Acide gras
Un acide gras est un acide carboxylique à chaîne aliphatique. Les acides gras naturels possèdent une chaîne carbonée de 4 à 36 atomes de carbone (rarement au-delà de 28[1]) et typiquement en nombre pair, car la biosynthèse des acides gras, catalysée par l'acide gras synthase, procède en ajoutant de façon itérative des groupes de deux atomes de carbone grâce à l'acétyl-CoA[2]. Par extension, le terme est parfois utilisé pour désigner tous les acides carboxyliques à chaîne hydrocarbonée non-cyclique. On parle d'acides gras à longue chaîne pour une longueur de 14 à 24 atomes de carbone et à très longue chaîne s'il y a plus de 24 atomes de carbone. Les acides gras sont présents dans les graisses animales et les graisses végétales, les huiles végétales ou les cires, sous forme d'esters[3].
En biochimie, les acides gras sont une catégorie de lipides qui comprend notamment les acides carboxyliques aliphatiques et leurs dérivés (acides gras méthylés, hydroxylés, hydroperoxylés, etc.) et les eicosanoïdes. Ces derniers dérivent de l'acide eicosapentaénoïque (oméga-3) ou de l'acide arachidonique (oméga-6) et ont souvent un rôle d'hormones. Les acides gras jouent un rôle structurel fondamental dans toutes les formes de vies connues à travers divers types de lipides (phosphoglycérides, sphingolipides…) qui, en milieu aqueux, s'organisent en réseaux bidimensionnels structurant toutes les membranes biologiques (membrane cellulaire, plasmique, mitochondriale, du réticulum endoplasmique, des thylacoïdes, etc.).
Ils constituent également des sources importantes d'énergie métabolique : les acides gras permettent aux êtres vivants de stocker environ 37 kJ d'énergie (9 kcal) par gramme de lipides, contre environ 17 kJ pour les glucides (4 kcal). Ils sont stockés par l'organisme sous forme de triglycérides, dans lesquels trois molécules d'acides gras forment un ester avec un résidu de glycérol. Lorsqu'ils ne sont pas liés à d'autres molécules, les acides gras sont dits « libres ». Leur dégradation produit de grandes quantités d'ATP, molécule énergétique privilégiée des cellules : la β-oxydation suivie de la dégradation par le cycle de Krebs d'un acide gras saturé à n = 2p atomes de carbone libère en effet (10p – 2) ATP + (p – 1) × (FADH2 + NADH+H+), soit l'équivalent énergétique de 106 ATP pour une molécule d'acide palmitique CH3(–CH2)14–COOH, qui contient 16 atomes de carbone (n = 16, et donc p = 8).
Les acides gras peuvent être synthétisés par l'organisme à travers un ensemble de processus métaboliques appelés lipogenèse. Ils sont également apportés en grande quantité par l'alimentation. En moyenne, les besoins énergétiques quotidiens s'élèvent à 2 000 kcal pour une femme et 2 500 kcal pour un homme adulte[4], dont les matières grasses ne devraient idéalement pas représenter plus de 35 %[5], soit 65 g pour une femme et 90 g pour un homme adulte[6]. L'alimentation humaine occidentale apporte néanmoins bien davantage de matières grasses, les valeurs annuelles individuelles pour la France, toutes tranches d'âges confondues, s'élevant en moyenne à 125 g par jour et par personne[7], autrement dit 160 % de la quantité maximale recommandée pour un adulte.
Nomenclature et classification
Nomenclature des acides gras
Il existe plusieurs nomenclatures parallèles pour désigner les acides gras.
- Nom d'usage : les plus communs des acides gras ont des noms d'usage non systématiques sous lesquels ils sont désignés dans la littérature. Ces noms ne suivent généralement aucune règle mais sont concis et non ambigus, désignant un diastéréoisomère précis d'un acide gras donné.
- Exemples : acide palmitique, acide palmitoléique, acide arachidique, acide arachidonique, etc.
- Nom IUPAC : tous les acides gras s'inscrivent également dans la nomenclature des composés organiques publiée en 1979 par l'IUPAC en complément de recommandations destinées spécifiquement à la nomenclature des lipides publiée en 1977[8]. Les atomes de carbone sont comptés à partir de l'extrémité carboxylique –COOH et la diastéréoisomérie des doubles liaisons est indiquée par la notation cis / trans ou par la notation (Z) / (E).
- Exemples : acide cis-9-hexadécénoïque ou acide (Z)-hexadéc-9-énoïque pour l'acide palmitoléique.
- Nomenclature Δx : la nomenclature « delta-x » concerne les acides gras insaturés, pour lesquels chaque double liaison est indiquée par le signe Δ précédé de sa configuration cis ou trans et suivi en exposant par la position de la double liaison le long de la chaîne aliphatique de l'acide gras depuis l'extrémité carboxylique –COOH de la molécule.
- Exemples : acide cis-Δ9-hexadécénoïque pour l'acide palmitoléique ; acide cis-Δ9,cis-Δ12-octadécadiénoïque, acide cis,cis-Δ9Δ12-octadécadiénoïque, acide cis,cis-Δ9,12-octadécadiénoïque, ou encore acide tout-cis-Δ9,12-octadécadiénoïque pour l'acide linoléique.
- Nomenclature biochimique : en biochimie, les acides gras sont souvent désignés par des numéros de la forme C:D dans laquelle C représente le nombre d'atomes de carbone de l'acide et D le nombre de doubles liaisons qu'il contient. Cette désignation est très souvent complétée par une mention de la forme n-x dans laquelle n symbolise le nombre d'atomes de carbone et x la position de la première double liaison comptée depuis l'extrémité méthyle –CH3 de la molécule — c'est-à-dire dans l'autre sens que celui qui prévaut pour la nomenclature IUPAC et la nomenclature Δx. Dans le grand public, la désignation n-x, préconisée par l'IUPAC, est généralement écrite « ω-x » ou « oméga-x ». Cette nomenclature est imprécise, plusieurs acides gras différents pouvant être décrits par les mêmes numéros C:D n-x, mais des acides gras de mêmes numéros auront souvent des propriétés biochimiques ou physiologiques semblables : les oméga-3 et oméga-6 sont ainsi bien connus du public pour leurs effets sur la santé.
- Exemples : 16:1 n-7 pour l'acide palmitoléique ; 18:2 n-6 ou 18:2 ω-6 pour l'acide linoléique, un oméga-6 essentiel ; 18:3 n-6 pour l'acide pinolénique, l'acide γ-linolénique (GLA) ou encore l'acide jacarique, respectivement diastéréoisomères tout-cis-Δ5,9,12, tout-cis-Δ6,9,12 et cis,trans,cis-Δ8,10,12 de l'acide octadécatriénoïque (18:3) — le GLA est en fait le seul des trois à être habituellement rangé parmi les oméga-6.
 Numérotation des atomes de carbone de l'acide stéaridonique, un acide gras oméga-3, selon les nomenclatures chimique (en bleu) et biochimique (en rouge).
Numérotation des atomes de carbone de l'acide stéaridonique, un acide gras oméga-3, selon les nomenclatures chimique (en bleu) et biochimique (en rouge).
Acides gras saturés
| Nom d'usage | Structure | C:D |
|---|---|---|
| Acide caprylique | CH3(–CH2)6–COOH | 8:0 |
| Acide caprique | CH3(–CH2)8–COOH | 10:0 |
| Acide laurique | CH3(–CH2)10–COOH | 12:0 |
| Acide myristique | CH3(–CH2)12–COOH | 14:0 |
| Acide palmitique | CH3(–CH2)14–COOH | 16:0 |
| Acide stéarique | CH3(–CH2)16–COOH | 18:0 |
| Acide arachidique | CH3(–CH2)18–COOH | 20:0 |
| Acide béhénique | CH3(–CH2)20–COOH | 22:0 |
| Acide lignocérique | CH3(–CH2)22–COOH | 24:0 |
| Acide cérotique | CH3(–CH2)24–COOH | 26:0 |
Un acide gras saturé est un acide carboxylique aliphatique comportant typiquement de 12 à 24 atomes de carbone et aucune double liaison carbone-carbone : tous les atomes de carbone sont saturés en hydrogène, la formule semi-développée d'un tel acide gras à n atomes de carbone étant H3C(–CH2)n-2–COOH.
Les molécules de ces composés organiques sont linéaires de longueur variable. Plus les membranes biologiques sont riches en acides gras saturés et plus les lipides qui portent ces acides gras auront tendance à « cristalliser » en formant des réseaux ordonnés dont la fluidité est entravée par de nombreuses liaisons hydrogène ; le taux d'acides gras saturés est un moyen de réguler la fluidité des membranes biologiques en fonction de la température. Un excès d'acides gras saturés dans les membranes cellulaires peut cependant altérer leur fonction d'interfaces biologiques entre le cytoplasme et le milieu extracellulaire, par exemple en réduisant la perméabilité de ces membranes ou en modifiant le comportement de certains récepteurs membranaires.

Acides gras insaturés
Un acide gras insaturé est un acide gras qui comporte une ou plusieurs doubles liaisons carbone-carbone. Ces doubles liaisons peuvent être réduites en liaisons covalentes simples par l'addition de deux atomes d'hydrogène, conduisant à un acide gras saturé. Elles introduisent une isomérie cis-trans au niveau de chacune d'elles, la configuration cis étant privilégiée dans les structures biologiques tandis que la configuration trans demeure assez rare dans le milieu naturel et est généralement le fruit d'une manipulation humaine sur la structure des acides gras.
Acides gras trans
Une configuration trans signifie que les atomes de carbone adjacents à la double liaison sont situés de part et d'autre de cette dernière. La chaîne hydrocarbonée adopte une configuration rectiligne très semblable à celle de l'acide gras saturé correspondant. La plupart des acides gras trans ne sont pas synthétisés naturellement par les organismes vivants et sont produits industriellement par des procédés tels que l'hydrogénation partielle, par exemple dans les procédés de l'industrie agroalimentaire destinés à donner une meilleure tenue aux aliments (viennoiseries plus « croustillantes ») et une plus grande résistance au rancissement (à la peroxydation des lipides en particulier), les acides gras insaturés étant particulièrement sensibles à cet égard.
Acides gras cis
Une configuration cis signifie que les atomes de carbone adjacents à la double liaison sont situés du même côté de cette dernière. Cette configuration introduit une courbure de la chaîne hydrocarbonée de l'acide gras tout en réduisant sa flexibilité mécanique. Plus un acide gras possède de doubles liaisons, qui sont naturellement cis, plus sa chaîne aliphatique est tordue et rigide : avec une double liaison, la molécule d'acide oléique (cis-Δ9 18:1) est coudée en son milieu, tandis que, avec deux doubles liaisons, la molécule d'acide linoléique (tout-cis-Δ9,12 18:2) présente une forme gauchie et, avec trois doubles liaisons, la molécule d'acide α-linolénique (tout-cis-Δ9,12,15 18:3) a une forme en crochet.
Ces formes particulières, associées à la plus grande rigidité de ces molécules, réduisent la faculté de ces dernières à constituer des réseaux de molécules ordonnées interagissant entre elles par liaisons hydrogène, ce qui tend à abaisser la température de fusion des structures biologiques qui contiennent de telles molécules, notamment des membranes, dont la fluidité augmente avec leur taux d'acides gras insaturés.
- Représentations 3D de molécules d'acides gras insaturés naturels


| Nom d'usage | Formule semi-développée | Δx | C:D | n−x | |
|---|---|---|---|---|---|
| Acide myristoléique | CH3(–CH2)3–CH=CH(–CH2)7–COOH | cis-Δ9 | 14:1 | n−5 | |
| Acide palmitoléique | CH3(–CH2)5–CH=CH(–CH2)7–COOH | cis-Δ9 | 16:1 | n−7 | |
| Acide sapiénique | CH3(–CH2)8–CH=CH(–CH2)4–COOH | cis-Δ6 | 16:1 | n−10 | |
| Acide oléique | CH3(–CH2)7–CH=CH(–CH2)7–COOH | cis-Δ9 | 18:1 | n−9 | |
| Acide élaïdique | CH3(–CH2)7–CH=CH(–CH2)7–COOH | trans-Δ9 | 18:1 | n−9 | |
| Acide trans-vaccénique | CH3(–CH2)5–CH=CH(–CH2)9–COOH | trans-Δ11 | 18:1 | n−7 | |
| Acide linoléique | LA | CH3(–CH2)3(–CH2–CH=CH)2(–CH2)7–COOH | tout-cis-Δ9,12 | 18:2 | n−6 |
| Acide linolélaïdique | CH3(–CH2)3(–CH2–CH=CH)2(–CH2)7–COOH | tout-trans-Δ9,12 | 18:2 | n−6 | |
| Acide α-linolénique | ALA | CH3(–CH2–CH=CH)3(–CH2)7–COOH | tout-cis-Δ9,12,15 | 18:3 | n−3 |
| Acide γ-linolénique | GLA | CH3(–CH2)3(–CH2–CH=CH)3(–CH2)4–COOH | tout-cis-Δ6,9,12 | 18:3 | n−6 |
| Acide dihomo-γ-linolénique | DGLA | CH3(–CH2)3(–CH2–CH=CH)3(–CH2)6–COOH | tout-cis-Δ8,11,14 | 20:3 | n−6 |
| Acide arachidonique | AA | CH3(–CH2)3(–CH2–CH=CH)4(–CH2)3–COOHNIST | tout-cis-Δ5,8,11,14 | 20:4 | n−6 |
| Acide eicosapentaénoïque | EPA | CH3(–CH2–CH=CH)5(–CH2)3–COOH | tout-cis-Δ5,8,11,14,17 | 20:5 | n−3 |
| Acide clupanodonique | DPA | CH3(–CH2–CH=CH)5(–CH2)5–COOH | tout-cis-Δ7,10,13,16,19 | 22:5 | n−3 |
| Acide docosahexaénoïque | DHA | CH3(–CH2–CH=CH)6(–CH2)2–COOH | tout-cis-Δ4,7,10,13,16,19 | 22:6 | n−3 |
Acides gras essentiels, oméga-3 et oméga-6
Les acides gras essentiels sont appelés ainsi parce qu'ils ne peuvent être biosynthétisés en quantité suffisante par le corps humain. Parmi ceux-ci, les acides gras oméga-3 et oméga-6 sont des acides gras insaturés dont la première double liaison comptée depuis le méthyle –CH3 terminal se trouve respectivement sur la troisième (ω-3) et sur la sixième (ω-6) liaison carbone-carbone.
Chez l'humain, seuls l'acide α-linolénique (ALA, tout-cis-Δ9,12,15 18:3, un acide gras ω-3) et l'acide linoléique (LA, tout-cis-Δ9,12 18:2, un acide gras ω-6) sont strictement essentiels, car ils ne sont pas synthétisés par l'organisme et doivent par conséquent lui être intégralement fournis par l'alimentation ; les autres acides gras qualifiés d'essentiels le sont en réalité selon les circonstances, car ils peuvent être synthétisés par l'organisme à partir d'autres acides gras, mais en quantité qui peut être insuffisante, d'où la nécessité de pallier ce manque éventuel par l'apport alimentaire :
- Ω-3 : l'acide α-linolénique peut donner de l'acide eicosapentaénoïque (EPA, tout-cis-Δ5,8,11,14,17 20:5), lequel peut donner à son tour de l'acide docosahexaénoïque (DHA, tout-cis-Δ4,7,10,13,16,19 22:6),
- Ω-6 : l'acide linoléique peut donner de l'acide γ-linolénique (tout-cis-Δ6,9,12 18:3), lequel peut donner à son tour de l'acide dihomo-γ-linolénique (tout-cis-Δ8,11,14 20:3), ainsi que de l'acide arachidonique (tout-cis-Δ5,8,11,14 20:4), la synthèse de ce dernier diminuant avec l'âge.
Ces acides gras agissent de manière complexe dans l'organisme. Les métabolites issus des oméga-6 sont pro-inflammatoires, prothrombotiques et hypertenseurs tandis que ceux issus des oméga-3 ont globalement un effet inverse. Le rapport alimentaire optimal entre ces deux classes d'acides gras est généralement estimé entre 1 et 4 fois plus d'oméga-6 que d'oméga-3[9]. Cependant, l'alimentation humaine occidentale apporte en moyenne 16 fois plus d'oméga-6 que d'oméga-3[9], avec des valeurs pouvant dépasser 30 fois dans certains régimes alimentaires particulièrement déséquilibrés. Un excès trop prononcé d'oméga-6 par rapport aux oméga-3 tendrait à favoriser le développement de diverses maladies telles que les maladies cardiovasculaires, des cancers et diverses maladies inflammatoires et auto-immunes.
Eicosanoïdes
Les eicosanoïdes sont des lipides participant à la signalisation cellulaire issus de l'oxydation d'acides gras polyinsaturés à 20 atomes de carbone. Ils agissent de façon complexe sur de nombreux processus physiologiques, essentiellement l'inflammation et le système immunitaire, et comme messagers dans le système nerveux central. Ils dérivent d'acides gras essentiels oméga-3 ou oméga-6 selon les cas. Les eicosanoïdes ω-6 sont généralement pro-inflammatoires, tandis que les eicosanoïdes ω-3 le sont sensiblement moins. L'équilibre entre ces deux types d'eicosanoïdes oriente les fonctions physiologiques gouvernées par ces molécules, un déséquilibre pouvant avoir des effets sur les maladies cardio-vasculaires, le taux sérique de triglycérides, la pression artérielle ou encore l'arthrite. Les anti-inflammatoires non stéroïdiens tels que l'aspirine agissent en réduisant la synthèse des eicosanoïdes.
Il existe quatre familles d'eicosanoïdes — les prostaglandines, les prostacyclines, les thromboxanes et les leucotriènes — ayant chacune deux ou trois séries de composés dérivées d'un acide gras essentiel ω-3 ou ω-6 :
- l'acide eicosapentaénoïque (EPA), un ω-3 à cinq doubles liaisons,
- l'acide arachidonique (AA), un ω-6 à quatre doubles liaisons,
- l'acide dihomo-γ-linolénique, un ω-6 à trois doubles liaisons.
L'activité physiologique de ces différentes séries rend largement compte des effets que peuvent avoir les acides gras ω-3 et ω-6 sur la santé[10] - [11] - [12].
Autres types d'acides gras et dérivés
Certains acides gras insaturés naturels possèdent une ou plusieurs triples liaisons dans leur chaîne carbonée. C'est par exemple le cas de l'acide crépénynique, issu de l'acide oléique par déshydrogénation de la double liaison cis-Δ12 par une enzyme spécifique, la Δ12-acide gras déshydrogénase (EC ).
Si les acides gras les plus courants dans les structures biologiques ont une chaîne aliphatique linéaire, il existe cependant, chez les bactéries, les algues et certaines plantes, ainsi que chez les animaux en petite quantité, des acides gras à chaîne hydrocarbonée ramifiée ou cyclique, tels que l'acide tuberculostéarique (acide 10-méthylstéarique), l'acide phytanique (un terpénoïde), l'acide lactobacillique (cyclopropanique), l'acide malvalique (cyclopropénique), l'acide 11-cyclohexylundécanoïque (terminé par un cyclohexane), l'acide 13-phényltridécanoïque (terminé par un groupe phényle), ou encore l'acide chaulmoogrique (terminé par un cyclobutène). Il existe également des acides gras furaniques, synthétisés par des végétaux mais qui se retrouvent chez un très grand nombre d'êtres vivants, notamment les animaux, chez qui ils jouent un rôle d'antioxydant protecteur contre les radicaux libres. Par ailleurs, certaines éponges des côtes australiennes renferment des acides gras dits « amphimiques » (du nom du genre d'éponges Amphimedon) qui contiennent un groupe méthylènecyclopropane. Enfin, on relèvera l'existence d'acides gras de type ladderane, dont le premier à avoir été découvert est l'acide pentacycloanammoxique.
- Exemples d'acides gras particuliers

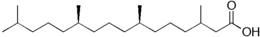
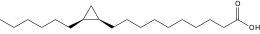
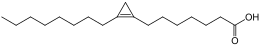
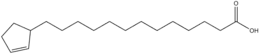
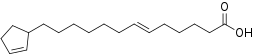
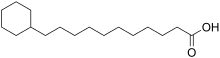
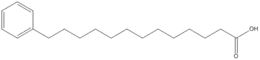
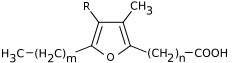
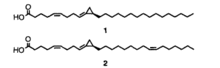 Exemples d'acides amphimiques.
Exemples d'acides amphimiques.
Rôle des acides gras
- Rôle métabolique : les acides gras sont une source d'énergie importante pour l'organisme. Ils sont stockés sous forme de triglycérides dans les tissus adipeux. Lors d'un effort, en particulier lors d'un effort de longue durée, l'organisme va puiser dans ces stocks et dégrader les acides gras afin de produire de l'énergie sous forme d'ATP.
- Rôle structural : les acides gras servent à la synthèse d'autres lipides, notamment les phospholipides qui forment les membranes autour des cellules et des organites. La composition en acides gras de ces phospholipides donne aux membranes des propriétés physiques (élasticité, viscosité) particulières.
- Rôle de messager : les acides gras sont les précurseurs de plusieurs messagers intra- et extracellulaires. Par exemple, l'acide arachidonique est le précurseur des eicosanoïdes, hormones intervenant dans l'inflammation, la coagulation sanguine, etc.
- Autres rôles : les acides gras sont stockés sous forme de triglycérides dans les bosses du chameau et de dromadaire. Leur dégradation amène à la formation de l'eau. De cette manière, les acides gras constituent une réserve d'eau pour ces animaux.
Métabolisme des acides gras
Le métabolisme des acides gras comprend deux voies métaboliques principales :
- La lipogenèse, ou synthèse de novo, qui consiste à synthétiser un acide gras par condensation de molécules d'acide acétique CH3–COOH (ou unités acétyle CH3–CO–) à deux atomes de carbone en consommant de l'énergie sous forme d'ATP et du pouvoir réducteur sous forme de NADPH+H+. Les unités acétyle sont activées sous la forme d'acétyl-coenzyme A.
- La β-oxydation, qui consiste à dégrader des acides gras en unités acétyle afin de libérer de l'énergie métabolique et du pouvoir réducteur sous la forme d'ATP, de NADH+H+ et de FADH2.
Lipogenèse
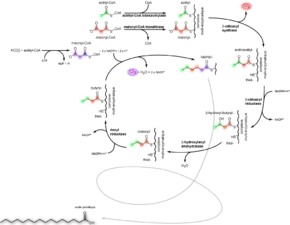
La lipogenèse permet la synthèse d'acides gras saturés par condensation de molécules d'acétate à deux atomes de carbone. Chez les mammifères, ce processus a lieu dans le cytoplasme des cellules, principalement du foie, des tissus adipeux et des glandes mammaires. Cependant, il ne permet pas la biosynthèse des acides gras saturés à plus de 16 atomes de carbone (acide palmitique) ni des acides gras insaturés. L'ensemble de la synthèse est réalisée au niveau d'un complexe multi-enzymatique appelé acide gras synthase[13]. Le bilan de la synthèse de l'acide palmitique est :
- 8 Acétyl-CoA + 7 ATP + 14 NADPH+H+ → Acide palmitique (16:0) + 8 CoA + 7 (ADP + Pi) + 14 NADP+ + 6 H2O.
Cette synthèse est consommatrice d'énergie sous forme d'ATP et nécessite comme cofacteurs de la coenzyme A (CoA) et du nicotinamide adénine dinucléotide phosphate (NADP). La coenzyme A permet de faciliter l'utilisation de l'acétate par la cellule. L'acétyl-CoA provient principalement des mitochondries, où elle est synthétisée à partir du pyruvate par le cycle de Krebs. Le NADP est le réducteur de la synthèse des acides gras. De fait, il est oxydé à la fin de la réaction et doit être régénéré.
L'élongation des acides gras saturés au-delà de 16 atomes de carbone est réalisée dans le réticulum endoplasmique et les mitochondries. Dans le premier cas, l'élongation implique des acides gras élongases. Dans le second cas, l'élongation implique paradoxalement certaines enzymes de la lipolyse.
La synthèse des acides gras insaturés à partir des acides gras saturés a lieu au niveau de la membrane du réticulum endoplasmique par des acides gras désaturases. La désaturation est consommatrice d'oxygène moléculaire O2 et utilise comme cofacteur du nicotinamide adénine dinucléotide (NAD) :
- Acide stéarique (18:0) + 2 NADH+H+ + O2 → Acide oléique (18:1) + NAD+ + 2 H2O.
Tous les organismes ne synthétisent pas nécessairement tous les acides gras saturés et insaturés existants. Ainsi, les humains ne peuvent synthétiser l'acide linoléique et l'acide α-linolénique : ces acides gras sont dits essentiels et doivent être apportés par l'alimentation.
Acides gras, nutrition et maladies cardio-vasculaires
L'alimentation est une source importante d'acides gras. Cet apport est vital pour maintenir une lipidémie stable et pour fournir au corps les acides gras essentiels. Les acides gras qualifiés d'essentiels incluent les oméga-3 et oméga-6. Le corps humain ne sachant pas les synthétiser, ou les synthétisant en quantité insuffisante, un apport minimal et régulier par l'alimentation est nécessaire.
Actuellement, selon l'AFSSA, l'alimentation apporte assez d'oméga-6 et trop peu d'oméga-3, avec un rapport oméga-3 / oméga-6 insuffisant.
En revanche, de nombreuses études ont montré qu'un excès d'acides gras (en particulier les gras insaturés trans) pouvait avoir des conséquences sur la santé et notamment augmenter de façon très significative les risques de maladies cardio-vasculaires[14]. Certaines études portent sur la consommation excessive d'acides gras insaturés trans issus de procédés industriels comme l'hydrogénation partielle d'acides gras d'origine végétale (huiles).
| Acides gras : | saturés | mono- insaturés | poly- insaturés | oméga-3[18] !! oméga-6[18] !! Cholestérol !! Vitamine E | |||
|---|---|---|---|---|---|---|---|
| g / 100 g | g / 100 g | g / 100 g | g / 100 g | g / 100 g | mg / 100 g | mg / 100 g | |
| Graisses animales | |||||||
| Lard | 40,8 | 43,8 | 9,6 | 93 | 0,0 | ||
| Beurre | 54,0 | 19,8 | 2,6 | 230 | 2,0 | ||
| Graisse de canard[19] | 33.2 | 49.3 | 12.9 | 0.101 | |||
| Graisses végétales | |||||||
| Huile de coco | 85,2 | 6,6 | 1,7 | 0 | 0,7 | ||
| Huile de lin | 11 | 10-20 | 52 - 80 | 45 - 70 | 12-24 | 17.5 | |
| Huile de palme | 45,3 | 41,6 | 8,3 | 0 | 33,1 | ||
| Huile de coton | 25,5 | 21,3 | 48,1 | 0 | 42,8 | ||
| Huile de germe de blé | 18,8 | 15,9 | 60,7 | 8 | 53 | 0 | 136,7 |
| Huile de soja | 14,5 | 23,2 | 56,5 | 5 | 50 | 0 | 16,3 |
| Huile d'olive | 14,0 | 69,7 | 11,2 | 0 | 7,5 | 0 | 5,1 |
| Huile de maïs | 12,7 | 24,7 | 57,8 | 0 | 17,4 | ||
| Huile de tournesol | 11,9 | 20,2 | 63,0 | 0 | 62 | 0 | 49,0 |
| Huile de carthame | 10,2 | 12,6 | 72,1 | 0,1-6 | 63-72 | 0 | 40,7 |
| Huile de colza | 5,3 | 64,3 | 21-28 | 6-10 | 21-23 | 0 | 22,2 |
Dans un avis publié en 1992[20], l'American Heart Association (AHA) a fait les recommandations suivantes :
- l'apport énergétique quotidien provenant des matières grasses ne devrait pas dépasser 35 à 40 % de l'apport journalier recommandé;
- ces matières grasses devraient contenir
- 50 % d'acide gras monoinsaturés de type oméga-9
- 25 % d'acide gras polyinsaturés de type oméga-3 et oméga-6
- 25 % d'acides gras saturés ;
- une partie des acides gras saturés peut être remplacée par des acides gras mono-insaturés.
Nb : Pour les analyses réalisées dans le cadre de la répression des fraudes, on détermine l'origine de la matière grasse en fonction du profil en acides gras, et en fonction des stérols (insaponifiables). Les acides gras à chaîne carbonée impaire sont très minoritaires et ne sont souvent pas quantifiés dans les analyses courantes.
Traceurs dans l'écosystème et la chaîne alimentaire
Certaines sortes de lipides (acides gras, alcools gras, hydrocarbures ou stérols) n'étant produits que par certaines plantes, leur suivi (et/ou celui de leurs descendants métaboliques) dans les réseaux trophiques peut permettre d'identifier des sources et puits de certaines matières organiques dans l'alimentation d'un individu, d'une espèce, d'une population et dans les apports de nutriments ou de carbone dans un compartiment d'un écosystème et d'en mieux comprendre certaines dynamiques(Pimm et al., 1991 cités par Napolitano[21]), en complément d'autres moyens d'études par exemple basé sur l'inspection du contenu stomacal, intestinal ou des fèces, sur des marqueurs biochimiques, immunologiques (Grisley et Boyle, 1985, cités par Napolitano[21]) ou des analyses isotopiques (d'isotopes stables naturels ou de marqueurs isotopiques artificiels (Peterson et Fry, 1987 cités par Napolitano[21]). Quand le lipide (acide gras ici) est métaboliquement stable et/ou qu'il conserve sa structure de base après avoir été digéré ou intégré, il peut permettre de tracer les transferts de carbone et d'énergie dans une chaîne alimentaire, et de par exemple affiner la connaissance d'une relation prédateur-proie[21].
Historique
- 1769: découverte du cholestérol dans des calculs biliaires par François Poulletier de La Salle[22];
- 1804: Nicolas Théodore de Saussure réalise une expérience montrant que l'oxygène peut se condenser à l'huile de lin[23]. Ceci est un premier pas vers la découverte des acides gras insaturés;
- 1813 : Eugène Chevreul décrit le concept d'acide gras.
- 1816 : Le même savant expérimentateur est le premier à découvrir et affirmer la structure du savon (sels métalliques d'acides gras).
- 1818 : Eugène Chevreul nomme « cholestérine » le lipide découvert par Poulletier de la Salle;
- 1823 : Eugène Chevreul publie son travail Recherches chimiques sur les corps gras d'origine animale, où il décrit pour la première fois de nombreux acides gras, dont les acides butyrique, caproïque, stéarique et oléique[24];
- 1827: William Prout est le premier à reconnaitre les graisses comme un important nutriment dans l'alimentation, au même titre que les protéines ou les sucres[25];
- 1847 : Théodore Nicolas Gobley isole la lécithine du jaune d'œuf[26]. Il est de fait le découvreur des phospholipides;
- 1869: Hippolyte Mège-Mouriès invente la margarine après que Napoléon III eut lancé un concours afin de trouver un substitut au beurre[27];
- 1903 : Wilhelm Normann dépose un brevet pour la « conversion des acides gras insaturés ou de leurs glycérides en composés saturés » par un procédé d'hydrogénation;
- 1909: découverte de l'acide arachidonique par Percival Hartley[28];
- 1913: McCollum et Davis montrent la nécessité de certains lipides dans l'alimentation lors de la croissance[29];
- 1930: George et Mildred Burr découvrent que certains acides gras polyinsaturés sont essentiels[30];
- 1957: Keys, Anderson et Grande montrent une relation entre le taux de cholestérol sanguin et la prise alimentaire de graisse[31];
- 1964: Konrad Bloch et Feodor Lynen reçoivent le prix Nobel de médecine pour « leur découverte concernant le mécanisme de régulation des métabolismes du cholestérol et des acides gras »[32];
- 1972: Bang et Dyerberg montrent grâce aux Eskimos du Groenland qui ont des taux bas de cholestérol, de LDL et de triglycérides dans le sang que ceux-ci sont corrélés avec un risque bas d'infarctus du myocarde, en comparaison avec la population danoise[33];
- 1982: Sune K. Bergström, Bengt I. Samuelsson et John R. Vane reçoivent le prix Nobel de médecine pour « leur découverte concernant les prostaglandines et les substances biologiquement actives associées »[34];
- 1985: Michael S. Brown et Joseph L. Goldstein reçoivent le prix Nobel de médecine pour « leur découverte portant sur la régulation du métabolisme du cholestérol »[35].
Notes et références
- (en) G.P. MOSS, Glossary of class names of organic compounds and reactivity intermediates based on structure (IUPAC Recommendations 1995), (DOI 10.1351/pac199567081307, lire en ligne), p1335
- Une exception notable à ces assertions est l'acide heptatriacontylique, C37 qui est présent dans plusieurs plantes et animaux.
- (en) D'après l'IUPAC dans le Glossaire basé sur la structure des noms de classes des composés organiques et réactifs intermédiaires
- (en) « What should my daily intake of calories be? », National Health Service, Royaume-Uni.
- (en) « FSA nutrient and food based guidelines for UK institutions », Food Standards Agency, Royaume-Uni.
- « Matières grasses », Ministère de la Santé et des Services sociaux du Québec : recommandation fixant la masse quotidienne de matières grasses consommées à ne pas dépasser par individu.
- exactement 45,6 kg en moyenne entre 2006 et 2008 : « Évolution sur dix ans de la consommation alimentaire : moins de matières grasses animales dans nos assiettes », Ministère de l'Alimentation, de l’Agriculture et de la Pêche, Centre d'études et de prospective, n° 12 - Mars 2010.
- (en) IUPAC-IUB Commission on Biochemical Nomenclature (CBN), « The Nomenclature of Lipids – Recommendations, 1976 », European Journal of Biochemistry, vol. 79, no 1, , p. 11-21 (lire en ligne) DOI 10.1111/j.1432-1033.1977.tb11778.x
- (en) Artemis P. Simopoulos, « The importance of the ratio of omega-6/omega-3 essential fatty acids », Biomedicine & Pharmacotherapy, vol. 56, no 8, , p. 365-379 (lire en ligne) DOI 10.1016/S0753-3322(02)00253-6
- (en) R. De Caterina et G. Basta, « n-3 Fatty acids and the inflammatory response — biological background », European Heart Journal Supplements, vol. 3, no Supplement D, , D42-D49 (lire en ligne) DOI 10.1016/S1520-765X(01)90118-X
- (en) Colin D. Funk, « Prostaglandins and Leukotrienes: Advances in Eicosanoid Biology », Science, vol. 294, no 5548, , p. 1871-1875 (lire en ligne) DOI 10.1126/science.294.5548.1871
- (en) Roy J. Soberman et Peter Christmas, « The organization and consequences of eicosanoid signaling », Journal of Clinical Investigation, vol. 111, no 8, , p. 1107-1113 (lire en ligne) DOI 10.1172/JCI18338
- Maier T., Jenni S. et Ban N., "« Architecture of Mammalian Fatty Acid Synthase at 4.5 A Resolution », Science, vol. 311:1258 - 1262, 2006
- (fr) (en) Autorité Européenne de Sécurité des Aliments, « Avis du groupe scientifique sur les produits diététiques, la nutrition et les allergies sur une question de la Commission relative à la présence d'acides gras trans dans les aliments et aux effets sur la santé humaine de la consommation d'acides gras trans », Question n° EFSA-Q-2003-022, juillet 2004.
- Proposition de classement des sources végétales d'acides gras en fonction de leur profil nutritionnel. Revue OCL.
- (en) Food Standards Agency, « McCance & Widdowson's The Composition of Foods », Fats and Oils, Royal Society of Chemistry, 1991.
- (en) Ted Altar, « More Than You Wanted To Know About Fats/Oils », Sundance Natural Foods Online.
- Les chiffres des deux dernières colonnes proviennent d'une autre source (Huilerie Noël), citée dans le livre « Le cholestérol : un ennemi qui vous veut du bien » de Catherine Martinez. Ils peuvent être considérés comme cohérents avec les autres sources du tableau à l'exception de l'huile de colza ou la somme oméga-3 + oméga-6 (27-33 g/100g) est supérieure aux chiffres de la colonne des polyinsaturés (21-28 g/100g). Les chiffres figurant dans l'article sur l'huile de colza sont respectivement de 28, 6 et 21 g/100g pour les poly-insaturés, les oméga-3 et les oméga-6.
- « Informations Nutritionnelles - Graisse de canard », sur informationsnutritionnelles.fr (consulté le )
- (en) American Heart Association « Guidelines for cardiopulmonary resuscitation and emergency cardiac care », JAMA Vol. 268:2171-302, 1992.
- Napolitano GE (1999) Fatty acids as trophic and chemical markers in freshwater ecosystems. In Lipids in freshwater ecosystems (pp. 21-44). Springer New York. (résumé)
- (en) Olson R.E., « Discovery of the Lipoproteins, Their Role in Fat Transport and Their Significance as Risk Factors », J. Nutr., vol. 128:439S-443S, 1998.
- (fr) de Saussure, N.T., « Recherches chimiques sur la végétation », Paris, 1804
- (fr) Chevreul M.E., « Recherches chimiques sur les corps gras d'origine animale », Levrault F.G. éd., Paris, 1823.
- (en) Prout W., « On the ultimate composition of simple alimentary substances, with some preliminary [remarks on the] analyses of organized bodies in general », Annales de chimie et de physique, vol. 36:366-378, 1827.
- (fr) Gobley N.T., « Recherches chimiques sur le jaune d'œuf - Examen comparatif du jaune d'œuf et de la matière cérébrale », J Pharm Chim, vol. 11:409, 1847.
- (fr) Chauvière F., « Hippolyte mege Mouries et la margarine », Portraits d'inventeurs, émission du 14 mai 2006
- (en) Hartley P., « On the nature of the fat contained in the liver, kidney and heart: Part II », J Physiol, vol. 38:353, 1909
- (en) McCollum E.V. et Davis M., « The necessity of certain lipins in the diet during growth », J Biol Chem, vol. 15:167-175, 1913
- (en) Burr G.O. et Burr M.M., « On the nature and role of the fatty acids essential in nutrition », J Biol Chem, vol. 86:587-621, 1930
- (en) Keys A., Anderson J.T. et Grande F., « Prediction of serum-cholesterol responses of man to changes in fats in the diet », Lancet, vol. 2:959-966, 1957
- (en) nobelprize.org, « The Nobel Prize in Physiology or Medicine 1964 ».
- (en) Bang H.O. et Dyerberg J., « Plasma lipids and lipoproteins in Greenlandic west coast Eskimos », Acta Med Scand., vol. 192:85-94, 1972
- (en) nobelprize.org, « The Nobel Prize in Physiology or Medicine 1982 ».
- (en) nobelprize.org, « The Nobel Prize in Physiology or Medicine 1985 ».
Voir aussi
Articles connexes
Bibliographie
- Cyberlipid.org, « Chronological history of lipid science »
- Robert E. Olson, « Evolution of Ideas about the Nutritional Value of Dietary Fat: Introduction », J. Nutr., vol. 28 (2):421S-422S, 1998
- Lubert Stryer, Jeremy Mark Berg, John L. Tymoczko (trad. Serge Weinman), Biochimie, Flammarion, « Médecine-Sciences », Paris, 2003, 5e éd. (ISBN 978-2-257-17116-0).
Liens externes
- Revue OCL (Oléagineux, Corps gras, lipides)
- Évaluation des substituts actuellement disponibles et des substituts possibles aux huiles et aux graisses partiellement hydrogénées (graisses trans)
- Ressources relatives à la santé :
- FMA
- (en) Medical Subject Headings
- (cs + sk) WikiSkripta
- Notices dans des dictionnaires ou encyclopédies généralistes :