilibre permettraient de réduire l'instabilité[31]. Cependant, avec une omnivorie trop importante, McCann retrouve les mêmes résultats que Pimm. Il existerait ainsi un juste milieu pour l'omnivorie permettant une stabilité optimale. Cet équilibre semble dépendre de l'efficacité du prédateur au sommet du réseau trophique, de concert avec la distribution de la force des interactions aux différents niveaux trophiques[32].
Compartimentation
Le rôle de la compartimentation dans la stabilité des réseaux a été longuement discutée, S.L Pimm en 1979[30] conclu que les compartiments au sein d'un réseau trophique font augmenter la probabilité que ce réseau soit instable. Il affirme alors que les interactions entre espèces ne doivent pas être organisées dans des compartiments étroits afin de maximiser la stabilité du système. Il n'existe dans les écosystèmes naturels que très peu de compartimentation, les écosystèmes sont donc fortement interconnectés et l’élimination de n’importe quelle espèce peut induire de grandes perturbations (J. Camacho et al 2002)[33]. Ce résultat, des modèles les plus compartimentés étant les moins stables, va à l'encontre d'un certain nombre de suggestions sur l'organisation des systèmes écologiques, les recherches ayant suivi affirment alors que théoriquement, les compartiments augmenteraient la stabilité dans les réseaux biologiques incluant les réseaux trophiques (A. Krause et al. 2003)[34]. Dans ce cas-ci, les interactions faibles entre les compartiments permettraient de cantonner une perturbation à son compartiment. L'ensemble du système serait alors plus résistant à une perturbation et donc plus stable. Ce dernier a démenti le fait que la compartimentation soit un cas isolé dans les milieux empiriques. Pour lui, le manque de compartimentation était dû à une trop faible résolution des modèles. Il a donc créé le sien mettant en évidence plus de compartimentation et obtenu ces résultats.
Rôle de l'environnement
Des recherches ont été menées sur le rôle de l'environnement sur la stabilité de la structure du réseau. Frédéric Briand (1983) a démontré le premier l'importance de la variabilité de l'environnement sur la connectance[35]. Les réseaux au sein d'environnements fluctuants (dont les ressources sont accessibles de manière restreinte dans le temps) ont une connectance moins importante que ceux des milieux constants (où la nourriture est présente constamment). Une connectance élevée est souvent associée à une force d'interactions plutôt faible pour garder le système stable avec l'augmentation de la diversité. Ce phénomène s'expliquerait par des adaptations évolutives: dans les environnements fluctuants, où la connectance est limitée par la variabilité de l'environnement, la stratégie la plus efficace sera de maximiser la force des interactions, les rendre les plus rentables possible, et ce au détriment d'une connectance de toute manière limitée. Pour les environnements constants, c'est l'inverse, miser sur la répétition des interactions peut s'avérer plus avantageux. Plus avant (1987) Frédéric Briand, associé à Joel Cohen, a découvert que la longueur moyenne et la longueur maximale de 113 réseaux trophiques étudiés augmentaient non pas en fonction de la productivité primaire mais en fonction du 'volume' de l'écosytème: elles sont ainsi plus élevées pour un écosystème pélagique ou la canopée d'une forêt que pour une prairie ou des fonds marins[36].
Conservation
Dans un contexte de conservation, les effets des facteurs précédents sur la stabilité semblent donc intéressants à prendre en compte. Les communautés avec une connectance élevée seront donc moins sensibles à des pertes d’espèces et des extinctions aléatoires auront aussi moins d’effet que des extinctions ciblées sur les espèces clés de voûte. Actuellement, il semblerait que les extinctions ne sont pas aléatoires car les activités comme la pêche ou la chasse ont plutôt visées les niveaux trophiques élevés qui contiennent souvent des espèces clés de voûte[26]. De plus, les extinctions des espèces omnivores pourraient avoir des effets importants sur la stabilité de la communauté. On voit donc l'importance pour la conservation de la biodiversité de prendre en compte le rôle des différentes espèces et la structure des réseaux trophiques.
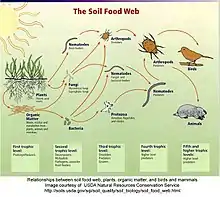
Les réseaux trophiques verts et bruns
Les « réseaux trophiques verts » désignent l’ensemble des interactions trophiques des organismes vivant au-dessus du sol. Ce réseau englobe les plantes (ou producteurs primaires) utilisant l’énergie solaire et les minéraux pour produire leur propre matière organique via la photosynthèse, les herbivores consommateurs de ces producteurs primaires jusqu’aux prédateurs (consommateurs secondaires) de ces herbivores.
On évoque le terme de « réseaux trophiques bruns » en référence aux interactions trophiques ayant lieu dans le sol. En effet, la microflore ou décomposeurs du sol (bactéries, champignons) dégrade la matière organique fraîche (minéralisation) issue des réseaux trophiques verts en surface pour produire leur propre matière organique, de l’énergie et des nutriments. Puis, ils sont à leur tour consommés par la microfaune et mésofaune ou détritivores prédateurs (nématodes, collemboles, acariens, arthropodes, protozoaires) qui complexifient la matière organique.
Ces interactions produisent finalement de l'énergie, de l’eau, du CO2 et des substances minérales telles que l’azote et le phosphore qui sont alors réintroduits dans les réseaux verts par les racines des végétaux.
Interactions entre les réseaux trophiques verts et bruns : fonctions des facteurs abiotiques et biotiques
Les réseaux trophiques verts et bruns sont reliés entre eux par le cycle de la matière organique et des nutriments. Les interactions existant entre ces deux réseaux trophiques via ces cycles déterminent le fonctionnement de l’écosystème auquel ces réseaux appartiennent.
Certains facteurs abiotiques et biotiques ont un rôle moteur dans la dynamique de ce cycle des nutriments, de la séquestration du dioxyde de carbone ainsi que de la matière organique entre ces réseaux verts et bruns.
Facteurs abiotiques
L'interaction de la nature du sol (sol dur ou souple) avec les espèces inféodées tend à déterminer la nature du réseau trophique vert ou brun. Par exemple, un substrat dur permet l’implantation d’espèces fixes ou benthiques (plantes, algues, mollusques, crustacés…) autotrophes et hétérotrophes. Riches en espèces autotrophes, ces substrats durs sont donc dominés par des réseaux verts. A contrario, les substrats souples (sous les substrats durs) tendent à être dominés par des réseaux trophiques bruns composés d’organismes hétérotrophes tels que les décomposeurs[37]. Ces facteurs abiotiques contrôlent les interactions au sein de chaque réseau trophique mais également les interactions entretenues entre ces derniers.
Par exemple, au sein des réseaux trophiques bruns, plusieurs facteurs environnementaux tels que la lumière ou encore la disponibilité en carbone et en nutriments déterminent l'activité détritique du sol. En effet, lorsque ces paramètres sont suffisamment abondants, l'assimilation des nutriments microbiens par les détritivores est favorisée. Ces paramètres influencent donc les flux d'énergie à la base des réseaux alimentaires bruns. Aussi, les contributions microbiennes aux demandes énergétiques et nutritives des détritivores diffèrent. Le carbone fongique est privilégié par les détritivores du fait de sa facilité d’assimilation comparé au carbone bactérien. Le carbone fongique constitue le plus grand contributeur à la croissance des détritivores dans les réseaux trophiques bruns[38].
Ces facteurs abiotiques sont également responsables en partie de l’interaction entre les réseaux trophiques verts et bruns. Ainsi, les effets en cascade de la chaîne alimentaire verte sur la production des décomposeurs dépendent fortement de la limitation en carbone ou en nutriments pour les décomposeurs. Par exemple, en cas de limitation en carbone, le taux de consommation des carnivores sur les herbivores des réseaux verts et le taux d’absorption des nutriments dans le sol par les producteurs primaires ont des effets positifs sur la production des décomposeurs, tandis que le taux de consommation des herbivores sur les producteurs primaires a un effet négatif sur la production des décomposeurs (retrait de matière organique potentielle future pour les décomposeurs). Si les décomposeurs sont limités seulement par le carbone, leur minéralisation des détritus profite aux producteurs primaires et mène à une interaction mutualiste entre ces décomposeurs et les producteurs primaires. Si les décomposeurs sont limités par l’azote ou par les substances nutritives, les décomposeurs sont en compétition avec les producteurs primaires pour ces ressources nutritives manquantes[39]. Les facteurs abiotiques peuvent ainsi changer les interactions entre les réseaux verts et bruns. Ces résultats permettent d’explorer les interactions et les effets en cascade entre les réseaux trophiques verts et bruns telles que les interactions pélagiques-benthiques par exemple[39].
Facteurs biotiques
Au même titre que les facteurs abiotiques, certains facteurs biotiques contrôlent les interactions au sein de chaque réseau trophique mais également les interactions entretenues entre ces derniers.
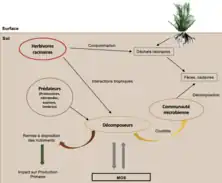
Au sein des réseaux trophiques bruns, les herbivores racinaires constituent un groupe d’animaux singulier dans les organismes du sol (nématodes, insectes, petits mammifères) car ils ont la particularité de décomposer les déchets racinaires plutôt que ceux issus des feuilles. Ils affectent la formation et la persistance de la matière organique des sols par modification de ses propriétés physicochimiques. La voie de l’herbivorie racinaire interagit avec celle des décomposeurs par des interactions directes et indirectes avec le microbiome. Les herbivores racinaires contrôlent directement la matière organique et les nutriments à proximité de la rhizosphère. La décomposition directe des racines permet d’incorporer la matière organique plus directement dans le sol afin d’être plus rapidement et facilement minéralisée par la microflore. Ces interactions autotrophes-hétérotrophes sont responsables de transferts trophiques d’énergie et de nutriments des plantes vers les réseaux alimentaires détritiques[40].
Les réseaux trophiques verts et bruns ne sont pas uniquement affectés par des facteurs biotiques propres à leur compartiment. La « boucle microbienne » désigne le processus tel que les prédateurs des réseaux trophiques bruns (protozoaires, nématodes, acariens, lombrics) peuvent accroître la minéralisation des nutriments dans les écosystèmes aquatiques et terrestres en consommant les décomposeurs, ce qui peut avoir une incidence indirecte sur la production primaire par une remise à disposition des nutriments[39]. Ainsi, les producteurs primaires sont affectés par les décomposeurs et leurs prédateurs.
À l’inverse, les prédateurs des chaînes trophiques vertes ont un rôle déterminant sur la dynamique des réseaux trophiques verts, mais il est également important de considérer leur influence sur les processus occurrents dans les réseaux trophiques bruns du sol. La nutrition “multicanale” fait référence à la consommation de proies autant sur les réseaux trophiques verts que les réseaux trophiques bruns (un prédateur peut à la fois consommer des herbivores et des organismes du sol). À cette nutrition « multicanale » peut être associée une « peur multicanale ». Autrement dit, les prédateurs multicanaux peuvent générer des répercussions à la fois sur les chaînes alimentaires herbivores ou détritivores.
De manière générale, les prédateurs affectent les niveaux trophiques inférieurs par deux mécanismes distincts: la consommation des proies ou le changement de physiologie et de comportement des proies en réponse à la prédation. Les effets non liés à la consommation correspondent à la modification du comportement de quête de nourriture chez des détritivores ou herbivores. Sous l’effet du risque de prédation, ces derniers peuvent diminuer leur activité de nutrition. Ces changements comportementaux de la part des détritivores ont un impact direct sur le taux de décomposition de la matière organique et la dynamique des nutriments du sol (impact sur le réseau brun). Des modifications peuvent aussi être observées sur la physiologie et la composition de la communauté chez les plantes en réponse à la pression de prédation (impact sur le réseau vert). Ce sont des conséquences indirectes de la pression prédatrice verte sur les deux compartiments verts et bruns[41].
Stabilité
Le couplage des réseaux trophiques verts et bruns joue un rôle crucial dans la stabilité de l’écosystème dans lequel ces réseaux évoluent. Stabiliser un écosystème signifie recycler les matières minérales et les rendre de nouveau disponible pour les autres végétaux. Il existe divers arguments en faveur de chacun des réseaux trophiques verts ou bruns pour défendre la stabilité supérieure de l’un par rapport à l’autre, cette stabilité d’un réseau trophique étant observable après une perturbation.
Tout d’abord, les réseaux trophiques bruns semblent être plus stables que les réseaux trophiques verts car la décomposition des débris organiques n’a pas de dynamique propre (pas de reproduction ni de croissance). Les réseaux trophiques bruns semblent donc être dominés par la dynamique bottom-up (la matière organique conditionne une partie de la dynamique des décomposeurs)[37]. Les décomposeurs présents dans le sol ont un rôle stabilisateur particulièrement important car ils permettent le recyclage des matières minérales, les rendant de nouveau disponibles pour les végétaux composants les réseaux trophiques verts : ils assurent donc la bonne circulation des nutriments au niveau du sol. Un taux de décomposition modéré tend à stabiliser l’écosystème environnant permettant d’augmenter sa résilience, c’est-à-dire le retour rapide à son état de départ. De plus, le degré d’ouverture de l’écosystème dans lequel se trouvent ces réseaux trophiques a un impact sur la stabilité de ces derniers. Les écosystèmes présentant un degré intermédiaire d’ouverture, autrement dit les écosystèmes semi-perméables aux échanges d’énergie et de matière avec les autres réservoirs tels que l’atmosphère, sont hautement stables[42]. La stabilité des réseaux bruns est accentuée par la présence de prédateurs omnivores bruns capables de se nourrir d’autres espèces en cas de perturbations. En dépendant d’autres ressources, ils permettent à leurs proies de prédilections affectées de croître de nouveau[37].
De plus, si l’on considère d’autres proxies, les réseaux bruns semblent encore se rétablir rapidement après une perturbation locale. Pour les substrats souples, les mesures de connectance, de la moyenne des niveaux trophiques et du clustering sont supérieurs à ceux des réseaux trophiques généraux. Néanmoins, pour les substrats durs, la connectance, le clustering et la moyenne des réseaux trophiques sont inférieurs à ceux dans les réseaux trophiques généraux.
Il faut donc prendre en compte tous les proxies et la complexité des réseaux trophiques (type habitat, degré d'ouverture, connectance, clustering, taux d’omnivores, taux de décomposition de la matière organique) pour évaluer leur stabilité.
Impacts anthropiques - restauration des écosystèmes par interactions trophiques
Le changement climatique et les activités anthropiques actuelles ont un impact significatif sur les interactions et la stabilité des réseaux trophiques. Chaque niveau trophique d’un écosystème peut répondre différemment au changement climatique, indépendamment de son appartenance aux réseaux trophiques verts ou bruns. Cette réaction hétérogène peut être à l’origine de déséquilibres trophiques entre organismes, dont les conséquences sur les communautés et les écosystèmes (flux de masse et d’énergie) sont encore peu connues[43].
Plus précisément, un déséquilibre trophique se caractérise par un changement de phénologie chez un organisme ressource sans que ce soit le cas pour son organisme consommateur. À titre d’exemple, la période de reproduction et le pic d’abondance d’une plante peuvent être modifiés sous l’effet des pressions climatiques, tandis qu’aucune modification n’est observée quant à l’attitude consommatrice de l’organisme herbivore envers cette plante. Le raisonnement inverse peut aussi exister. Si un prédateur de surface, tel qu’une araignée, change sa période d’incubation mais que sa proie, un collembole, n’est pas soumise aux effets du changement climatique, un déséquilibre trophique est alors un scénario probable.
Ainsi, ces déséquilibres trophiques dus au réchauffement climatique sont susceptibles d'éloigner les réseaux trophiques d’un état de stabilité en modifiant les forces d’interaction entre ressources et consommateurs. Plus particulièrement, les réseaux trophiques verts seraient plus susceptibles d’être exposés à des décalages trophiques que les réseaux trophiques bruns. C’est ce qu’on appelle « l’hypothèse du déséquilibre brun-vert ». Effectivement, les réseaux verts offrent des habitats favorables à l’existence de fortes interactions trophiques spécialisées entre organismes (relations hôtes parasites ou encore herbivores spécialistes) contrairement aux réseaux bruns plus généralistes. De plus, la tolérance thermique des microorganismes du sol (ressource basale des réseaux bruns) étant plus accrue, leurs consommateurs sont moins sujets à être sous-alimentés à la suite de l’augmentation des températures du sol. Cette plus faible sensibilité des microorganismes du réseau brun face aux changements de température permettrait de minimiser les déséquilibres trophiques avec leurs consommateurs, rendant ce milieu en moyenne plus stable que le réseau vert.
Ainsi, les flux de masses et d’énergie entre les deux compartiments sont indispensables pour le maintien du bon fonctionnement de l’écosystème, qu’il s’agisse de la séquestration du carbone, de la production primaire ou de la minéralisation des nutriments. Une perturbation de ces flux au travers du déséquilibre brun-vert peut participer au dérèglement des processus piliers d’un environnement donné. Or, ces interactions écologiques entre les deux réseaux permettent des réponses adaptatives participant à leur stabilité après de tels impacts. Ces interactions permettent aussi la restauration des écosystèmes perturbés. Pour cela, il faut considérer trois niveaux d’interactions : les interactions interspécifiques (cibler les actions sur des espèces précises ou sur des intéractions comme le mutualisme et la symbiose), les interactions entre les communautés des réseaux verts et bruns et les interactions concernant les fonctions dans l’écosystème.
Concernant les interactions interspécifiques des réseaux bruns, il est démontré que la microflore (bactéries et champignons) et les micro-mésofaunes (acariens, nématodes, lombrics, araignées, collemboles) ayant des actions directes sur les plantes seraient plus favorables pour restaurer les sols. Par exemple, pour les pathogènes, il est postulé qu’introduire certains pathogènes issus de la terre native de plantes envahissantes pourrait mieux contrôler et inhiber le développement de ces plantes envahissantes dans l’habitat qu’elles colonisent. Les pathogènes de l’habitat envahit seront moins efficaces sur l’espèce envahissante que les pathogènes inféodés à l’habitat natif de la plante[44]. Par exemple, les herbivores racinaires utilisent trois voies pour favoriser la séquestration du dioxyde de carbone dans le sol. Premièrement, la voie simple de consommation des racines et de transformation en déchets par les fèces apporte une grande quantité de carbone à la microflore. Deuxièmement, la voie d’altération physique des racines par les insectes et d’une infiltration des exsudats nutritifs dans le sol permet de stocker du carbone. Finalement, les herbivores racinaires interagissent avec les autres organismes du sol, acteurs clé de la médiation du flux de carbone souterrain[41].
Entre les communautés, les facteurs biotiques sont cruciaux pour comprendre la vitesse de restauration des plantes. Par exemple, il existe une corrélation positive entre l’abondance et la nature de l'interaction plante-mycorhizes avec la richesse spécifique des plantes au niveau supérieur. Cependant, les feedbacks négatifs entre plantes-pathogènes du sol conduisent à des gazons ou à des prairies uniformes (moins de séquestration du dioxyde de carbone, moins de ressources pour les herbivores et donc moins de richesse spécifique). Les consommateurs primaires (herbivores du réseau vert) jouent également un rôle dans la restauration et le maintien des végétaux. Par exemple, la réapparition des troupeaux de gnous après la peste bovine en 1960 a rétabli la stabilité des végétaux. Cela a favorisé l’essor des organismes du sol capables de reminéraliser la matière organique au sein d’un cercle vertueux pour les plantes. À terme, les plantes ont de nouveau pu séquestrer le dioxyde de carbone. Il existe donc un double contrôle entre les réseaux bruns et verts[44].
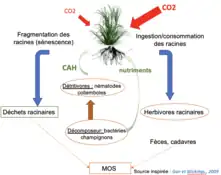
Enfin, la majorité des fonctions de l'écosystème telles que la séquestration du dioxyde de carbone sont rendues possibles par les différents niveaux trophiques. Par exemple, les forêts séquestrent beaucoup plus de carbone que les prairies ou les cultures. Il conviendrait donc de favoriser l’implantation de forêts en croissance qui sont les plus efficaces pour la séquestration du dioxyde de carbone. La modification de l’utilisation des sols perturbe également les communautés du sol.
Il est donc indispensable de prendre en compte les liens interactifs entre les communautés supérieures et les communautés du sol afin de comprendre la stabilité et la réponse des réseaux trophiques bruns et verts à une perturbation, d’autant plus que les fonctions écosystémiques du sol dépendent directement de l’abondance et de la qualité des organismes bruns. Par exemple, dans certaines expériences de réhabilitation des sols, l’activité des organismes du sol contribue à la diversité des plantes[44].
Réseau limité
D'après la loi de Raymond Lindeman (1942)[45], la quantité d'énergie passant d'un maillon à l'autre de la chaîne est de seulement 10 %. Ceci implique que les chaînes trophiques sont limitées ; par ailleurs, dans les échelons les plus bas de la chaîne, l'énergie est allouée en grande quantité à la reproduction. En bout de chaîne en revanche, les espèces ont une reproduction plus limitée et l'énergie est allouée à la survie (chasse, défense, etc.).
Certaines substances toxiques, « indésirables » ou polluantes sont bioaccumulées dans la chaîne alimentaire[46]. En contexte pollué notamment, elles constituent aussi une limite probable au nombre de maillons (niveaux trophiques) de la chaîne.
Contrôle dans les réseaux trophiques
On qualifie de « contrôle » dans un réseau trophique l’influence de certains paramètres sur la structure et la dynamique de celui-ci. On regroupe sous ce terme de “paramètres” les interactions ou les niveaux trophiques jouant sur la distribution et l’abondance des organismes. Si la nature de ces contrôles fait plutôt consensus dans la communauté scientifique, à savoir le contrôle top-down (contrôle par les prédateurs) et le contrôle bottom-up (contrôle par la compétition), la part de chacun dans les réseaux reste encore débattue[47].
Contrôle bottom-up
Le contrôle bottom-up correspond à une régulation de la biomasse d'un réseau trophique, uniquement ou principalement, induite par la compétition entre individus d'un même niveau trophique pour l'accès aux ressources. On observe ainsi une covariation positive de la biomasse entre niveaux trophiques adjacents. Dans la littérature scientifique, deux visions de ce concept de régulation transparaissent, en s'opposant sur certains point[47].
La vision de White[48] où le contrôle bottom-up est présenté comme la force de régulation globale de tous les écosystèmes. White part du postulat suivant : le monde est un endroit inhospitalier où la survie d'un individu dépend plus de son accès à la nourriture que de la prédation ou l'herbivorie. Ainsi, la principale ressource limitante des écosystèmes est la nourriture et, par extension, la quantité de nutriments présents dans le sol (comme l'azote), car étant un des facteurs principaux influant sur la production primaire, elle-même impactant les herbivores, et ainsi de suite jusqu'au sommet du réseau trophique.
Ce postulat induit par la même occasion un paradoxe vis-à-vis de la théorie du Monde Vert : un contrôle induit par la quantité de ressources n'expliquerait pas la forte proportion de végétaux et de prédateurs par rapport à celle des herbivores sur Terre. Ce paradoxe est désamorcé par White, qui avance l'hypothèse qu'une faible proportion de la végétation peut réellement être considérée comme comestible (et donc en tant que « ressources ») du fait de la faible qualité nutritive de la majorité des végétaux (induite par de trop faibles concentrations d'azote dans les sols). Cette vision s'inspire des travaux de Chitty (1960)[49] et de Wynne-Edwards (1962) qui proposaient que la limitation du nombre d'individus trouve son origine dans des mécanismes intrinsèquement comportementaux ou génétiques d'autorégulation, comme la territorialité permettant la préservation d'une ressource limitée par une limitation de la reproduction (et donc un maintien de la population à son plus haut niveau). Ce concept de populations « limitées par le bas » plutôt que « contrôlées par le haut » conduit à une réévaluation du rôle de la prédation, du comportement social et territorial ou tout autres interactions entre niveaux trophiques, en tant que facteurs susceptibles d'influencer l'abondance des populations dans un environnement.
La vision de Hunter & Price[50] où le contrôle bottom-up est présenté comme une des forces de régulation des réseaux trophiques, sans exclusivité, et soumise à l'hétérogénéité de l'environnement et à la diversité des écosystèmes.
Plusieurs facteurs abiotiques (climat, ressource en eau, nutriments…) et biotiques (prédation/parasitisme, symbiotes, compétiteurs, décomposeurs…) entre alors en jeu et les rôles relatifs de chacun quant à la structure des communautés sont propres à chaque écosystème.
_%E2%80%93_interpr%C3%A9tation_de_l%E2%80%99article_de_Hunter_%2526_Price_(1992).jpg.webp)
Hunter & Price proposent un modèle théorique « bottom-up dominé » (voir figure ci-contre), s'appuyant sur l'observation que la suppression des niveaux trophiques les plus élevés laisse les niveaux inférieurs présents (mais possiblement fortement modifiés), alors que la suppression des producteurs primaires ne laisse aucun système. Dans ce modèle, les facteurs déterminants la production végétale sont les facteurs abiotiques (climat, paramètres du sol…) et les facteurs biotiques (décomposeurs, symbiotes…) de l’environnement, puisqu’ils afféctent directement l'hétérogénéité de la production végétales. La production primaire détermine ainsi la distribution dans l'espace et le temps des populations d’herbivores, ainsi que leur qualité intrinsèque en tant que ressources pour leurs prédateurs. Cette hétérogénéité parmi les herbivores détermine à son tour la densité, la diversité des espèces et la distribution du niveau supérieur, et ainsi de suite jusqu’au sommet. De plus, ce modèle prend en compte des facteurs plus complexes, comme les interactions entre espèces et les effets de l'hétérogénéité abiotique, et ce à tous les niveaux trophiques.
Ce modèle théorique est présenté par Hunter & Price comme une première piste à suivre pour la compréhension de systèmes biologiques. Il a en effet pour avantages d’intégrer les boucles de rétroaction biotique/abiotiques ainsi que de prendre en compte l'hétérogénéité des écosystèmes. Malgré tout, ce modèle demande à être complexifié car ne prenant pas en compte les processus évolutifs, ni la diversité de facteurs biotiques et abiotiques, ni les interactions biologiques.
Top-down
Le concept de contrôle top-down (contrôle du sommet vers la base) stipule que la prédation prévaut sur la compétition dans les réseaux trophiques, et donc que la quantité et l’efficacité des prédateurs détermine les effectifs des populations de niveaux trophiques inférieurs[51]. Ce concept n’exclut pas l’impact des ressources sur les organismes de l’écosystème : un milieu plus riche pourra toujours contenir de plus grands effectifs de plantes, d’herbivores et donc de prédateurs. Cependant, le contrôle top-down indique un rétrocontrôle négatif (direct) ou positif (indirect) des effectifs des prédateurs sur les densités des niveaux trophiques inférieurs.
Ce concept découle de l’« Hypothèse du Monde Vert » de Hairston et ses collaborateurs[51] qui présente le fait que bien qu’en général les plantes soient présentes en grand nombre dans tous les environnements terrestres, il existe cependant des cas où les herbivores sont capables d’épuiser complètement leur ressource et de localement provoquer la disparition des plantes (herbivores protégés par l’humain ou par des phénomènes naturels, espèces introduites, etc.). Cette observation peut être expliquée non pas par le fait que les espèces introduites soient « mieux adaptés » à leur nouvel environnement et capables d’exploiter mieux les ressources que les espèces natives, mais plutôt que leur capacité à exploiter le milieu est simplement dû à l’absence de leur prédateur naturel. Ce sont les prédateurs qui contrôlent ainsi les effectifs des herbivores, et non pas la quantité de ressource à leur disposition.
La validation expérimentale de ce concept fait suite aux expériences des années 1960 afin de comprendre pourquoi et comment la biodiversité est répartie sur le globe[52] - [53] - [54]. L’expérience de Robert T. Paine (1966)[55] portant sur la complexité des réseaux trophiques et leur lien avec la diversité spécifique par de l’hypothèse de travail que « La diversité locale est directement liée à l’efficacité des prédateurs à empêcher la monopolisation de la grande partie des ressources nécessaires par une espèce ». Pour la tester il réalise une expérience d’un an dans laquelle il retire le prédateur au sommet du réseau trophique de sa zone d’étude, l’étoile de mer Pisaster ochraceus dans la zone intertidale, et regarde l’impact qu’a sa disparition sur la diversité spécifique du milieu. En un an, le réseau trophique passe de 15 espèces à 8 espèces : Paine parle de système « trophiquement simplifié ». Ainsi en l’absence d’un facteur de régulation (ici la prédation), il y a un « gagnant » dans la compétition interspécifique pour l’espace, et le système tend vers la simplicité. L’absence de contrôle par les prédateurs induit ainsi l’exclusion compétitive d’espèces de niveaux trophiques inférieurs. Cette étude illustre l’importance du contrôle top-down dans un réseau trophique et la relation directe entre diversité spécifique et quantité et efficacité de prédateurs dans le système afin d’empêcher une seule espèce à monopoliser la ressource limitante du milieu.
Dans les réseaux comportant un seul prédateur au sommet du réseau, celui-ci est donc le seul à réguler toute la biomasse du réseau : on peut alors parler d’espèce clef de voûte de l’écosystème, sa disparition entraîne une suite de conséquences dans l’écosystème appelé cascade trophique. La réintroduction du loup Canis lupus dans le parc National de Yellowstone est une illustration typique de ce contrôle top-down : sa réintroduction dans les années 1990 a induit une réduction par trois de la population des wapitis Cervus canadensis et une forte augmentation du couvert végétal.
Mécanismes de modulation
Les principales forces Bottom-up et Top-down contrôlant les interactions dans les réseaux trophiques peuvent être modulées par des mécanismes influençant les interactions entre proies et prédateurs[47].
Hétérogénéité de l’habitat
En effet, les proies peuvent avoir recours à des changements de localisation[56] leur permettant d’échapper aux prédateurs, comme l’utilisation de refuges spatiaux[57]. L’hétérogénéité structurale de l’environnement peut ainsi procurer aux proies des refuges où le risque de prédation est réduit. Ces refuges peuvent être permanents (absence complète des prédateurs) ou transitoires (prédateurs rarement présents). Une augmentation de la richesse spécifique est observée dans les environnements plus structurés puisque le taux de recherche de proies par les prédateurs est réduit, ainsi ces refuges améliorent la persistance des proies en réduisant l'efficacité des prédateurs.
Ces changements de localisation des proies peuvent être accompagnés de changements de comportements de celles-ci afin d’être plus difficiles à capturer. Par exemple, la proie peut diminuer son temps d’alimentation en diminuant son temps de recherche de nourriture puis en manipulant celle-ci dans des lieux sûrs afin d’éviter de se faire attraper par un prédateur. Cependant cela n’est possible que pour des espèces pour lesquels l’apport de nourriture pour le développement et la croissance n’est pas nécessaire en continu (discontinuouss feeders)[56]. En effet dans le cas des « continuous feeders », cette perte de temps d’alimentation peut engendrer un allongement du temps de développement et donc un risque d’être prédaté avant l’âge de reproduction.
Stratégies comportementales
Les proies peuvent également privilégier certains comportements de groupe afin d’échapper au prédateur, comme les cigales qui émergent toutes en même temps tous les 13 ans : leur grand nombre submerge les oiseaux prédateurs et les contraint à ne se focaliser que sur un nombre restreint de proies.
D’autres changements de comportements sont observables, comme l’induction de défenses chimiques ou morphologiques, des stratégies anti-prédateurs telles que le camouflage, ou la présence d’un état de vigilance qui augmente avec le risque de prédation, comme chez les mésanges bleues lors de leur recherche de nourriture[58], ou encore la coloration aposématique et le dégoût associé, comme chez les espèces de papillons qui présentent sur leurs ailes des couleurs d'avertissement qui signalent aux prédateurs un danger à les prédater[59].
Stratégies physiologiques
Lorsque les proies sont des plantes et les prédateurs des herbivores, les défenses anti-prédateurs peuvent être directes comme les épines, les cires ou encore des composés volatils interagissant directement avec le prédateur de la plante en perturbant sa croissance, ou en limitant voire inhibant sa capacité à s’alimenter. Les défenses peuvent également être indirectes et n’ont donc pas d’influence directe sur les herbivores mais bien sur les prédateurs de ces derniers, impactant donc des niveaux trophiques supérieurs. Ce type de défenses se fait grâce à des métabolites secondaires comme les HIVOC (Herbivore-Induced Volatiles Organic Compounds) ou l’EFN (Extra-Floral Nectar).
La réponse non linéaire de la productivité primaire au pâturage
À de faibles pressions de pâturage, les pertes des plantes peuvent être compensées par une stratégie de croissance rapide, stimulée par le recyclage des nutriments. À mesure que le pâturage augmente, les pertes de biomasse pour les végétaux atteindront des niveaux qui ne peuvent être compensés par une croissance stimulée, et la productivité primaire spécifique à la zone diminuera. Des relations hautement non linéaires entre la pression de pâturage et le renouvèlement des plantes pourraient ainsi dissocier et déstabiliser l’équilibre du contrôle top-down dans ces réseaux, rendant difficiles ou impossibles à prédire les rendements de cultures sur pied sous différents régimes trophiques[47].
Dans les sols, le recyclage des nutriments entre la matière organique morte, les microorganismes décomposeurs, leurs consommateurs et les producteurs primaires fournit un bon exemple de la façon dont les forces bottom-up et top-down sont importantes et étroitement liées dans les réseaux trophiques, dont les effets sont visibles à des échelles spatiotemporelles suffisamment larges. Il a été démontré à maintes reprises que le broutage des microorganismes par la faune améliore la minéralisation des nutriments minéraux (N, P, S, etc.) dans le sol, en particulier l'azote (N) et le phosphore (P). La disponibilité accrue de ces éléments augmente avec la complexité de la chaîne alimentaire, ce qui se traduit par une amélioration de la croissance des plantes et une augmentation de la concentration de ces éléments dans la biomasse végétale. Ainsi, les « nourrisseurs microbiens » comme les prédateurs peuvent exercer un contrôle top-down sur les microbes par des interactions d'alimentation, et également affecter indirectement la croissance microbienne en influençant la quantité et la qualité de la litière végétale, la ressource nécessaire aux communautés microbiennes. Cet exemple montre que des rétroactions positives, provoquées par des interactions indirectes entre les niveaux trophiques non adjacents dans les réseaux trophiques du sol, peuvent exercer un certain contrôle sur la structure et la fonction des réseaux trophiques détritiques[60].
Effet de la couverture fournie par les plantes
L’intensité de prédation des proies par les prédateurs change également avec l’importance de la couverture végétale : les plantes, en s'accumulant, fournissent une couverture aux organismes en limitant leur détection par les prédateurs[47]. Ainsi la disponibilité en ressource pour les plantes et l’accroissement du couvert végétal (force bottom-up) peut avoir pour conséquence de tamponner la réduction des effectifs d’herbivores par les prédateurs (force top-down).
Régulation intrinsèque et théorie de Fretwell
Il a été également démontré l’importance des mécanismes de régulations intrinsèques de certains niveaux trophiques dans le contrôle général des réseaux. En effet, le phénomène de cascade trophique illustre l’importance des apex prédateurs dans le but de limiter les tailles de populations de divers taxons (contrôle top-down), contrôlant ainsi la densité de leur proie et celle des mésoprédateurs. La question se posant alors est celle de comment les apex prédateurs sont eux-mêmes contrôlés : la présence d’une auto-régulation au sein de ces prédateurs en serait la réponse. L’étude a donc été menée chez ces prédateurs au sommet de la chaine trophique afin de savoir quels traits d’histoires de vie leur permettait de ne pas surexploiter leurs ressources. De ce fait, la disponibilité en proies et par conséquent la biodiversité sont maintenues[61].
Un autre exemple de régulation intrinsèque serait la théorie du professeur Fretwell. En effet, il applique cette théorie afin d’expliquer les multiples interactions trophiques d’un écosystème qui rendrait un paysage plus « vert » ou non, en fonction de la parité du nombre de niveaux trophiques[47]. Selon lui, si les chaînes alimentaires ont des nombres impairs, les brouteurs seraient limités aux prédateurs et les paysages devraient être remplis d'une végétation luxuriante. Alors que les chaînes alimentaires ont un nombre de niveaux trophiques pair, les plantes seraient limitées par les brouteurs et les paysages devraient apparaître stériles. D’après Fretwell, ce serait l’augmentation de productivité primaire qui ajouterait des niveaux trophiques de sorte que, les paysages apparaissent alternativement verts ou stériles.
Histoire du concept
Les chaînes alimentaires sont connues des humains depuis bien longtemps, et ont notamment été étudiées en détail par le philosophe grec Aristote :
« Toutes les fois que les animaux habitent les mêmes lieux et qu'ils tirent leur vie des mêmes substances, ils se font mutuellement la guerre. Si la nourriture est par trop rare, les bêtes, même de race semblable, se battent entre elles. […] Tous les animaux sont en guerre avec les carnivores, qui mutuellement sont, eux aussi, en guerre avec tous les autres, puisqu'ils ne peuvent vivre que d'animaux. […] L'aigle et le serpent sont ennemis, parce que l'aigle se nourrit de serpents. »
— Aristote, Histoire des animaux (v. 343 av. J.-C.), VI, 2.
Les réseaux trophiques ont été théorisés dès 1927 par le biologiste Charles Elton (Animal ecology). Certaines espèces, dites espèces clé de voûte, ont un rôle important au sein des écosystèmes, par leur influence sur plusieurs autres espèces pouvant être réparties dans différents « niveaux trophiques ».
Depuis les années 1970, les écologues tentent de mieux comprendre les réseaux trophiques.
On a d'abord cru que le degré de séparation entre 2 espèces d'un réseau trophique était d'au moins 4 liens, c'est-à-dire 4 relations de type « prédateur-proie », ce qui laissait penser que la disparition d'une espèce pouvait facilement être compensée par l'occupation de sa niche écologique par une autre. Cependant, l'observation des réseaux trophiques naturels a montré[62] que plus de 95 % des espèces d'un habitat naturel (aquatique ou terrestre) étaient liées à un réseau trophique dépendant de l'habitat en question, et à moins de trois liens les unes des autres (deux liens en moyenne).
De plus des interactions durables de type symbiose/parasitisme complexifient ces réseaux en rendant les espèces plus dépendantes les unes des autres que ce que l'on pensait, d'où les inquiétudes pour la régression accélérée de la biodiversité causée par les activités humaines.
Cette proximité des espèces dans le réseau trophique (deux degrés de séparation) signifie que l'extinction d'une espèce peut avoir des impacts en cascade plus importants que ce qu'on pensait antérieurement. Même des espèces autrefois jugées banales et ubiquistes, parce que communes, pourraient être concernées par les impacts indirects d'un recul « en réseau » de la biodiversité.
Dans les milieux aquatiques, marin notamment, est peu à peu apparu que l'importance des réseaux trophiques microbiens a été sous-estimée[63]. Les interactions fungi-fungi ou fungi-bactéries jouent en particulier un rôle important, voire critique dans les processus de décomposition subaquatique de la matière organique, et donc dans le cycle du carbone. Ces réseaux trophiques microbiens, partout présents, mais souvent invisibles à nos yeux, commencent à être étudiés, avec une approche se voulant plus holistique[64] grâce notamment au barcoding moléculaire.
Structuration de la recherche
L'écologie trophique a bénéficié du développement de l'écologie isotopique et de l'analyse génétique. Elle se développe en France notamment via le réseau pluridisciplinaire de chercheurs GRET (Recherche en écologie trophique), mis en place en France en 2012 puis labellisé GDR 3716[65] par le CNRS début 2015. Ce réseau vise à mieux décrire et comprendre les réseaux trophiques en France et leur rôle pour les écosystèmes, biocénoses et services écoystémiques.
Notes et références
- (en) E.A. Canuel, A.C. Spivak, E.J. Waterson et J. Emmett Duffy, « Biodiversity and food web structure influence short-term accumulation of sediment organic matter in an experimental seagrass system », Limnol. Ocean., vol. 52, , p. 590-602 (lire en ligne [PDF]).
- (en) A.R. Ives et B.J. Cardinale, « Food–web interactions govern the resistance of communities after non-random extinctions », Nature, vol. 429, , p. 174–177.
- (en) M. Emmerson, M. Bezemer, M.D. Hunter, T.H. Jones, « Global change alters the stability of food webs », Global Change Biology, vol. 11, , p. 490–501.
- (en) O.L. Petchey, P.T. McPhearson, T.M. Casey et P.J. Morin, « Environmental warming alters food-web structure and ecosystem function », Nature, vol. 402, , p. 69-72 (lire en ligne [PDF]).
- (en) E. Sanford, « Regulation of keystone predation by small changes in ocean temperature », Science, vol. 283, , p. 2095–2097 (lire en ligne [PDF]).
- (en) D.E. Burkepile et M.E. Hay, « Herbivore Vs Nutrient control of marine primary producers », Ecology, vol. 87, no 12, , p. 3128-3139 (DOI 10.1890/0012-9658(2006)87[3128:HVNCOM]2.0.CO;2).
- (en) James E. Cloern, « Our evolving conceptual model of the coastal eutrophication problem », Marine Ecology Progress series, vol. 210, , p. 223–253 (lire en ligne [PDF]).
- (en) N.F. Caraco, J.J. Cole et D.L. Strayer, « Top down control from the bottom: Regulation of eutrophication in a large river by benthic grazing », Limnol. Oceanogr., vol. 51, , p. 664-670 (lire en ligne [PDF]).
- (en) D.L. Strayer, M.L. Pace, N.F. Caraco, J.J. Cole et S.E.G. Findley, « Hydrology and grazing jointly control a large-river food web », Ecology, vol. 89, no 1, , p. 12-18 (lire en ligne [PDF]).
- Caraco, N.F. 2006. Water Chestnut impacts on oxygen in the tidal Hudson River. Aquatic Invaders 17:8-9.
- Fernald, S.H., N.F. Caraco, and J.J. Cole. 2007.Hanges in cyanobacterial dominance following the invasion of the zebra mussel Dreissena polymorpha: Long-term results from the Hudson River estuary. Estuaries and Coasts 30(1):163-170.
- Peters RH (1983) The Ecological Implications of Body Size. Cambridge University Press, Cambridge.
- Andrea E. Alpine et James E. Cloern ; Trophic interactions and direct physical effects control phytoplankton biomass and production in an estuary ; Limnol. Oceanogr., 37(S), 1992, 946-955 0 1992, by the American Society of Limnology and Oceanography, Inc.
- Leonardos et Lucas, 2000 ; The nutritional value of algae grown under different culture conditions for Mytilus edulis L. larvae ; Aquaculture (Ed : Elservier), Volume 182, Number 3, 15 février 2000, pp. 301-315(15) (Résumé).
- « Chaîne alimentaire : l'homme, un anchois comme les autres », sur liberation.fr, (consulté le ).
- (en) Sylvain Bonhommeau, Laurent Dubroca, Olivier Le Pape, Julien Barde, David M. Kaplan, Emmanuel Chassot et Anne-Elise Nieblas, « Eating up the world’s food web and the human trophic level », PNAS, vol. 110, no 51, , p. 20617-20620 (DOI 10.1073/pnas.1305827110).
- (en) Jennifer A. Dunne, Richard J. Williams et Neo D. Martinez, « Food-web structure and network theory: The role of connectance and size », Proceedings of the National Academy of Sciences, vol. 99, no 20, , p. 12917–12922 (ISSN 0027-8424 et 1091-6490, PMID 12235364, PMCID PMC130560, DOI 10.1073/pnas.192407699, lire en ligne, consulté le ).
- (en) J.E. Cohen, F. Briand et C.M. Newman, Community Food Webs: Data and Theory, Berlin, Heidelberg, New York, Springer, (ISBN 9783642837869, lire en ligne), p. 308
- (en) Joel E. Cohen, C. M. Newman et F. Briand, « A stochastic theory of community food webs II. Individual webs », Proceedings of the Royal Society of London. Series B. Biological Sciences, vol. 224, no 1237, , p. 449–461 (lire en ligne).
- (en) Nicolas Lœuille et Michel Loreau, « Evolutionary emergence of size-structured food webs », Proceedings of the National Academy of Sciences of the United States of America, vol. 102, no 16, , p. 5761–5766 (ISSN 0027-8424, PMID 15824324, DOI 10.1073/pnas.0408424102, lire en ligne, consulté le ).
- (en) O. L. Petchey, A. P. Beckerman, J. O. Riede et P. H. Warren, « Size, foraging, and food web structure », Proceedings of the National Academy of Sciences, vol. 105, no 11, , p. 4191–4196 (ISSN 0027-8424 et 1091-6490, PMID 18337512, PMCID PMC2393804, DOI 10.1073/pnas.0710672105, lire en ligne, consulté le ).
- (en) Marie-France Cattin, Louis-Félix Bersier, Carolin Banašek-Richter et Richard Baltensperger, « Phylogenetic constraints and adaptation explain food-web structure », Nature, vol. 427, no 6977, , p. 835–839 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature02327, lire en ligne, consulté le ).
- (en) T. Bukovinszky, F. J. F. van Veen, Y. Jongema et M. Dicke, « Direct and Indirect Effects of Resource Quality on Food Web Structure », Science, vol. 319, no 5864, , p. 804–807 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1148310, lire en ligne, consulté le ).
- (en) Neil Rooney et Kevin S. McCann, « Integrating food web diversity, structure and stability », Trends in Ecology & Evolution, vol. 27, no 1, , p. 40–46 (DOI 10.1016/j.tree.2011.09.001, lire en ligne, consulté le ).
- (en) Robert MacArthur, « Fluctuations of Animal Populations and a Measure of Community Stability », Ecology, vol. 36, no 3, , p. 533 (ISSN 0012-9658, DOI 10.2307/1929601, lire en ligne, consulté le ).
- (en) Jennifer A. Dunne, Richard J. Williams et Neo D. Martinez, « Network structure and biodiversity loss in food webs: robustness increases with connectance », Ecology Letters, vol. 5, no 4, , p. 558–567 (ISSN 1461-023X et 1461-0248, DOI 10.1046/j.1461-0248.2002.00354.x, lire en ligne, consulté le ).
- (en) E. L. Berlow, « Strong effects of weak interactions in ecological communities », Nature, vol. 398, no 6725, , p. 330–334 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/18672, lire en ligne, consulté le ).
- (en) P. Yodzis, « The stability of real ecosystems », Nature, vol. 289, no 5799, , p. 674–676 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/289674a0, lire en ligne, consulté le )
- (en) ROBERT M. MAY, « Will a Large Complex System be Stable? », Nature, vol. 238, no 5364, , p. 413–414 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/238413a0, lire en ligne, consulté le ).
- (en) Stuart L. Pimm, « The structure of food webs », Theoretical Population Biology, vol. 16, no 2, , p. 144–158 (ISSN 0040-5809, DOI 10.1016/0040-5809(79)90010-8, lire en ligne, consulté le )
- (en) Kevin McCann et Alan Hastings, « Re–evaluating the omnivory–stability relationship in food webs », Proceedings of the Royal Society of London. Series B: Biological Sciences, vol. 264, no 1385, , p. 1249–1254 (ISSN 0962-8452 et 1471-2954, DOI 10.1098/rspb.1997.0172, lire en ligne, consulté le )
- (en) Jordi Bascompte, Carlos J. Melián et Enric Sala, « Interaction strength combinations and the overfishing of a marine food web », Proceedings of the National Academy of Sciences of the USA., vol. 102, no 15, (ISSN 0962-8452, DOI 10.1073/pnas.0501562102)
- (en) Juan Camacho, Roger Guimerà et Luís A. Nunes Amaral, « Robust Patterns in Food Web Structure », Physical Review Letters, vol. 88, no 22, , p. 228102 (ISSN 0031-9007 et 1079-7114, DOI 10.1103/PhysRevLett.88.228102, lire en ligne, consulté le )
- (en) Ann E. Krause, Kenneth A. Frank, Doran M. Mason et Robert E. Ulanowicz, « Compartments revealed in food-web structure », Nature, vol. 426, no 6964, , p. 282–285 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature02115, lire en ligne, consulté le )
- (en) Frédéric Briand, « Environmental Control of Food Web Structure », Ecology, vol. 64, no 2, , p. 253–263 (ISSN 0012-9658, DOI 10.2307/1937073, lire en ligne)
- (en) Frederic Briand et Joel E. Cohen, « Environmental Correlates of Food Chain Length », Science, vol. 238, no 4829, , p. 956-960 (DOI 10.1126/science.3672136, lire en ligne)
- (en) Georgina Cordone, Vanesa Salinas, Tomás I. Marina et Santiago R. Doyle, « Green vs brown food web: Effects of habitat type on multidimensional stability proxies for a highly-resolved Antarctic food web », Food Webs, vol. 25, , e00166 (DOI 10.1016/j.fooweb.2020.e00166, lire en ligne, consulté le ).
- (en) Taylor L. Price, Jennifer Harper, Steven N. Francoeur et Halvor M. Halvorson, « Brown meets green: light and nutrients alter detritivore assimilation of microbial nutrients from leaf litter », Ecology, vol. 102, no 6, (ISSN 0012-9658 et 1939-9170, DOI 10.1002/ecy.3358, lire en ligne, consulté le ).
- (en) Kejun Zou, Elisa Thébault, Gérard Lacroix et Sébastien Barot, « Interactions between the green and brown food web determine ecosystem functioning », Functional Ecology, vol. 30, no 8, , p. 1454–1465 (ISSN 0269-8463 et 1365-2435, DOI 10.1111/1365-2435.12626, lire en ligne, consulté le ).
- (en) Huijie Gan et Kyle Wickings, « Root herbivory and soil carbon cycling: Shedding “green” light onto a “brown” world », Soil Biology and Biochemistry, vol. 150, , p. 107972 (DOI 10.1016/j.soilbio.2020.107972, lire en ligne, consulté le ).
- (en) Michael I. Sitvarin, Ann L. Rypstra et James D. Harwood, « Linking the green and brown worlds through nonconsumptive predator effects », Oikos, vol. 125, no 8, , p. 1057–1068 (DOI 10.1111/oik.03190, lire en ligne, consulté le ).
- (en) Akihiko Mougi, « Coupling of green and brown food webs and ecosystem stability », Ecology and Evolution, vol. 10, no 17, , p. 9192–9199 (ISSN 2045-7758 et 2045-7758, DOI 10.1002/ece3.6586, lire en ligne, consulté le ).
- (en) Madhav P. Thakur, « Climate warming and trophic mismatches in terrestrial ecosystems: the green–brown imbalance hypothesis », Biology Letters, vol. 16, no 2, , p. 20190770 (ISSN 1744-9561 et 1744-957X, DOI 10.1098/rsbl.2019.0770, lire en ligne, consulté le ).
- (en) Paul Kardol et David A. Wardle, « How understanding aboveground–belowground linkages can assist restoration ecology », Trends in Ecology & Evolution, vol. 25, no 11, , p. 670–679 (DOI 10.1016/j.tree.2010.09.001, lire en ligne, consulté le ).
- Loi de Raymond Lindeman (1942).
- Exemple : granulés de résine plastique, enduis de pesticides qui s'y sont agglomérés, confondus avec le frai de poisson dans le garbage patch, et consommés par divers animaux aquatiques, eux-mêmes mangés par les prédateurs marins.
- (en) Mary E. Power, « Top-Down and Bottom-Up Forces in Food Webs: Do Plants Have Primacy », Ecology, vol. 73, no 3, , p. 733–746 (DOI 10.2307/1940153, lire en ligne, consulté le ).
- (en) T. C. R. White, « The importance of a relative shortage of food in animal ecology », Oecologia, vol. 33, no 1, , p. 71–86 (ISSN 0029-8549 et 1432-1939, DOI 10.1007/BF00376997, lire en ligne, consulté le ).
- (en) Dennis Chitty, « Population processes in the vole and their relevance to general theory », Canadian Journal of Zoology, vol. 38, no 1, , p. 99–113 (ISSN 0008-4301, DOI 10.1139/z60-011, lire en ligne, consulté le ).
- (en) Mark D. Hunter et Peter W. Price, « Playing Chutes and Ladders: Heterogeneity and the Relative Roles of Bottom-Up and Top-Down Forces in Natural Communities », Ecology, vol. 73, no 3, , p. 724–732 (lire en ligne, consulté le ).
- (en) « Community Structure, Population Control, and Competition on JSTOR » (consulté le ).
- (en) Robert H. MacArthur et John W. MacArthur, « On Bird Species Diversity », Ecology, vol. 42, no 3, , p. 594–598 (DOI 10.2307/1932254, lire en ligne, consulté le ).
- (en) « Latitudinal Variations in Organic Diversity on JSTOR » (consulté le ).
- (en) « Species Density of North American Recent Mammals on JSTOR » (consulté le ).
- (en) « Food Web Complexity and Species Diversity on JSTOR » (consulté le ).
- (en) Eric L. Charnov, Gordon H. Orians et Kim Hyatt, « Ecological Implications of Resource Depression », The American Naturalist, vol. 110, no 972, , p. 247–259 (lire en ligne, consulté le ).
- (en) Andrew Sih, Philip Crowley, Mark McPeek et James Petranka, « Predation, Competition, and Prey Communities: A Review of Field Experiments », Annual Review of Ecology and Systematics, vol. 16, , p. 269–311 (lire en ligne, consulté le ).
- Milinski et Heller 1978.
- (en) James Mallet et Michael C. Singer, « Individual selection, kin selection, and the shifting balance in the evolution of warning colours: the evidence from butterflies », Biological Journal of the Linnean Society, vol. 32, no 4, , p. 337–350 (ISSN 0024-4066, DOI 10.1111/j.1095-8312.1987.tb00435.x, lire en ligne, consulté le ).
- (en) Kirk O. Winemiller et Gary A. Polis, Food Webs, Springer, Boston, MA, (ISBN 9781461570097 et 9781461570073, DOI 10.1007/978-1-4615-7007-3_1, lire en ligne), p. 1–22.
- (en) Arian D. Wallach, Ido Izhaki, Judith D. Toms et William J. Ripple, « What is an apex predator? », Oikos, vol. 124, no 11, , p. 1453–1461 (ISSN 1600-0706, DOI 10.1111/oik.01977, lire en ligne, consulté le ).
- Williams, R.J., E.l. Berlow, J.a. Dunne, AL Barabasi et N.d Martinez, 2002 Two derees of separation in complex food webs, Proceedings of the National Academy of QSciences 99:12913-12916).
- Mostajir B, Amblard C, Buffan-Dubau E, De Wit, R, Lensi R & Sime-Ngando T (2012) Les réseaux trophiques microbiens des milieux aquatiques et terrestres ; J.C. Bertrand et al. Presses Universitaires de Pau et des Pays de l'Adour, p. 28 (PDF, 30 p).
- Buchan, A., Newell, S. Y., Butler, M., Biers, E. J., Hollibaugh, J. T., & Moran, M. A. (2003). Dynamics of bacterial and fungal communities on decaying salt marsh grass. Applied and Environmental Microbiology, 69(11), 6676-6687 (résumé).
- CNRS (2015) Ecologie de l’annélide polychète Arenicola marina : du cycle de vie à son rôle de bio-indicateur de la santé des écosystèmes marins.
Voir aussi
Bibliographie
- (en) Berlow EL, Dunne JA, Martinez ND, Stark PB,Williams RJ, Brose U (2009) Simple prediction of interaction strengths in complex food webs. Proceedings of the National Academy of Sciences, 106, 187–191.
- (en) Berlow EL, Neutel A, Cohen JE et al. (2004) Interaction strengths in food webs: issues and opportunities. Journal of Animal Ecology, 73, 585–598.
- (en) Brose U (2008) Complex food webs prevent competitive exclusion among producer species. Proceedings of the Royal Society B: Biological Sciences, 275, 2507–2514.
- (en) Brose U (in press) Body-mass constraints on foraging behaviour determine population and food-web dynamics. Functional Ecology: doi:10.1111/j.1365-2435.2009.01618.x.
- (en) Emmerson M, Yearsley JM (2004) Weak interactions, omnivory and emergent food-web properties. Proceedings of the Royal Society B-Biological Sciences, 271, 397–405.
- (en) Rall BC, Guill C, Brose U (2008) Food-web connectance and predator interference dampen the paradox of enrichment. Oikos, 117, 202–213.
Articles connexes
Liens externes
- Notices dans des dictionnaires ou encyclopédies généralistes :