Bactériorhodopsine
La bactériorhodopsine est une petite protéine qu'on trouve chez certaines archées, notamment les halobactéries, où elle fonctionne comme une pompe à protons utilisant l'énergie lumineuse pour générer un gradient de protons à travers la membrane cellulaire.
| 001 | Glu | Ala | Gln | Ile | Thr | Gly | Arg | Pro | Glu | Trp |
| 011 | Ile | Trp | Leu | Ala | Leu | Gly | Thr | Ala | Leu | Met |
| 021 | Gly | Leu | Gly | Thr | Leu | Tyr | Phe | Leu | Val | Lys |
| 031 | Gly | Met | Gly | Val | Ser | Asp | Pro | Asp | Ala | Lys |
| 041 | Lys | Phe | Tyr | Ala | Ile | Thr | Thr | Leu | Val | Pro |
| 051 | Ala | Ile | Ala | Phe | Thr | Met | Tyr | Leu | Ser | Met |
| 061 | Leu | Leu | Gly | Tyr | Gly | Leu | Thr | Met | Val | Pro |
| 071 | Phe | Gly | Gly | Glu | Gln | Asn | Pro | Ile | Tyr | Trp |
| 081 | Ala | Arg | Tyr | Ala | Asp | Trp | Leu | Phe | Thr | Thr |
| 091 | Pro | Leu | Leu | Leu | Leu | Asp | Leu | Ala | Leu | Leu |
| 101 | Val | Asp | Ala | Asp | Gln | Gly | Thr | Ile | Leu | Ala |
| 111 | Leu | Val | Gly | Ala | Asx | Gly | Ile | Met | Ile | Gly |
| 121 | Thr | Gly | Leu | Val | Gly | Ala | Leu | Thr | Lys | Val |
| 131 | Tyr | Ser | Tyr | Arg | Phe | Val | Trp | Trp | Ala | Ile |
| 141 | Ser | Thr | Ala | Ala | Met | Leu | Tyr | Ile | Leu | Tyr |
| 151 | Val | Leu | Phe | Phe | Gly | Phe | Thr | Ser | Lys | Ala |
| 161 | Glx | Ser | Met | Arg | Pro | Glu | Val | Ala | Ser | Thr |
| 171 | Phe | Lys | Val | Leu | Arg | Asn | Val | Thr | Val | Val |
| 181 | Leu | Trp | Ser | Ala | Tyr | Pro | Val | Val | Trp | Leu |
| 191 | Ile | Gly | Ser | Glu | Gly | Ala | Gly | Ile | Val | Pro |
| 201 | Leu | Asn | Ile | Glu | Thr | Leu | Leu | Phe | Met | Val |
| 211 | Leu | Asp | Val | Ser | Ala | Lys | Val | Gly | Phe | Gly |
| 221 | Leu | Ile | Leu | Leu | Arg | Ser | Arg | Ala | Ile | Phe |
| 231 | Gly | Glu | Ala | Glu | Ala | Pro | Glu | Pro | Ser | Ala |
| 241 | Gly | Asp | Gly | Ala | Ala | Ala | Thr | Ser | ||
| Séquence peptidique de la bactériorhodopsine[1]. | ||||||||||
On trouve cette protéine notamment dans la membrane pourpre de l'archée halophile Halobacterium salinarum.
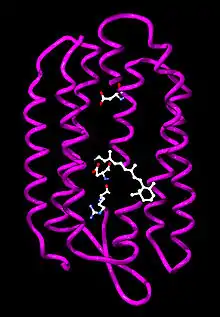
Structure
Cette protéine de 248 acides aminés se présente sous forme d'un homotrimère à symétrie cylindrique. Chacune des trois unités identiques a une structure en sept hélices α transmembranaires — structure dite opsine — emprisonnant un chromophore, ici une molécule de rétinal liée au résidu lysine 216 par une base de Schiff protonée.
Fonctionnement biologique
La molécule se comporte globalement comme une pompe à protons générant une différence de pH entre la zone extracellulaire et l'intérieur de la cellule, ce gradient de protons permettant à son tour de produire de l'ATP.
Le pompage des protons hors de la cellule est actionné par les changements conformationnels de la bactériorhodopsine induits par l'alternance entre la forme 13-cis, 15-syn excitée et la forme all-trans, 15-anti à l'état fondamental du rétinal[2] :
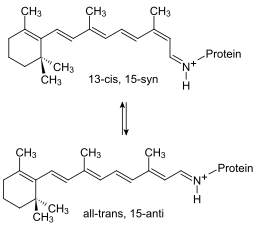 Changements de conformation de la bactériorhodopsine. |
Le mécanisme du pompage peut être résumé de la façon suivante :
- État L : l'excitation de la bactériorhodopsine par un photon — maximum d'absorption à 568 nm — déclenche directement le passage de la forme all-trans protonée à la forme 13-cis protonée du rétinal.
- État M : le rétinal 13-cis transfère le proton de sa base de Schiff au résidu aspartate 85.
- État N : le rétinal 13-cis reprotone sa base de Schiff à partir du résidu aspartate 96, ce dernier tirant son proton de l'intérieur de la cellule. Dans le même temps, le proton stationnant sur le résidu aspartate 85 est expulsé vers l'extérieur de la cellule en direction du résidu arginine 82 à travers un réseau de molécules d'eau maintenues ensemble par des liaisons hydrogène.
Le rétinal reprotoné retrouve sa configuration all-trans et la protéine, dont les charges électriques internes se sont rééquilibrées, reprend sa conformation initiale. Chacun de ces photocycles dure in vivo de 10 à 20 ms.
Utilisation nano-informatique
La simplicité de la bactériorhodopsine en a fait un modèle pour l'étude de la bioénergétique et pour le transport membranaire. Elle intéresse aussi l'industrie du stockage de données, dans la mesure où elle pourrait servir d'unité de stockage extrêmement miniaturisée pilotable par des impulsions lumineuses (à raison d'un bit par molécule, un disque de 12 cm de diamètre pourrait contenir de 20 à 50 To). L'emploi de la bactériorhodopsine constitue l'une des premières applications de l'électronique moléculaire organique, une discipline émergente de la nano-informatique.
Notes et références
- (en) H. Gobind Khorana, Gerhard E. Gerber, Walter C. Herlihy, Christopher P. Gray, Robert J. Anderedd, Kayoro Nihei et Klaus Biemann, Amino acid sequence of bacteriorhodopsin, Proc. Natl. Acad. Sci. USA, Vol. 76, No. 10, pp. 5046-5050, Octobre 1979.
- (en) Shigehiko Hayashi, Emad Tajkhorshid et Klaus Schulten, « Molecular Dynamics Simulation of Bacteriorhodopsin's Photoisomerization Using Ab Initio Forces for the Excited Chromophore », Biophysical Journal, vol. 85, no 3, , p. 1440-1449 (PMID 12944261, DOI 10.1016/S0006-3495(03)74576-7, lire en ligne)
Voir aussi
Bibliographie
- Science, 1997, p. 277-1676.
- Nature, 1999, p. 401-822.
- Nature, 2000, p. 406-645.
- Nano-informatique et Intelligence Ambiante, Jean-Baptiste Waldner, Hermes Science, London, 2006, (ISBN 2-7462-1516-0), p. 204-215
Articles connexes
Liens externes
- Thése de doctorat de l'École Polytechnique – 19 septembre 2005 « Polarisation ultrarapide et mouvements vibrationnels dans la bactériorhodopsine étudiés par spectroscopie cohérente d’émission infrarouge », par Anne Colonna.