Grand échange faunique interaméricain
Le grand échange faunique interaméricain[1] est un évènement paléozoogéographique majeur du Cénozoïque tardif, durant lequel la faune terrestre et d'eau douce a migré d'Amérique du Nord vers l'Amérique du Sud et inversement. Il est survenu lorsque la formation de l'isthme de Panama a relié les deux parties du continent (auparavant séparées) et permis la jonction de l'écozone néotropique avec l'écozone néarctique. Bien qu'il y ait eu des dispersions antérieures, probablement par voie marine, la migration s'est accélérée de façon spectaculaire à la suite de l'apparition du corridor biologique méso-américain il y a environ 2,8 Ma (Plaisancien)[2]. Constaté par la biostratigraphie et la néontologie, cet événement a eu un impact très important sur la zoogéographie des mammifères. Il a également permis la migration entre les deux Amériques de reptiles, d'amphibiens, d'arthropodes, d'oiseaux inaptes au vol et même de poissons d'eau douce. L'apparition d'un pont terrestre entre les deux continents ne semble en revanche pas avoir eu d'impact significatif sur la flore : des études biogéographiques moléculaires ont en effet montré des temps de divergence génétique beaucoup plus précoces pour les plantes et des exemples répétés de dispersion à longue distance à travers les barrières océaniques[3].
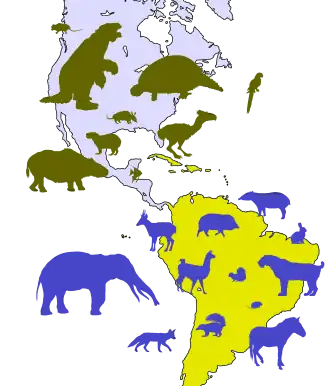
Très étudié depuis les années 1970, cet échange faunique a représenté une expérimentation grandeur nature des théories évolutionnistes en illustrant la compétition créée entre les espèces partageant une même niche écologique[4]. Des phénomènes analogues se sont produits plus tôt durant le Cénozoïque, lorsque les masses terrestres autrefois isolées de la plaque indienne[5] et de l'Afrique[6] sont entrées en contact avec l'Eurasie, il y a respectivement 50 et 30 Ma.
L'isthme de Panama a été rouvert au début du XXe siècle par les 77 kilomètres du canal de Panama (ouvert à la circulation des navires en 1914). Seuls les oiseaux et quelques espèces d'insectes peuvent encore facilement passer d'une Amérique à l'autre, même si quelques propagules et animaux circulent via les bateaux, colis ou véhicules.
Contexte zoologique nord-américain
L'Amérique du Nord forme une masse continentale unique dès le début du Tertiaire. Mais contrairement à sa voisine du sud, elle reste connectée par intermitence aux autres continents (Europe, Asie, puis Afrique) durant tout le Cénozoïque via des ponts terrestres[7]. Le plus ancien est la « route de De Geer » qui reliait le Groenland à la Fennoscandie entre la fin du Crétacé et le début du Paléocène (environ 63 Ma)[8]. Elle est remplacée bien plus tard par la « route thuléenne » qui connecte à deux reprises le même Groenland aux îles Britanniques via les îles Féroé et l'Islande actuelles entre 57 et 56 Ma[8]. À l'ouest, la Béringie permet le passage en Asie durant la majeure partie du Cénozoïque, mais sa latitude plus élevée encore qu'aujourd'hui (75° nord au Paléocène) limite les migrations[7]. Jusqu'au milieu de l'Oligocène, le détroit de Tourgaï sépare l'Europe de l'Asie au niveau de la mer Caspienne, bien que des passages aient pu exister temporairement[9]. Enfin, l'Afrique reste relativement isolée de l'Eurasie par la Téthys jusqu'à la fin de l'Éocène, même si certains échanges semblent s'être produits par l'actuel détroit de Gibraltar[10].
Mammifères
À partir de l'Oligocène, la faune mammalienne nord-américaine est dominée par les représentants des groupes connus encore aujourd'hui sur la plupart des continents, comme les ongulés, les carnivores, les rongeurs ou les chauve-souris. Ces animaux regroupés dans le clade des Boreoeutheria se sont développés dans l'hémisphère nord après leur séparation avec les Xenarthra (Amérique du Sud) et les Afrotheria (Afrique), puis ont formé différents assemblages fauniques au gré des migrations successives entre les trois continents boréaux.
Parmi les ongulés modernes, les périssodactyles (caractérisés par un nombre impair de doigts aux membres postérieurs) montraient une diversité bien plus importante qu'aujourd'hui. On estime que les premiers représentants de l'ordre seraient apparus en Asie, puis auraient migré en Europe et en Amérique du Nord vers la fin du Paléocène[11]. Les premiers équidés font leur apparition au début de l'Éocène en Amérique du Nord avec Eohippus puis suivent une histoire évolutive complexe faite de nombreuses diversifications et migrations. Leur diversité au Miocène est telle, qu'on a trouvé dans une carrière du Nebraska les fossiles de pas moins de douze espèces distinctes qui cohabitaient à la même période il y a 12 Ma[12]. L'unique genre moderne Equus s'est finalement développé sur le continent nord-américain au Pliocène, avant de migrer vers les autres continents, où il s'est diversifié pour donner naissance aux populations actuelles de zèbres, d'ânes et de chevaux[13]. Les rhinocéros apparaissent en Asie (en Chine et en Mongolie) et migrent en Amérique du Nord à la fin de l'Éocène. Le premier fossile connu du continent, Teletaceras, avait la taille d'un Berger allemand, puis ses descendants atteignent celle d'une vache. Les derniers rhinocéros nord-américains disparaissent au début du Pliocène et ne participent donc pas au Grand échange[12]. L'origine des tapirs est quant à elle débattue, car le registre fossile montre leur apparition presque simultanée en Amérique du Nord, en Europe et en Asie. Les plus anciennes attestations de la famille (genre Protapirus (en)) datent du début de l'Oligocène en Allemagne, mais une origine européenne est peu probable[14].
Les artiodactyles (au nombre de doigts pair) sont aujourd'hui beaucoup plus nombreux et diversifiés. Les plus anciens fossiles connus datent du tout début de l'Éocène et apparaissent simultanément en Eurasie et en Amérique du Nord. Il a été proposé qu'ils auraient évolué durant le Paléocène dans l'isolement de l'Inde qui était alors une île-continent, puis se seraient rapidement dispersés après son rattachement à l'Asie, mais aucun fossile indien de cette époque ne supporte cette hypothèse[15]. En Amérique du Nord, les camélidés apparaissent il y a 40 Ma et restent confinés à ce continent jusqu'au Pliocène, époque à laquelle ils migrent à la fois en Amérique du Sud (lamas, vigognes) et en Asie (chameaux, dromadaires)[16]. Les pécaris (Tayassuidae) sont des animaux proches des suidés, dont les ancêtres communs évoluaient en Asie orientale (Chine et Thaïlande) durant l'Éocène moyen. Ils se dispersent en Amérique du Nord pendant l'Oligocène et disparaissent ailleurs, certainement à cause de la compétition avec leurs cousins porcins. Ces derniers, comme d'ailleurs les hippopotames, restent confinés à l'Ancien Monde[15]. L'important groupe des ruminants apparait vraisemblablement il y a 50 Ma en Asie du Sud-Est et se dispersent rapidement jusqu'en Amérique du Nord. Les premiers représentants ont la taille d'un lapin actuel, se nourrissent plutôt de fruits, d'insectes et de jeunes pousses et n'ont pas encore la panse (rumen) qui les caractérise aujourd'hui[17]. Une seconde radiation remplace ces premiers ruminants (dont ne subsistent que les chevrotains hors d'Amérique) au début du Miocène et conduit au développement d'une famille toujours endémique du continent nord-américain : les Antilocapridae. L'unique espèce actuelle, nommée à tort « Antilope d'Amérique », est la seule survivante d'un groupe très diversifié qui comptait pas moins de 12 lignées distinctes au Miocène tardif[15]. Les cervidés et les bovidés se développent durant la même période dans l'Ancien Monde, puis envahissent l'Amérique du Nord à partir de 6 Ma, lors de l'extension des prairies dans l'hémisphère nord[17]. Malgré de nombreuses extinctions, ils constituent toujours une part importante de la mégafaune nord-américaine : bison, bœuf musqué, mouflon canadien, chèvre des montagnes Rocheuses (Bovidae), caribou, orignal, cerf de Virginie (Cervidae), etc[15].
Contexte zoologique sud-américain
À la suite du morcellement du Gondwana (un supercontinent qui réunissait à la fois l'Amérique du Sud, l'Antarctique, l'Afrique, Madagascar, l'Inde et l'Australie) à la fin du Crétacé, l'Amérique du Sud se retrouva séparée des autres terres continentales. Elle demeura une île-continent pendant la majeure partie du Cénozoïque. Son isolement permit à une faune très diverse et originale, en grande partie éteinte aujourd'hui, de s'y développer[18].
Mammifères endémiques

La faune mammalienne sud-américaine était constituée à l'origine de marsupiaux et de placentaires ayant suivi une évolution différenciée à partir d'ancêtres communs aux autres continents. D'autres mammifères non thériens étaient également présents au Paléocène (Monotremata, Gondwanatheria, Dryolestida (en) et peut-être Multituberculata). Bien qu'aucun de ceux-ci ne se soit diversifié de manière significative et que la plupart des lignées aient rapidement disparu, des formes telles que Necrolestes (en) ou Patagonia (en) ont survécu jusqu'au Miocène[19].
Les marsupiaux ont probablement pu migrer d'Amérique du Sud vers l'Australie et inversement, à travers l'Antarctique, entre la fin du Crétacé et le début du Tertiaire[20]. L'actuel Monito del monte est le seul représentant américain du super-ordre des Australidelphia qui regroupe tous les marsupiaux australiens, et sa position basale dans la phylogénie de ce groupe prouve que ces animaux sont apparus d'abord en Amérique du Sud, avant de coloniser l'Australie[20]. Les Ameridelphia, qui comprennent les rats marsupiaux et les opossums, sont quant à eux typiquement sud-américains. On comptait parmi ces derniers quelques macroprédateurs tel Thylophorops (en)[21]. Groupe frère des marsupiaux, les sparassodontes constituaient un ordre de mammifères métathériens aujourd'hui éteint qui présentaient certaines similitudes avec les actuels carnivores placentaires (convergence évolutive). Les Borhyaenidae (en) ressemblaient ainsi aux hyènes, alors que Thylacosmilus avait l'apparence d'un tigre à dents de sabre comme Smilodon[22].
Parmi les quatre principaux clades de mammifères placentaires, celui des Xenarthra s'est développé de façon très originale dans l'isolement du continent sud-américain. Il a connu une grande diversification écologique à partir d'un ancêtre probablement myrmécophage[23], alors que ce régime alimentaire a plutôt constitué une spécialisation chez les mammifères d'autres continents[note 1]. Il n'est plus représenté aujourd'hui que par trois groupes très différenciés : les tatous, les fourmiliers et les paresseux. Mais les fossiles montrent une diversité bien plus importante : on comptait ainsi des tatous géants, comme les Pampatheriidae ou les glyptodons, des espèces prédatrices comme Macroeuphractus (en), ainsi que divers paresseux terrestres (dont Megatherium, de la taille d'un éléphant) et même semi-aquatiques (comme Thalassocnus)[23].
.jpg.webp)
L'Amérique du Sud comprenait enfin un certain nombre d'ongulés endémiques, dont le lien de parenté avec ceux des autres continents reste discuté. Parfois regroupés dans le clade unique des Meridiungulata (en) (« ongulés du Sud »), ils seraient finalement plus proches des périssodactyles (rhinocéros, équidés et tapirs) et descendraient d'ongulés primitifs nord-américains (Condylarthra) parvenus, on ne sait comment, à atteindre l'Amérique du Sud[24]. On les répartit habituellement en cinq ordres : Xenungulata (en), Pyrotheria (en), Astrapotheria, Notoungulata et Litopterna, dont seuls les deux derniers ont survécu jusqu'au grand échange du Pliocène[25]. Parmi ceux-ci, on comptait certaines formes étranges, comme Macrauchenia, une sorte de dromadaire dotée d'une trompe. D'autres types présentaient des silhouettes plus familières et constituent des exemples de convergence évolutive avec les placentaires des autres continents : ainsi, Thoatherium (en) avait des membres comparables à ceux des chevaux se terminant par un seul doigt, Pachyrukhos ressemblait à un lapin, Toxodon avait l'allure d'un hippopotame et Trigodon (en) possédait une corne sur son museau comme le rhinocéros[26].
Oiseaux géants

L'une des particularités remarquables des écosystèmes sud-américains du début du Cénozoïque est l'absence de grands prédateurs placentaires, alors qu'ils dominent dans cette position dans le reste du monde[27]. Les fossiles découverts sur le continent montrent au contraire un assemblage impressionnant d'oiseaux zoophages : faucons, aigles et alliés, cigognes et alliés, anhingidas, condors et vautours du Nouveau Monde, chouettes et hiboux, ainsi que d'autres rapaces aujourd'hui éteints, comme les Teratornithidae[27]. Parmi ces derniers figurait Argentavis, probablement le plus grand oiseau ayant jamais volé[28]. Connu au Miocène supérieur de l'Argentine, il pesait jusqu'à 72 kg, pour une envergure de près de 7 mètres[29].
Plus spécifiquement, une famille d'oiseaux inaptes au vol, les Phorusrhacidae, semblent avoir profité de la niche écologique laissée vide par les mammifères pour se développer comme grands prédateurs « coureurs ». Il a ainsi été suggéré que ces « oiseaux-terreur » auraient déplacé les grands sparassodontes durant le Paléogène et seraient peut-être à l'origine de l'extinction de la famille des Proborhyaenidae (en)[30]. Tous n'étaient pas géants (de 70 à 240 cm selon les espèces), mais partageaient une certaine unité anatomique, notamment caractérisée par un bec puissant, très comprimé latéralement et terminé par un crochet comme chez les rapaces[31]. Ils étaient probablement capables d'avaler leurs petites proies entières et de déchiqueter les plus grosses au moyen de leurs griffes. Comme les cariamas actuels qui leur sont étroitement apparentés, ils régurgitaient peut-être des pelotes[32]. Très diversifiée en Amérique du Sud, la famille a longtemps été considérée comme endémique du continent. Mais la découverte récente de fossiles de Phorusrhacidae en Afrique (Algérie) et en Europe (France, Suisse) datant de l'Éocène remet ce modèle en question. La distribution de ces oiseaux implique donc des dispersions à travers des étendues marines, et donc la traversée de l'océan Téthys (pour atteindre l'Europe) et de l'Atlantique Sud[1]. La question de l'origine africaine ou sud-américaine des Phorusrhacidae reste sujette à débat, mais il semble clair que les « oiseaux-terreur » ont trouvé en Amérique du Sud des conditions qui leur ont permis de se diversifier et de survivre plus longtemps que sur les autres continents[33].
Une seconde famille d'oiseaux terrestres géants n'est connue que très partiellement à travers l'unique genre Brontornis (en). Ils étaient parfois classés parmi les Phorusrhacidae, mais la plupart des auteurs s'accordent désormais à les considérer come une famille à part, les Brontornithidae[34]. Ils semblent plutôt adaptés aux milieux fermés, ce qui expliquerait leur disparition supposée au début du Miocène, lorsque les changements climatiques ont entraîné une diminution des zones boisées au profit des steppes[35].
Immigrants venus de la mer

L'invasion du continent sud-américain a débuté il y a environ 40 Ma (Éocène moyen) avec une première vague migratoire de rongeurs, très probablement venus d'Afrique à travers l'océan Atlantique[36]. La distance séparant alors l'Afrique de l'Ouest du Brésil était moins importante qu'aujourd'hui. Par ailleurs, leur traversée a pu être facilitée par l'existence d'un chapelet d'îles (comme les rochers Saint-Pierre et Saint-Paul) et de courants marins orientés favorablement d'Est en Ouest. On suppose que la migration a eu lieu accidentellement sur des radeaux de végétation, constitués de bois flottés ou de mangrove détachées du continent, qui auraient dérivé en transportant des groupes d'animaux comportant au moins une femelle fertile. Ce modèle de dispersion s'applique également à la colonisation successive par les rongeurs des Caraïbes : ils atteignent en effet les Grandes Antilles au début de l'Oligocène[37].
Sur le continent, les rongeurs connaissent un succès fulgurant, au détriment des petits marsupiaux herbivores, et se diversifient rapidement en une multitude d'espèces. Au fil du temps, ils évoluent vers des formes plus grandes qui concurrencent certains des ongulés endémiques et contribuent peut-être à la disparition de plusieurs de ces derniers après le début de l'Oligocène. Au Pliocène, certains rongeurs sud-américains comme Josephoartigasia atteignent des tailles de l'ordre de 1 500 kg[38]. Regroupés aujourd'hui dans le micro-ordre des Caviomorpha, les représentants actuels forment quatre clades bien différenciés : Cavioidea (agoutis, pacas, pacaranas, cobayes, maras, capybaras, etc.), Erethizontoidea (porcs-épics du Nouveau Monde), Chinchilloidea (chinchillas, viscaches et rats-chinchillas) et Octodontoidea (octodons, rats à peigne, rats épineux, ragondins, hutias, etc.)[36].
Une deuxième vague de migration se serait produite, il y a environ 25 Ma, et serait à l'origine de la présence des primates. Tout comme les rongeurs, ils étaient probablement originaires d'Afrique et seraient arrivés sur le continent sud-américain de la même manière. Les espèces capables de migrer devaient être de petite taille. Ne rencontrant pas non plus de concurrence sérieuse, ils se diversifièrent rapidement et donnèrent naissance aux singes du Nouveau Monde.
On retrouve des tortues terrestres sur le continent sud-américain depuis l'Oligocène. Pendant longtemps, on a pensé qu'elles étaient originaires d'Amérique du Nord, mais une étude de génétique comparative a démontré récemment que les membres sud-américains avaient pour plus proches parents les Kinixys, un genre de tortue terrestre africain. Les tortues ont pu franchir les océans grâce à leurs extraordinaires aptitudes à la survie : elles sont capables de flotter en maintenant la tête hors de l'eau et de rester sans manger pendant 6 mois. Les tortues sud-américaines auraient, par la suite, colonisé les îles Galápagos et les Caraïbes.
Contexte tectonique et climatique
Fermeture de l'isthme
_-_Speciation_of_marine_organisms_(w_annot).png.webp)
La formation de l'isthme de Panama a débuté par la subduction de la plaque Farallon, dont est issue l'actuelle plaque de Cocos, sous les plaques caraïbe et sud-américaine, entraînant le développement d'un arc volcanique. Ce dernier est apparu aux environs de 73 Ma lorsque la plaque caraïbe a commencé à se déplacer vers l'est, atteignant sa position actuelle vers 50 Ma. En parallèle, la convergence des plaques caraïbe et sud-américaine a produit la surrection à la fois des Andes du Nord et de la plaque de Panama[2]. La collision entre cette dernière et l'Amérique du Sud a débuté aux alentours de 23 - 25 Ma[39]. Durant cette période, l'arc de Panama formait vraisemblablement une série d'îles séparées par des détroits, dont certains atteignaient 1 200 m de profondeur, permettant des échanges encore importants entre l'océan Pacifique et la mer des Caraïbes. Ces échanges d'eau profonde sont interrompus autour de 9,2 Ma, mais des eaux de surface continuent de passer, suggérant de forts courants interocéanique (qui dissuadent peut-être la migration des espèces terrestres)[2]. Enfin, on enregistre un pic de divergence des espèces marines de la zone néritique entre les deux océans vers 3,2 Ma[2]. Ces observations, couplées à celles des contrastes des assemblages de plancton marin et de salinité des eaux, fixent la fermeture de l'isthme stricto sensu à 2,8 Ma[2]. Cet événement coïncide avec un épisode de glaciation qui a donné lieu à un abaissement significatif du niveau de la mer[2].
Cette datation dite « classique » et communément acceptée depuis les années 1970 a été récemment remise en question par plusieurs chercheurs. Selon certains, un premier pont terrestre serait apparu vers 23 Ma et la fermeture de l'isthme serait intervenue plus tôt, entre 10 et 6 Ma [40]. Pour d'autres, le passage maritime entre les deux océans a été définitivement interrompu dès 15 Ma[41]. Les hypothèses d'une apparition plus ancienne du pont terrestre demandent de repenser en profondeur les causalités des changements environnementaux, écologiques et évolutifs de la région. Elles impliquent également des révisions majeures de la compréhension qu'on a de la paléocéanographie mondiale et du changement climatique, ainsi que de trouver une explication alternative pour le Grand échange. Concernant ce dernier, on évoque notamment la possibilité d'un biais dans la datation des fossiles, ou l'existence de barrières écologiques, plutôt que géologiques, ayant empêché des vagues migratoires plus anciennes[42]. Ces nouveaux modèles ne font pas l'unanimité, et on leur a notamment opposé la similitude avec l'archipel indonésien, où l'on constate de vastes échanges maritimes entre les océans Pacifique et Indien à travers des passages peu nombreux et étroits tout en maintenant une séparation complète des faunes terrestres (ligne Wallace) des deux plateaux continentaux de la Sonde et du Sahul[43].
Début des glaciations quaternaires

Malgré la présence d'un passage entre les deux continents, au moins formé d'une série d'îles, dès la fin du Miocène, la migration des espèces terrestres est restée très limitée avant 3 Ma. Il semble donc crédible que le les conditions climatiques aient été le principal paramètre de l'initiation et du déroulement du Grand échange faunique[45].
L'analyse d'un certain nombre de communautés végétales du Néogène ont ainsi indiqué que l'Amérique centrale était dominée durant le Miocène par la fôret tropicale humide, comme elle l'est encore aujourd'hui[46]. Il est vraisemblable que cela ait constitué, et constitue encore, une barrière écologique majeure à la dispersion des espèces. Ces conditions ont commencé à changer au Pliocène, lorsqu'une période glaciaire a entraîné l'extension de la savane dans la région isthmique, ce qui est largement compatible avec les adaptations écologiques et la diversité des taxons qui ont migré durant le Grand échange[44]. En effet, après un optimum vers le milieu du Miocène (15 Ma), le climat est devenu saisonnier, de plus en plus froid et sec[45]. L'inlandsis antarctique s'est alors formé au sud, mais les régions boréales sont restées sans banquise ni glaciers jusqu'au Plaisancien[47]. La glaciation de l'hémisphère nord a débuté vers 3,6 Ma, avec une augmentation progressive du volume de glace, et s'est terminée vers 2,4 Ma[48]. Il est couramment suggéré qu'elle ait été directement liée à la fermeture de l'isthme de Panama, qui aurait induit une augmentation du déplacement de sel et de chaleur vers les pôles et définitivement altéré la circulation thermohaline de l'océan Atlantique[49]. Il s'est pourtant agi d'un processus lent et progressif : malgré un premier épisode glaciaire vers 3,3 Ma, la période 3,25–3,05 Ma marque l'optimum climatique du Pliocène, et il est très plausible que l'Amérique centrale ait connu un environnement tropical au moins jusqu'à 2,7 Ma[45]. Le Pléistocène est ensuite marqué par plusieurs épisodes glaciaires, qui correspondent peut-être aux différentes vagues de migration des mammifères terrestres enregistrées pendant le Grand échange[50].
La reconstruction de la végétation d'Amérique centrale au moment du dernier maximum glaciaire (18 000 ans avant le présent) reflète un climat nettement plus froid qu'aujourd'hui, avec des températures de surface de 4 à 7 °C plus basses, et une des couvertures forestières probablement plus ouvertes, en raison des niveaux plus faibles de CO2 atmosphérique et de précipitations moindres (de 30% à 50%)[51]. On observait aussi du côté des Caraïbes une zone côtière plus étendue qu'aujourd'hui grâce à l'abaissement du niveau de la mer consécutif à la mobilisation des eaux dans les calottes glaciaires[52]. Ces conditions semblent avoir perduré jusqu'à 10 500 ans avant le présent et les forêts tropicales de basses terres qui couvrent désormais le pont terrestre entre les deux Amériques ne seraient apparues qu'après cette période. Les mammifères adaptés à la savane ont donc pu bénéficier le long des côtes de l'Amérique centrale des habitats secs et ouverts nécessaires à leur dispersion durant la majeure partie du Pléistocène[51].
Géochronologie des échanges
Le Grand échange est traditionnellement divisé en deux phases : la première inclut les animaux qui ont rejoint l'un ou l'autre continent avant la formation du pont terrestre, la seconde comprend les migrations beaucoup plus importantes et rapprochée survenues après la surrection totale de l'isthme (datée autour de 3 Ma). Ainsi, aux premiers « hérauts du Sud[53] » parvenus en Amérique du Nord au Miocène supérieur ont succedé plusieurs dispersions épisodiques, avant que les « légions[53] » des deux continents ne se mélangent à intervalles réguliers à la jonction Pliocène-Pléistocène. Cette seconde phase est celle auquel se réfère le plus souvent le terme de Grand échange faunique sticto sensu (Great American Biotic Interchange, GABI) et comprend elle-même quatre épisodes distincts[50], alors que les migrations antérieures sont nommées Pre-GABI[54] ou Proto-GABI[55].
L'événement a été particuliérement étudié pour les mammifères terrestres, pour lesquels on dispose de nombreuses preuves paléontologiques. Les échanges fauniques pour les autres groupes d'animaux sont nettement moins documentés, car ils n'ont laissé que peu de fossiles. Néanmoins, l'analyse de l'horloge moléculaire a montré que l'apparition d'un pont terrestre entre les deux continents a également influencé la migration des oiseaux, particuliérement celle des groupes spécialisés dans les forêts tropicales, qui colonisent rarement les îles océaniques et ont de faibles capacités de dispersion à travers les barrières maritimes (comme les Thamnophilidae ou les Dendrocolaptidae). Contrairement aux mammifères, ces échanges se sont principalement déroulés du Sud vers le Nord[56].
Une troisième vague majeure d'échanges est intervenue à la suite du développement de la forêt tropicale en Amérique centrale, mais reste peu documentée en raison de la pauvreté du registre fossile dans la région[57]. Ce « mélange tropical final[53] » s'est déroulé durant le dernier demi-million d'années avant le présent, bien qu'il n'existe pas de chronologie précise, et constitue la dernière phase de constitution de l'actuelle région néotropicale. Par analogie aux « hérauts » et aux « légions » des premières phases, ces derniers migrants sont appelés les « vivandières » (en anglais : camp followers)[58].
Échanges précoces

Les premiers mammifères à avoir migré entre les deux Amériques semblent être les paresseux terrestres. Des fossiles du Miocène tardif (entre 9 et 8,5 Ma) de Thinobadistes (en) (famille des Mylodontidae) et de Pliometanastes (en) (famille des Megalonychidae) ont en effet été retrouvés en Amérique du Nord[50]. Ils auraient atteint le continent en nageant à travers de petites étendues d'eau séparant les îles de la mer des Caraïbes. Les Megalonychidae sont en effet attestés dans les Antilles dès le début de l'Oligocène, où ils seraient parvenus grâce à une voie terrestre temporaire reliant les Grandes Antilles en formation au continent sud-américain[59].
Le chemin inverse est emprunté par un carnivore de la famille des Procyonidae, Cyonasua (en), qui est présent en Amérique du Sud vers 7,3 Ma. Il est suivi par les premiers rongeurs (sous-famille des Sigmodontinae) autour de 6 Ma[50].
L'échange suivant a lieu entre 5 et 4,7 Ma. Il est marqué par l'arrivée en Amérique du Nord d'un oiseau-terreur, Titanis, dont les restes ont été retrouvés au Texas et en Floride. Probablement victime de la concurrence des grands mammifères carnivores (ursidés, canidés, félidés) qui se diversifient dans la région au Pliocène, il s'éteint avant le début du Pléistocène[60]. À la même époque, un tatou géant (Plaina, Pampatheriidae) et un autre paresseux terrestre (Glossotherium, Mylodontidae) sont attestés dans le Guanajuato, au Mexique[50].
Ces mêmes dépôts montrent la présence vers 3,9 — 3,1 Ma d'un autre tatou géant (Glyptotherium, Glyptodontidae) et d'un rongeur caviomorphe (Neochoerus (en), Caviidae)[50]. Cet animal éteint, proche des actuels capybaras, est retrouvé bien plus tard dans la pampa. Cela pourrait signifier qu'après s'être dispersé en Amérique du Nord, il aurait traversé l'isthme de Panama en sens inverse lors du deuxième épisode du grand échange (GABI 2)[61]. C'est également autour de 3,7 Ma que Pampatherium, un autre Pampatheriidae, entre au Mexique. Cela correspond au passage inverse d'un pécari (Platygonus (en)) en Argentine[50]. Enfin, vers 3,3 Ma, les premiers camélidés (genre Lama) se dispersent en Amérique du Sud. La migration vers le sud d'espèces adaptées à la savane comme les pécaris et les lamas pourrait être liée à un premier épisode de glaciation de l'hémisphère nord[62].
GABI 1
Le premier épisode de dispersion majeure entre les deux continents intervient entre 2,6 et 2,4 Ma, ce qui correspond à l'époque de la fermeture définitive de l'isthme de Panama. Les fossiles montrent ainsi les premières attestations de quatre familles de mammifères placentaires nord-américains en Amérique du Sud et, peu après, la présence de quatre familles sud-américaines de Xenarthra en Amérique du Nord[50].
La migration du Nord au Sud est illustrée par deux carnivores, un grison (Galictis, Mustelidae) et un renard († Dusicyon, Canidae), deux équidés († Hippidion et † Onohippidium) et peut-être un gomphotère (pachyderme préhistorique semblable aux éléphants). Celle du Sud au Nord est marquée par trois familles de tatous, Dasypodidae (Dasypus), Pampatheriidae († Holmesina (en)) et Glyptodontidae[note 2] († Pachyarmatherium (en)), et par un paresseux terrestre géant († Eremotherium, Megatheriidae)[50].
GABI 2

Si le premier échange semble relativement équilibré entre les deux continents, la seconde vague de migration est très clairement à l'avantage de la faune nord-américaine. On constate ainsi vers 1,8 Ma l'arrivée massive en Amérique du Sud de gros mammifères de l 'hémisphère nord. Les carnivores atteignent leur pic de diversité[63] avec des ours († Arctotherium), des félins comme les pumas (Puma ), les jaguars (Panthera) et des tigres à dents de sabre († Smilodon), et des canidés comme Cerdocyon et Chrysocyon. On note également la migration de nombreux ongulés (Artiodactyla et Perissodactyla), représentés par des pécaris († Catagonus), des camelidés († Palaeolama (en) et † Hemiauchenia (en)), des cervidés († Antifer (en) et † Epieuryceros (nl)), des tapirs (Tapirus), ainsi que deux genres de gomphotères († Stegomastodon et † Cuvieronius)[50].
La migration inverse n'est en revanche attestée que pour le tamanoir (Myrmecophaga), dont le fossile le plus ancien a été retrouvé dans le nord-ouest du Sonora, au Mexique[64]. Un paresseux terrestre (Paramylodon) est présent à la même époque en Floride, mais il pourrait s'agir d'un descendant de Glossotherium, attesté au Mexique bien avant la fermeture de l'isthme[65].
GABI 3
Un troisième échange bilatéral semble intervenir autour de 0,8 Ma. Il est marqué par le passage au Nord des opossums (Didelphis), qui concorde avec la dispersion en Amérique du Sud d'un félin, le jaguarondi (Herpailurus), d'un cervidé († Paraceros) et d'un pécari (Pecari)[50].
GABI 4
Le dernier épisode du grand échange est strictement unilatéral et se déroule il y a 125 000 ans environ. Le continent sud-américain se voit ainsi envahir par de nouveaux mammifères de l'hémisphère boréal représentant une certaine diversité écologique : coatis (Nasua), loutres (Lutra), canidés (Canis), ocelots (Leopardus), mais aussi des lapins (Sylvilagus) et des chevaux sauvages (Equus)[50]. Cette période est également marquée par le retour sur son continent d'origine du tatou géant Glyptotherium[66].
Échanges tardifs
Les forêts tropicales d'Amérique centrale et du Sud du Mexique abritent un assemblage de mammifères de taille moyenne dont les ancêtres ont traversé l'isthme de Panama à un stade tardif du Grand échange. Parmi les taxons originaires d'Amérique du Sud, on trouve notamment plusieurs Xenarthra spécialisés dans les habitats tropicaux (par opposition aux espèces généralistes ayant migré plus tôt) : des fourmiliers (tamanoir, tamandou et myrmidon), des paresseux arboricoles (paresseux tridactyle et unau) et des tatous (Cabassous centralis et Dasypus novemcinctus)[58]. On compte également une quarantaine d'espèces d'autres groupes, surtout des opposums, singes du Nouveau Monde et des rongeurs caviomorphes, ainsi que plus de 150 espèces de chauves-souris[57]. Dans le sens inverse, des espèces tropicales comme le kinkajou ou certains écureuils arboricoles pourraient avoir migré à la même époque.
Résultats et analyse
Résultats
À sa mise en place, l'échange était équilibré, les courants migratoires étant d'égale importance dans les deux sens. Mais par la suite, il s'avéra que les espèces sud-américaines réussissaient moins bien à s'implanter que les espèces nord-américaines. Alors que les espèces sud-américaines migrant vers le continent nord-américain n'arrivaient pas à concurrencer les espèces déjà présentes, les espèces nord-américaines migrant vers le continent sud-américain s'établirent par contre en grand nombre et se diversifièrent considérablement.
Bien que les oiseaux-terreurs réussirent au début à s'implanter en Amérique du Nord (il y a environ 5 Ma en utilisant l'archipel d'Amérique centrale), ce succès ne fut que temporaire. Tous les plus grands oiseaux sud-américains et les prédateurs métathériens disparurent par la suite. Les petits marsupiaux réussirent mieux, tandis que les édentés à l'aspect pourtant primitif se montrèrent, chose surprenante, fort compétitifs.
La présence aujourd'hui de tatous, d'opossums et de porcs-épics en Amérique du Nord s'explique par le grand échange inter-américain. Les opossums et les porcs-épics furent les migrants sud-américains qui connurent la plus grande réussite, atteignant respectivement le Canada et l'Alaska. Les paresseux terrestres géants qui migrèrent vers le nord connurent aussi une belle réussite, puisque Megalonyx colonisa jusqu'au Yukon et l'Alaska, et serait peut-être parvenu en Eurasie s'il n'avait subi la crise biologique de la fin du Quaternaire.
Les rongeurs de la sous-famille des Sigmodontinae (sous-famille regroupant les rats et les souris du Nouveau Monde) sont les mammifères nord-américains qui ont le mieux prospéré et qui se sont le plus diversifiés en Amérique du Sud. D'autres groupes de mammifères nord-américains ont également migré avec beaucoup de succès, notamment les canidés et les cervidés, avec chacun 6 genres présents en Amérique du Sud.
Raisons des succès et des échecs de colonisation

La dissymétrie entre les deux sens de migrations, avec plus de « victimes » d’origine sud-américaine (tel le thylacosmilus) que de nord-américaines (comme le solénodonte), résulte de plusieurs facteurs. Le premier est climatique : les espèces migrant vers le nord, une fois franchi la Sierra Nevada, sont confrontées à des conditions climatiques bien plus rudes, bien plus froides que celles qui existaient dans la forêt tropicale humide sud-américaine. Le second facteur est lié à l'aire géographique dont disposaient les ancêtres des espèces nord-américaines par rapport aux ancêtres des espèces sud-américaines. Durant le Cénozoïque, l'Amérique du Nord était reliée périodiquement à l'Eurasie via la Béringie, si bien que des échanges réciproques de leur faune étaient possibles, contribuant à l'uniformisation de celle-ci. L'Eurasie était elle-même reliée à l'Afrique, ce qui élargissait encore plus l'espace disponible. Au contraire, l'Amérique du Sud s'est trouvée très tôt dans son histoire séparée de l'Antarctique et de l'Australie. Par ailleurs, ces continents étaient de taille plus modeste, et peu d'animaux (uniquement des marsupiaux parmi les mammifères) semblent avoir réussi à circuler entre eux. Cela signifie que la faune d'Amérique du Nord disposait d'un espace terrestre pratiquement six fois plus vaste que celle d'Amérique du Sud pour se développer.
Les xénarthres, qui ont réussi à résister à la vague migratoire nord-américaine, constituent néanmoins une exception. Leur succès peut s'expliquer par les moyens de défense efficaces qu'ils ont su mettre en place pour faire face aux prédateurs : griffes puissantes, armures corporelles. Ils ont probablement développé ces moyens de défense pour compenser leur métabolisme très lent. Par ailleurs, cette lenteur leur permettait de réduire leurs besoins nutritifs au strict minimum.
Développements ultérieurs et conséquences
Extinctions de la fin du Pléistocène

À la fin du Pléistocène, il y a environ 12 000 ans, trois événements majeurs se produisirent : les ancêtres des Amérindiens colonisèrent le Nouveau Monde, la dernière glaciation prit fin et la mégafaune des Amériques connut une importante vague d'extinctions. Cette crise biologique fit de nombreuses victimes et n'épargna pas les espèces qui avaient su profiter avec succès du grand échange interaméricain. Tous les glyptodons et apparentés, les paresseux terrestres, les équidés, les proboscidiens, les loups et lions d'Amérique, et les smilodons des deux continents disparurent. Les derniers des notongulés d'Amérique centrale et du Sud s'éteignirent aussi, de même que les castors géants, les dholes, les guépards autochtones, les homothères qui peuplaient l'Amérique du Nord. Certains groupes disparurent d'Amérique du Nord, leur continent d'origine, mais survécurent dans leur nouvel habitat en Amérique du Sud ; c'est le cas des camélidés (vigognes et lamas), des tapiridés et des ours du genre Tremarctos (ours à lunette). D'autres espèces survécurent dans leur habitat d'origine mais s'éteignirent ailleurs, comme le capybara.
Il s'avère que toute la faune sud-américaine ayant migré en Amérique du Nord dont le poids dépassait 10 kg (le poids d'un grand tatou), ainsi que toute la faune sud-américaine vivant dans son habitat d'origine dont le poids dépassait 65 kg (le poids d'un fourmilier géant ou d'un grand capybara), a disparu. En revanche, le bison laineux, la plus grande espèce nord-américaine à n'avoir pas migré ayant survécu, dépasse les 900 kg, et le tapir de Baird, la plus grande espèce nord-américaine à avoir migré en Amérique du Sud ayant survécu, atteint 400 kg.
La quasi simultanéité de l'extinction de la mégafaune avec le peuplement par l'homme de l'Amérique et le retrait des glaciers suggère que ces événements ne sont pas totalement indépendants. Cependant, ce sujet prête à discussion. Bien que le changement climatique ait affecté toutes les régions du globe, la mégafaune d'Afrique et d'Asie du Sud-Est n'a pas été touchée par les extinctions massives. Par ailleurs, au cours de ces derniers millions d'années, des périodes de glaciaires et interglaciaires se sont succédé sans que cela ait un impact dramatique sur la mégafaune d'alors.
Annexes
Notes
Références
- Angst et Buffetaut 2018, p. 145.
- O’Dea et al. 2016.
- Cody et al. 2010.
- Francis Duranthon, Histoires de mammifères, Rosny, Editions Bréal, , 176 p. (ISBN 2-7495-2081-9, lire en ligne), p. 99-100.
- (en) K. Praveen Karanth, « Out-of-India Gondwanan origin of some tropical Asian biota », Current science, vol. 90, no 6, , p. 789-792 (lire en ligne [PDF]).
- (en) Stephen Blair Hedges, « Afrotheria: Plate tectonics meets genomics », Proceedings of the National Academy of Sciences, vol. 98, no 1, , p. 1–2 (ISSN 0027-8424 et 1091-6490, PMID 11136239, PMCID PMC33345, DOI 10.1073/pnas.98.1.1, lire en ligne, consulté le ).
- Rose 2006, p. 18.
- (en) Leonidas Brikiatis, « The De Geer, Thulean and Beringia routes: key concepts for understanding early Cenozoic biogeography », Journal of Biogeography, vol. 41, no 6, , p. 1036–1054 (DOI 10.1111/jbi.12310, lire en ligne, consulté le ).
- Rose 2006, p. 19.
- Rose 2006, p. 20.
- Kemp 2005, p. 260.
- Prothero 2016, chap. 15, p. 186-202.
- (en) Lorenzo Rook, Raymond L. Bernor, Leonardo S. Avilla, Omar Cirilli et al., « Mammal Biochronology (Land Mammal Ages) Around the World From Late Miocene to Middle Pleistocene and Major Events in Horse Evolutionary History », Frontiers in Ecology and Evolution, vol. 7, , p. 278 (ISSN 2296-701X, DOI 10.3389/fevo.2019.00278, lire en ligne, consulté le ).
- (en) Laureline Scherler, Damien Becker et Jean-Pierre Berger, « Tapiridae (Perissodactyla, Mammalia) of the Swiss molasse basin during the Oligocene - Miocene transition », Journal of Vertebrate Paleontology, vol. 31, no 2, , p. 479-496 (lire en ligne).
- Prothero 2016, chap. 14, p. 151-185.
- (en) Peng Cui, Rimutu Ji, Feng Ding, Dan Qi et al., « A complete mitochondrial genome sequence of the wild two-humped camel (Camelus bactrianus ferus): an evolutionary history of camelidae », BMC Genomics, vol. 8, no 1, , p. 241 (PMID 17640355, PMCID PMC1939714, DOI 10.1186/1471-2164-8-241, lire en ligne, consulté le ).
- (en) Timothy J. Hackmann et James N. Spain, « Invited review: Ruminant ecology and evolution: Perspectives useful to ruminant livestock research and production », Journal of Dairy Science, vol. 93, no 4, , p. 1320–1334 (DOI 10.3168/jds.2009-2071, lire en ligne, consulté le ).
- Simpson 1980.
- (en) Nicolás R. Chimento, Federico L. Agnolin et Fernando E. Novas, « The bizarre ‘metatherians’ Groeberia and Patagonia , late surviving members of gondwanatherian mammals », Historical Biology, vol. 27, no 5, , p. 603–623 (ISSN 0891-2963 et 1029-2381, DOI 10.1080/08912963.2014.903945, lire en ligne, consulté le )
- Nilsson et al. 2010.
- Goin et al. 2009.
- (en) Christine Argot, « Functional-adaptive features and palaeobiologic implications of the postcranial skeleton of the late Miocene sabretooth borhyaenoid Thylacosmilus atrox (Metatheria) », Alcheringa: An Australasian Journal of Palaeontology, vol. 28, no 1, , p. 229–266 (ISSN 0311-5518 et 1752-0754, DOI 10.1080/03115510408619283, lire en ligne, consulté le ).
- Gaudin et Croft 2015.
- Defler 2019.
- (en) Michael Buckley, « Ancient collagen reveals evolutionary history of the endemic South American ‘ungulates’ », Proceedings of the Royal Society B: Biological Sciences, vol. 282, no 1806, , p. 20142671 (ISSN 0962-8452 et 1471-2954, PMID 25833851, PMCID PMC4426609, DOI 10.1098/rspb.2014.2671, lire en ligne, consulté le ).
- Prothero 2016, chap. 16, p. 203-208.
- Tambussi et Degrange 2013, p. 98.
- Tambussi et Degrange 2013, p. 94.
- (en) Sankar Chatterjee, R. Jack Templin et Kenneth E. Campbell, « The aerodynamics of Argentavis, the world's largest flying bird from the Miocene of Argentina », Proceedings of the National Academy of Sciences, vol. 104, no 30, , p. 12398–12403 (ISSN 0027-8424 et 1091-6490, PMID 17609382, PMCID PMC1906724, DOI 10.1073/pnas.0702040104, lire en ligne, consulté le ).
- Prevosti et Forasiepi 2018, p. 176.
- Angst et Buffetaut 2018, p. 152.
- Angst et Buffetaut 2018, p. 161.
- (en) Delphine Angst, Eric Buffetaut, Christophe Lécuyer et Romain Amiot, « “Terror Birds” (Phorusrhacidae) from the Eocene of Europe Imply Trans-Tethys Dispersal », PLoS ONE, vol. 8, no 11, , e80357 (ISSN 1932-6203, PMID 24312212, PMCID PMC3842325, DOI 10.1371/journal.pone.0080357, lire en ligne, consulté le ).
- Angst et Buffetaut 2018, p. 166-167.
- Angst et Buffetaut 2018, p. 175.
- Poux et al. 2006.
- (en) Jorge Vélez-Juarbe, Thomas Martin, Ross D. E. Macphee et Diana Ortega-Ariza, « The earliest Caribbean rodents: Oligocene caviomorphs from Puerto Rico », Journal of Vertebrate Paleontology, vol. 34, no 1, , p. 157–163 (ISSN 0272-4634 et 1937-2809, DOI 10.1080/02724634.2013.789039, lire en ligne, consulté le ).
- (en) Virginie Millien, « The largest among the smallest: the body mass of the giant rodent Josephoartigasia monesi », Proceedings of the Royal Society B: Biological Sciences, vol. 275, no 1646, , p. 1953–1955 (ISSN 0962-8452 et 1471-2954, PMID 18495621, PMCID PMC2596365, DOI 10.1098/rspb.2008.0087, lire en ligne, consulté le ).
- Farris et al. 2011.
- Bacon et al. 2015.
- Montes et al. 2015.
- (en) Charles G. Willis et Charles C. Davis, « Rethinking migration », Science, vol. 348, no 6236, , p. 766–766 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.348.6236.766-a, lire en ligne, consulté le ).
- (en) Anthony G Coates et Robert F Stallard, « How old is the Isthmus of Panama? », Bulletin of Marine Science, vol. 89, no 4, , p. 801–813 (DOI 10.5343/bms.2012.1076, lire en ligne, consulté le ).
- Webb 1991.
- Woodburne 2010, Climate.
- (en) Alan Graham et David L. Dilcher, « Studies in Neotropical paleobotany. XII. A palynoflora from the Pliocene Rio Banano Formation of Costa Rica and the Neogene vegetation of Mesoamerica », American Journal of Botany, vol. 85, no 10, , p. 1426–1438 (DOI 10.2307/2446400, lire en ligne, consulté le ).
- Pierre Martin, Le climat dans tous ses états, Louvain-la-Neuve, De Boeck Superieur, , 192 p. (ISBN 978-2-8073-0660-8 et 2-8073-0660-8, OCLC 982061720, lire en ligne), p. 75.
- (en) Manfred Mudelsee et Maureen E. Raymo, « Slow dynamics of the Northern Hemisphere glaciation », Paleoceanography, vol. 20, no 4, , p. 1-14 (DOI 10.1029/2005PA001153, lire en ligne, consulté le ).
- (en) Gretta Bartoli, Michael Sarnthein, Mara S. Weinelt, Helmut Erlenkeuser, Dieter Garbe-Schönberg et David W. Lea, « Final closure of Panama and the onset of northern hemisphere glaciation », Earth and Planetary Science Letters, vol. 237, nos 1-2, , p. 33–44 (DOI 10.1016/j.epsl.2005.06.020, lire en ligne, consulté le ).
- Woodburne 2010, Dispersal pattern.
- Woodburne 2010, Plants and ecologies.
- Woodburne 2010, Sea level.
- Stehli et Webb 1985, chap. 14, fig. 1.
- Goin et al. 2012, p. 39.
- Cione et al. 2015, p. 2.
- Weir, Bermingham et Schluter 2009.
- Morgan 2008, p. 131.
- McDonald 2005.
- (en) Jennifer L. White et Ross D. E. MacPhee, chap. 14 « The Sloths of the West Indies : A Systematic and Phylogenetic Review », dans Charles A. Woods et Florence E. Sergile, Biogeography of the West Indies : patterns and perspectives, CRC Press, , 608 p. (ISBN 1-4200-3948-2, OCLC 71231607, lire en ligne), p. 201-235.
- Angst et Buffetaut 2018, p. 163.
- Cione et al. 2015, p. 76.
- Goin et al. 2012.
- Cione et al. 2015, p. 75.
- (en) Christopher A. Shaw et H. Gregory Mcdonald, « First Record of Giant Anteater (Xenarthra, Myrmecophagidae) in North America », Science, vol. 236, no 4798, , p. 186–188 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.236.4798.186, lire en ligne, consulté le ).
- (en) Robert K. Mcafee, « Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae) », Zoological Journal of the Linnean Society, vol. 155, no 4, , p. 885–903 (DOI 10.1111/j.1096-3642.2008.00468.x, lire en ligne, consulté le ).
- (en) Alfredo A. Carlini, Alfredo E. Zurita et Orangel A. Aguilera, « North American Glyptodontines (Xenarthra, Mammalia) in the Upper Pleistocene of northern South America », Paläontologische Zeitschrift, vol. 82, no 2, , p. 125–138 (ISSN 0031-0220 et 1867-6812, DOI 10.1007/BF02988404, lire en ligne, consulté le ).
Sur le grand échange
- [Cione et al. 2015] (en) Alberto Luis Cione, Germán Mariano Gasparini, Esteban Soibelzon, Leopoldo Héctor Soibelzon et Eduardo Pedro Tonni, The great American biotic interchange : a South American perspective, Dordrecht/New York, Springer, , 97 p. (ISBN 978-94-017-9792-4, 94-017-9792-7 et 94-017-9791-9, OCLC 908103326, lire en ligne).
- [Cody et al. 2010] (en) Sarah Cody, James E. Richardson, Valentí Rull, Christopher Ellis et R. Toby Pennington, « The Great American Biotic Interchange revisited », Ecography, , p. 326-332 (DOI 10.1111/j.1600-0587.2010.06327.x, lire en ligne, consulté le ).
- [McDonald 2005] (en) H. Gregory McDonald, « Paleoecology of extinct Xenarthrans and the Great American Biotic Interchange », Bulletin of the Florida Museum of Natural History, no 45, , p. 313-333 (lire en ligne).
- [Morgan 2008] (en) Gary S. Morgan, « Vertebrate fauna and geochronology of the Great American Biotic Interchange in North America », New Mexico Museum of Natural History and Science Bulletin, Abuquerque, vol. 44 « Neogene Mammals », , p. 93-140 (lire en ligne).
- [Stehli et Webb 1985] (en) Francis G. Stehli et S. David Webb, The Great American Biotic Interchange, New York, Springer US, , 532 p. (ISBN 978-1-4684-9181-4 et 1-4684-9181-4, OCLC 853264357, lire en ligne).
- [Weir, Bermingham et Schluter 2009] (en) Jason T. Weir, Eldredge Bermingham et Dolph Schluter, « The Great American Biotic Interchange in birds », Proceedings of the National Academy of Sciences, vol. 106, no 51, , p. 21737–21742 (ISSN 0027-8424 et 1091-6490, PMID 19996168, PMCID PMC2799814, DOI 10.1073/pnas.0903811106, lire en ligne, consulté le ).
- [Webb 1991] (en) S. David Webb, « Ecogeography and the Great American Interchange », Paleobiology, vol. 17, no 3, , p. 266–280 (ISSN 0094-8373 et 1938-5331, DOI 10.1017/S0094837300010605, lire en ligne, consulté le ).
- [Webb 2006] (en) S. David Webb, « The Great American Biotic Interchange: patterns and processes », Annals of the Missouri Botanical Garden, vol. 93, no 2, , p. 245–257 (ISSN 0026-6493, DOI 10.3417/0026-6493(2006)93[245:TGABIP]2.0.CO;2, lire en ligne, consulté le ).
- [Woodburne 2010] (en) Michael O. Woodburne, « The Great American Biotic Interchange: Dispersals, Tectonics, Climate, Sea Level and Holding Pens », Journal of Mammalian Evolution, vol. 17, no 4, , p. 245–264 (ISSN 1064-7554 et 1573-7055, PMID 21125025, PMCID PMC2987556, DOI 10.1007/s10914-010-9144-8, lire en ligne, consulté le ).
Sur la faune
- [Angst et Buffetaut 2018] Delphine Angst et Éric Buffetaut, Paléobiologie des oiseaux géants terrestres, Londres, ISTE Editions Ltd, , 280 p. (ISBN 978-1-78405-397-0 et 1-78405-397-X, OCLC 1049915246, lire en ligne).
- [Croft 2016] (en) Darin A. Croft (ill. Velizar Simeonovski), Horned armadillos and rafting monkeys : the fascinating fossil mammals of South America, Bloomington et Indianapolis, Indiana University Press, , 320 p. (ISBN 978-0-253-02094-9 et 0-253-02094-8, OCLC 951975229, lire en ligne).
- [Defler 2019] (en) Thomas Defler, « The Native Ungulates of South America (Condylarthra and Meridiungulata) », dans History of Terrestrial Mammals in South America, vol. 42, Springer International Publishing, coll. « Topics in Geobiology », , 89–115 p. (ISBN 978-3-319-98448-3, DOI 10.1007/978-3-319-98449-0_5, lire en ligne).
- [Fariña, Vizcaíno et De Iuliis 2012] (en) Richard A. Fariña, Sergio F. Vizcaíno et Gerardo De Iuliis (ill. Sebastián Tambusso), Megafauna : giant beasts of Pleistocene South America, Bloomington et Indianapolis, Indiana University Press, , 448 p. (ISBN 978-0-253-00719-3 et 0-253-00719-4, OCLC 844728531, lire en ligne).
- [Gaudin et Croft 2015] (en) Timothy J. Gaudin et Darin A. Croft, « Paleogene Xenarthra and the evolution of South American mammals », Journal of Mammalogy, vol. 96, no 4, , p. 622–634 (ISSN 0022-2372 et 1545-1542, DOI 10.1093/jmammal/gyv073, lire en ligne, consulté le ).
- [Goin et al. 2012] (en) Francisco J. Goin, Javier N. Gelfo, Laura Chornogubsky, Michael O. Woodburne et Thomas Martin, « Origins, radiations, and distribution of South American mammals : from greenhouse to icehouse worlds », dans Bruce D. Patterson et Leonora Pires Costa, Bones, clones, and biomes : the history and geography of recent neotropical mammals, Chicago et Londres, University of Chicago Press, , 432 p. (ISBN 978-0-226-64921-4 et 0-226-64921-0, OCLC 793511380, lire en ligne), p. 20-50.
- [Kemp 2005] (en) Thomas Stainforth Kemp, The origin and evolution of mammals, Oxford, Oxford University Press, , 331 p. (ISBN 978-0-19-154517-7, lire en ligne).
- [Nilsson 2010] (en) Maria A. Nilsson, Gennady Churakov, Mirjam Sommer, Ngoc Van Tran, Anja Zemann, Jürgen Brosius et Jürgen Schmitz, « Tracking Marsupial Evolution Using Archaic Genomic Retroposon Insertions », PLoS Biology, vol. 8, no 7, , e1000436 (ISSN 1545-7885, PMID 20668664, PMCID PMC2910653, DOI 10.1371/journal.pbio.1000436, lire en ligne, consulté le ).
- [Poux et al. 2006] (en) Céline Poux, Pascale Chevret, Dorothée Huchon, Wilfried W. de Jong et Emmanuel J. P. Douzery, « Arrival and Diversification of Caviomorph Rodents and Platyrrhine Primates in South America », Systematic Biology, vol. 55, no 2, , p. 228–244 (ISSN 1076-836X et 1063-5157, DOI 10.1080/10635150500481390, lire en ligne, consulté le ).
- [Prevosti et Forasiepi 2018] (en) Francisco J. Prevosti et Analía M. Forasiepi, Evolution of South American mammalian predators during the Cenozoic : paleobiogeographic and paleoenvironmental contingencies, Springer, , 196 p. (ISBN 978-3-319-03701-1, 3-319-03701-3 et 3-319-03700-5, OCLC 1018307594, lire en ligne).
- [Prothero 2016] (en) Donald R. Prothero, The Princeton field guide to prehistoric mammals, Princeton et Oxford, Princeton University Press, , 240 p. (ISBN 978-1-4008-8445-2 et 1-4008-8445-4, OCLC 960349875, lire en ligne).
- [Rose 2006] (en) Kenneth David Rose, The beginning of the age of mammals, Baltimore, Johns Hopkins University Press, , 428 p. (ISBN 978-0-8018-9221-9, 0-8018-9221-X et 978-0-8018-8022-3, OCLC 646769601, lire en ligne).
- [Simpson 1980] (en) George Gaylord Simpson, Splendid Isolation : The Curious History of South American Mammals, New Haven, Connecticut, Yale University Press, (ISBN 978-0-300-02434-0, OCLC 5219346, lire en ligne).
- [Tambussi et Degrange 2013] (en) Claudia Tambussi et Federico Degrange, South American and Antarctic continental cenozoic birds : paleobiogeographic affinities and disparities, Dordrecht, Springer, , 113 p. (ISBN 978-94-007-5467-6 et 94-007-5467-1, OCLC 823635299, lire en ligne).
Sur l'isthme de Panama
- [Bacon et al. 2015] (en) Christine D. Bacon, Daniele Silvestro, Carlos Jaramillo, Brian Tilston Smith et al., « Biological evidence supports an early and complex emergence of the Isthmus of Panama », Proceedings of the National Academy of Sciences, vol. 112, no 19, , p. 6110–6115 (ISSN 0027-8424 et 1091-6490, PMID 25918375, PMCID PMC4434730, DOI 10.1073/pnas.1423853112, lire en ligne, consulté le ).
- [Coates et al. 2004] (en) Anthony G. Coates, Laurel S Collins, Marie-Pierre Aubry et William A. Berggren, « The Geology of the Darien, Panama, and the late Miocene-Pliocene collision of the Panama arc with northwestern South America », Geological Society of America Bulletin, vol. 116, nos 11-12, , p. 1327–1344 (ISSN 0016-7606 et 1943-2674, DOI 10.1130/B25275.1, lire en ligne, consulté le ).
- [Farris et al. 2011] (en) David W. Farris, Carlos Jaramillo, German Bayona, Sergio A. Restrepo-Moreno et al., « Fracturing of the Panamanian Isthmus during initial collision with South America », Geology, vol. 39, no 11, , p. 1007–1010 (ISSN 1943-2682 et 0091-7613, DOI 10.1130/G32237.1, lire en ligne, consulté le ).
- [Montes et al. 2015] (en) Camilo Montes, A. Cardona, C. Jaramillo, A. Pardo et al., « Middle Miocene closure of the Central American Seaway », Science, vol. 348, no 6231, , p. 226–229 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.aaa2815, lire en ligne, consulté le ).
- [O’Dea et al. 2016] (en) Aaron O’Dea, Harilaos A. Lessios, Anthony G. Coates, Ron I. Eytan et al., « Formation of the Isthmus of Panama », Science Advances, vol. 2, no 8, , e1600883 (ISSN 2375-2548, PMID 27540590, PMCID PMC4988774, DOI 10.1126/sciadv.1600883, lire en ligne, consulté le ).