Cycle de Krebs
Le cycle de Krebs, aussi appelé cycle de l'acide citrique par anglicisme, est une voie métabolique présente chez tous les organismes aérobies et dont la fonction première est d'oxyder les groupes acétyle, issus notamment de la dégradation des glucides, des lipides et des protéines, pour en récupérer l'énergie sous forme de huit électrons à haut potentiel de transfert et d'une molécule de GTP ou d'ATP ; les électrons à haut potentiel de transfert, récupérés sur le NADH et l'ubiquinol (CoQ10H2, ou coenzyme Q10 réduite), circulent ensuite à travers la chaîne respiratoire pour former à leur tour des molécules d’ATP supplémentaires par la phosphorylation oxydative.
Ce cycle a été découvert par étapes dans les années 1930, plusieurs de ses éléments ayant été identifiés par le biologiste moléculaire hongrois Albert Szent-Györgyi[1] tandis que son fonctionnement cyclique était mis en évidence par le biochimiste allemand Hans Adolf Krebs en 1937[2]. Il se déroule dans le cytoplasme des procaryotes et dans les mitochondries des eucaryotes. Il s'agit d'un cycle car le dernier métabolite, l'oxaloacétate, est aussi impliqué dans la première réaction. La première étape consiste à transférer le groupe acétyle de l'acétyl-CoA sur de l'oxaloacétate pour former du citrate, lequel a donné son nom au cycle en anglais et dans les langues germaniques. Les étapes suivantes forment une séquence de réactions, catalysées chacune par une enzyme spécifique, qui conduit à l'oxydation progressive du groupe acétyle en deux molécules de dioxyde de carbone (CO2). Ce faisant, ce cycle produit également des précurseurs pour la biosynthèse de certains acides aminés protéinogènes, tandis que le NADH peut être utilisé dans un grand nombre de réactions biochimiques.
Le cycle de Krebs est le point final commun de la dégradation des polyholosides (glycolyse, voie des pentoses phosphates), des lipides (β-oxydation) et des acides aminés, qui aboutissent à la formation d'acétyl-CoA ; ce dernier est une forme de transport des groupes acétyle provenant du pyruvate. Il existe par ailleurs des réactions d'échappement au cycle qui permettent diverses biosynthèses ; le cycle du glyoxylate, absent chez les animaux mais présent notamment chez les plantes, est un exemple parmi d'autres de telles voies métaboliques utilisant le cycle de Krebs.
Le fait que le cycle de Krebs soit indispensable à de nombreuses voies métaboliques, tant anaboliques que cataboliques, suggère qu'il a probablement été l'un des premiers éléments fondateurs du métabolisme cellulaire à s'être mis en place au cours de l'évolution, peut-être par abiogenèse. Il est apparenté à des réactions se produisant chez les bactéries anaérobies et aurait évolué en plusieurs fois[3] ; il existe théoriquement plusieurs alternatives au cycle de Krebs, mais ce dernier semble être le plus efficace : si plusieurs alternatives ont évolué indépendamment, elles ont dû toutes converger vers le cycle de Krebs[4] - [5].

Étapes du cycle de Krebs
Le tableau ci-dessous résume les dix étapes du cycle de Krebs, catalysées par huit enzymes différentes. Ces étapes sont remarquablement conservées selon les espèces, mais les enzymes peuvent en revanche différer assez sensiblement d'un organisme à un autre. Les réactions et les enzymes présentées dans ce tableau sont celles qui prévalent chez les mammifères.
| Substrats | Produits | Enzyme | Type de réaction | Remarques | |
|---|---|---|---|---|---|
| 1 | Oxaloacétate + Acétyl-CoA + H2O |
Citrate + CoA-SH |
Citrate synthase (EC ) |
Crotonisation | Irréversible, allonge l'oxaloacétate (4C) en une molécule à six atomes de carbone |
| 2 | Citrate | cis-Aconitate + H2O |
Aconitase (EC ) |
Déshydratation | Isomérisation réversible |
| 3 | cis-Aconitate + H2O |
Isocitrate | Hydratation | ||
| 4 | Isocitrate + NAD+ |
Oxalosuccinate + NADH + H+ |
Isocitrate déshydrogénase (EC ) |
Oxydation | Produit du NADH (équivalent à 2,5 ATP) |
| 5 | Oxalosuccinate | α-Cétoglutarate + CO2 |
Décarboxylation | Réaction limitante, étape irréversible, produisant une molécule à cinq atomes de carbone. | |
| 6 | α-Cétoglutarate + NAD+ + CoA-SH |
Succinyl-CoA + NADH + H+ + CO2 |
Complexe α-cétoglutarate déshydrogénase (EC ) |
Décarboxylation oxydative |
Étape irréversible, produisant du NADH (équivalent à 2,5 ATP), conduisant à une molécule à quatre atomes de carbone (hors coenzyme A) |
| 7 | Succinyl-CoA + GDP + Pi |
Succinate + CoA-SH + GTP |
Succinyl-CoA synthétase (EC ) |
Phosphorylation | ou ADP → ATP à la place de GDP → GTP, produit une molécule d'ATP ou d'un équivalent La réaction de condensation du GDP avec le Pi et l'hydrolyse de la succinyl-CoA implique la molécule d'H2O requise pour l'équilibre de la réaction. |
| 8 | Succinate + CoQ10 |
Fumarate + CoQ10H2 (ubiquinol) |
Succinate déshydrogénase (EC ) |
Oxydation | Utilise le FAD comme groupe prosthétique (FAD → FADH2 à la première étape de la réaction), équivalent à 1,5 ATP |
| 9 | Fumarate + H2O |
L-Malate | Fumarase (EC ) |
Hydratation | |
| 10 | L-Malate + NAD+ |
Oxaloacétate + NADH + H+ |
Malate déshydrogénase (EC ) |
Oxydation | Réversible (en réalité, l'équilibre favorise la formation du L-malate), produit du NADH (équivalent à 2,5 ATP) |
Synthèse du citrate
La citrate synthase condense l'oxaloacétate et l'acétyl-CoA en citrate avec libération de la coenzyme A. Il se forme un intermédiaire transitoire, la citroyl-CoA. La liaison thioester de l'acétyl-CoA est une liaison à haut potentiel d'hydrolyse. Le couplage des activités hydrolase et synthase rend la réaction de synthèse thermodynamiquement possible. Cette réaction est une étape régulatrice du cycle, avec comme effecteur négatif sur la vitesse de réaction la succinyl-CoA, le NADH, l'acétyl-CoA, le citrate et l'ATP.
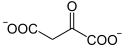
+ acétyl-CoA + H2O → CoA + 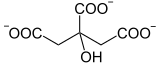
Oxaloacétate Citrate Citrate synthase – EC
Déshydratation du citrate
L'aconitase, une lyase, catalyse la déshydratation du citrate en cis-aconitate. Bien que la molécule de citrate semble être symétrique, on a montré que le départ d'eau a lieu entre les atomes de carbone issus de l'oxaloacétate :

Hydratation du cis-aconitate
L'aconitase catalyse également l'hydratation du cis-aconitate en isocitrate :
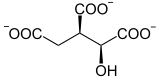
Les deux étapes précédentes, catalysées par l'aconitase, aboutissent à l'isomérisation du citrate en isocitrate :
Oxydation de l’isocitrate
L'isocitrate déshydrogénase, une oxydoréductase, catalyse l'oxydation de l'isocitrate en oxalosuccinate :
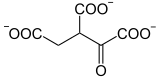
Les eucaryotes utilisent une isocitrate déshydrogénase NAD+-dépendante (EC ), qui exige comme cofacteur des ions Mn2+ ou Mg2+. Les procaryotes utilisent quant à eux une isocitrate déshydrogénase NADP+-dépendante (EC )[6].
Décarboxylation de l'oxalosuccinate
L'isocitrate déshydrogénase catalyse également la décarboxylation de l'oxalosuccinate, instable, en α-cétoglutarate avec dégagement de CO2, en une réaction irréversible :
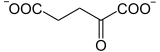
C'est également une étape régulatrice du cycle, avec comme effecteurs négatifs le NADH et l'ATP. La présence d'ADP favorise au contraire l'activité de l'isocitrate déshydrogénase, et ainsi la vitesse de cette réaction.
Décarboxylation oxydative de l'α-cétoglutarate
Le complexe α-cétoglutarate déshydrogénase catalyse la décarboxylation oxydative de l'α-cétoglutarate en succinyl-CoA avec production de NADH+H+ et dégagement de CO2. C'est une réaction semblable à celle convertissant le pyruvate en acétyl-CoA, catalysée par le complexe pyruvate déshydrogénase. Ce complexe enzymatique fait intervenir successivement trois enzymes — l'α-cétoglutarate déshydrogénase, la dihydrolipoamide S-succinyltransférase et la dihydrolipoyl déshydrogénase — et cinq cofacteurs : le TPP, le lipoamide, la coenzyme A, le FAD et le NAD+. Cette réaction est irréversible.
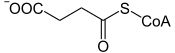
Le NADH, le GTP et la succinyl-CoA sont des effecteurs négatifs sur l'activité du complexe enzymatique.
Formation du succinate
La succinyl-CoA synthétase, ou succinate thiokinase, convertit la succinyl-CoA en succinate et coenzyme A avec formation d'une molécule de GTP chez les animaux ou d'ATP chez les plantes et les bactéries. Cette réaction est réversible.
Oxydation du succinate
La succinate déshydrogénase, une oxydoréductase, catalyse l'oxydation du succinate en fumarate avec réduction concomitante de l'ubiquinone (coenzyme Q10) en ubiquinol (CoQ10H2). Cette enzyme flavoprotéique à FAD est le complexe II de la chaîne respiratoire. Elle est inhibée par le malonate. Le FAD étant un groupe prosthétique lié de manière covalente à l'enzyme, il ne fait que transmettre les électrons et protons au « vrai » substrat CoQ10.
Cette réaction est la quatrième et dernière réaction régulatrice du cycle. Le malonate est ici l'inhibiteur compétitif.
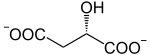
Oxydation du malate : fermeture du cycle
La malate déshydrogénase, une oxydoréductase, convertit le L-malate en oxaloacétate avec formation de NADH+H+.
Cette réaction est catalysée par une malate déshydrogénase NAD+-dépendante (EC ) chez les eucaryotes et quinone-dépendante (EC ) chez les procaryotes[7].
Bilan du cycle de Krebs
Le cycle de Krebs est composé de 10 étapes catalysées par huit enzymes différentes. Au cours du cycle sont produites, à partir d'une mole d'acétate et jusqu'au stade CO2 et H2O :
On constate que le cycle de Krebs ne produit qu'un seul équivalent ATP (une GTP), soit moins que la glycolyse (quatre molécules d'ATP pour une molécule de glucose dont deux sont utilisées lors de la phase « d'activation » de la glycolyse — étapes 1 et 3, qui correspondent à des phosphorylations). L'essentiel de l'énergie chimique potentielle est produite sous forme de pouvoir réducteur (NADH + H+ et CoQ10H2). Ce pouvoir réducteur est ultérieurement utilisé dans la chaîne respiratoire des mitochondries pour produire 11 autres molécules d'ATP via un gradient de protons et une ATP synthase qu'on attribue parfois de façon erronée au cycle de Krebs.
| Description | Substrats | Produits |
| La somme de toutes les réactions d'oxydation de l'acétyl-CoA par le cycle de Krebs (hors chaîne respiratoire) correspond à : | Acétyl-CoA + 3 NAD+ + CoQ10 + GDP + Pi + 2 H2O | → CoA-SH + 3 (NADH + H+) + CoQ10H2 + GTP + 2 CO2 |
| En remontant jusqu'à la décarboxylation du pyruvate, le bilan devient : | Pyruvate + 4 NAD+ + CoQ10 + GDP + Pi + 2 H2O | → 4 (NADH + H+) + CoQ10H2 + GTP + 3 CO2 |
| En remontant jusqu'à l'oxydation du glucose par la glycolyse, le bilan devient : | Glucose + 10 NAD+ + 2 CoQ10 + 2 ADP + 2 GDP + 4 Pi + 2 H2O | → 10 (NADH + H+) + 2 CoQ10H2 + 2 ATP + 2 GTP + 6 CO2 |
Ce qui correspond, au total, pour l'ensemble de la respiration aérobie (glycolyse, cycle de Krebs, réduction des coenzymes NAD+ et CoQ10 par la chaîne respiratoire) entre 30 et 38 ATP pour une molécule de glucose selon les estimations[8], dépendant en partie de la navette mitochondriale ATP-dépendante utilisée pour transporter le NAD+ de la glycolyse.
L’utilisation du glucose par respiration aérobie est plus énergétique que les fermentations.
En présence d'une grande quantité d'acétyl-CoA, le cycle de Krebs peut être débordé, en particulier chez les diabétiques ayant un déficit sévère en insuline ou après un jeûne prolongé (cf. cétoacidose diabétique pour de plus amples détails).
Variations
Bien que le cycle de Krebs soit généralement très conservé entre espèces, on relève des variations importantes dans les enzymes présentes dans les différents taxons. Il existe notamment des différences entre procaryotes et eucaryotes. Ainsi, la conversion du D-thréo-isocitrate en α-cétoglutarate est catalysée par une isocitrate déshydrogénase NAD+-dépendante (EC ) chez les eucaryotes mais NADP+-dépendante (EC ) chez les procaryotes[6]. Il en va de même de la conversion du L-malate en oxaloacétate, catalysée par une malate déshydrogénase NAD+-dépendante (EC ) chez les eucaryotes et quinone-dépendante (EC ) chez les procaryotes[7].
La conversion de la succinyl-CoA en succinate par la succinyl-CoA synthétase présente une variabilité significative. La plupart des êtres vivants utilisent une enzyme ADP-dépendante (EC ), tandis que les mammifères utilisent également l'isoforme GDP-dépendante (EC ) de cette enzyme. Le taux d'utilisation de chacune de ces deux formes de l'enzyme dépend des tissus considérés[9]. Chez certaines bactéries produisant de l'acétate comme Acetobacter aceti (en), c'est une enzyme totalement différente qui catalyse cette réaction, en l'occurrence la succinyl-CoA:acétate CoA-transférase (EC ) : cette enzyme réalise la jonction du métabolisme de l'acide acétique avec le cycle de Krebs chez ces organismes[10]. Certaines bactéries telles que Helicobacter pylori utilisent une enzyme encore différente pour cette réaction, en l'occurrence la 3-oxoacide CoA-transférase (EC )[11].
Il existe une certaine variabilité également à l'étape précédente, c'est-à-dire au niveau de la conversion de l'α-cétoglutarate en succinly-CoA. La plupart des êtres vivants utilisent le complexe α-cétoglutarate déshydrogénase, mais certaines bactéries utilisent l'α-cétoglutarate synthase (EC )[12]. D'autres organismes, parmi lesquels des bactéries et des archées autotrophes et méthanotrophes obligatoires, court-circuitent complètement la succinyl-CoA et convertissent l'α-cétoglutarate en succinate via le semialdéhyde succinique sous l'action successivement de l'α-cétoglutarate décarboxylase (EC ) et de la succinate-semialdéhyde déshydrogénase (EC )[13].
Régulation
Les étapes irréversibles du cycle de Krebs peuvent être régulées : étape de la citrate synthase, de l'isocitrate déshydrogénase et du complexe α-cétoglutarate déshydrogénase :
- la citrate synthase est activée par l'ADP mais inhibée par le NADH, l'ATP et le citrate. Elle est donc respectivement inhibée par le pouvoir réducteur, la charge énergétique et le produit de la réaction qu'elle catalyse ;
- l'isocitrate déshydrogénase est activée par le calcium et l'ADP, et est inhibée par le NADH et l'ATP ;
- le complexe α-cétoglutarate déshydrogénase est activé par le calcium et inhibé par le NADH, l'ATP et son produit de réaction, la succinyl-CoA.
La régulation du cycle de Krebs repose essentiellement sur la disponibilité du substrat et l'inhibition par les produits de réaction. Si ces réactions n'étaient pas régulées, le cycle de Krebs gaspillerait de grandes quantités d'énergie métabolique en produisant un excès d'ATP et de coenzymes réduites telles que le NADH.
- Le NADH est produit par toutes les déshydrogénases du cycle hormis la succinate déshydrogénase et a pour effet d'inhiber le complexe pyruvate déshydrogénase, l'isocitrate déshydrogénase, le complexe α-cétoglutarate déshydrogénase ainsi que la citrate synthase.
- L'acétyl-CoA inhibe le complexe pyruvate déshydrogénase, tandis que la succinyl-CoA inhibe le complexe α-cétoglutarate déshydrogénase et la citrate synthase. Les essais in vitro sur les enzymes du cycle de Krebs montrent que l'ATP inhibe la citrate synthase et le complexe α-cétoglutarate déshydrogénase ; cependant, le taux d'ATP in vivo ne varie pas de plus de 10 % entre les périodes de repos et d'activité soutenue, et aucun mécanisme allostérique connu ne permet de moduler significativement l'activité d'une enzyme avec une variation d'au plus 10 % de la concentration d'un effecteur allostérique.
- Le cation de calcium Ca2+ intervient comme régulateur. Le taux de calcium dans la matrice mitochondriale peut atteindre quelques dizaines de micromoles par litre[14]. Il active la pyruvate déshydrogénase phosphatase, laquelle active à son tour la composante pyruvate déshydrogénase (E1) du complexe pyruvate déshydrogénase. Il active également l'isocitrate déshydrogénase et le complexe α-cétoglutarate déshydrogénase[15]. Ceci accroît la vitesse de réaction de plusieurs étapes du cycle, et accroît donc le flux de composés à travers cette voie métabolique.
- Le citrate agit en inhibant la phosphofructokinase, laquelle intervient dans la glycolyse et donc contribue à la formation de pyruvate via le fructose-1,6-bisphosphate. Ceci constitue une boucle de rétroaction entre les deux voies métaboliques.
Des travaux publiés en 2007 ont montré un lien important entre les intermédiaires du cycle de Krebs et la régulation des facteurs induits par l'hypoxie (HIF). Ces derniers jouent un rôle dans l'homéostasie de l'oxygène comme facteurs de transcriptions intervenant dans l'angiogenèse, le remodelage vasculaire, la mobilisation du glucose, le transport des ions et l'apoptose. Les HIF sont synthétisés de façon constitutive et l'hydroxylation d'au moins l'un de deux résidus de proline intervient dans leur interaction avec le complexe ubiquitine ligase, ce qui les désigne comme cible pour une dégradation rapide. Cette réaction est catalysée par des procollagène-proline dioxygénases. Le fumarate et le succinate sont des inhibiteurs efficaces de ces enzymes, ce qui pourrait stabiliser les HIF[16].
Intégration avec les grandes voies métaboliques
Voies anaplérotiques et cataplérotiques
Plusieurs voies métaboliques convergent vers le cycle de Krebs. La plupart de ces réactions produisent des métabolites participant au cycle et sont par conséquent des réactions anaplérotiques ; les processus qui, en revanche, consomment des métabolites du cycle de Krebs sont dites cataplérotiques.
Tous les intermédiaires du cycle de Krebs — comme le citrate, l'isocitrate, l'α-cétoglutarate, le succinate, le fumarate, le L-malate et l'oxaloacétate — sont régénérés à chaque tour du cycle. Le fait d'accroître la quantité disponible de l'un de ces métabolites a pour effet d'accroître la quantité totale de tous les métabolites du cycle, dans la mesure où chaque intermédiaire est converti de proche en proche en tous les autres intermédiaires du cycle. C'est la raison pour laquelle le fait de produire l'un quelconque des métabolites du cycle de Krebs a un effet anaplérotique général sur ce cycle, tandis que le fait d'en consommer un métabolite quelconque a un effet cataplérotique également sur l'ensemble du cycle.
Les molécules de pyruvate issues de la glycolyse sont transportées activement depuis le cytosol vers la matrice mitochondriale à travers la membrane mitochondriale interne. Une fois dans la matrice, elles peuvent être oxydées et réagir avec la coenzyme A pour former du CO2, de l'acétyl-CoA et du NADH, ou elles peuvent être carboxylées par la pyruvate carboxylase pour former de l'oxaloacétate. Il s'agit d'une réaction anaplérotique permettant d'accroître le flux et donc le débit traversant le cycle de Krebs lorsque la cellule est confrontée à un besoin accru d'énergie métabolique, par exemple dans les myocytes.
Alimentation du cycle par l'acétyl-CoA
L'acétyl-CoA, en revanche, qui dérive de l'oxydation du pyruvate ou de la β-oxydation des acides gras, n'est jamais régénéré par le cycle de Krebs : au contraire, chaque tour du cycle consomme une molécule d'acétyl-CoA par molécule d'oxaloacétate de la matrice mitochondriale, tandis que le résidu acétyle de l'acétyl-CoA est entièrement oxydé en CO2 et en H2O à travers la chaîne respiratoire, permettant d'en récupérer l'énergie métabolique sous forme d'ATP par phosphorylation oxydative.
Les voies métaboliques qui produisent ou consomment de l'acétyl-CoA ne sont donc pas anaplérotiques ou cataplérotiques pour le cycle de Krebs.
Néoglucogenèse
Dans le foie, la carboxylation du pyruvate cytosolique en oxaloacétate mitochondrial est une étape avancée de la néoglucogenèse, laquelle permet de convertir le lactate et l'alanine désaminée en glucose sous l'effet d'un taux élevé de glucagon et/ou d'adrénaline dans le sang. Dans ces conditions, la formation d'oxaloacétate dans les mitochondries ne présente pas d'effet anaplérotique car du L-malate mitochondrial est consommé pour former de l'oxaloacétate cytosolique, lequel est converti en glucose.
Dégradation des protéines
Dans le processus de dégradation des protéines, les chaînes polypeptidiques sont clivées par des peptidases en les acides aminés qui les constituent. Leur chaîne carbonée peut alors :
- entrer dans le cycle de Krebs — comme c'est par exemple le cas du glutamate et de la glutamine qui entrent sous forme d'α-cétoglutarate — ce qui présente un effet anaplérotique — on parle d'acides aminés glucoformateurs car ils peuvent être orientés vers la néoglucogenèse via le L-malate ;
- être convertie en acétyl-CoA pour être oxydée et produire de l'énergie, comme c'est le cas de la leucine, de l'isoleucine, de la lysine, de la phénylalanine, du tryptophane et de la tyrosine ;
- être convertie en corps cétoniques, susceptibles d'être ou bien oxydés par d'autres tissus que le foie où ils sont produits, ou bien excrétés par voie urinaire ou par la respiration — on parle d'acides aminés cétoformateurs.
Dégradation des lipides
Dans le processus de lipolyse, les triglycérides sont hydrolysés en glycérol et en acides gras. Dans le foie, le glycérol peut être converti en glucose via la dihydroxyacétone phosphate et le glycéraldéhyde-3-phosphate dans le cadre de la néoglucogenèse. Dans de nombreux tissus, notamment le cœur et les muscles squelettiques, les acides gras sont dégradés à travers la β-oxydation, qui produit de l'acétyl-CoA mitochondriale susceptible de rejoindre le cycle de Krebs. Les acides gras à nombre impair d'atomes de carbone produisent de la propionyl-CoA, laquelle est convertie en succinyl-CoA et rejoint le cycle de Krebs comme intermédiaire anaplérotique[17].
Métabolites utilisés comme substrats pour des biosynthèses
Plusieurs métabolites du cycle de Krebs interviennent dans la biosynthèse de composés importants, ce qui présente un effet cataplérotique significatif sur le cycle.
Production de glucides et de lipides
L'acétyl-CoA ne pouvant quitter tel quel les mitochondries, c'est le citrate qui est transporté à travers la membrane mitochondriale interne depuis la matrice mitochondriale vers le cytosol, où il est clivé en acétyl-CoA et oxaloacétate par l'ATP citrate lyase. L'oxaloacétate peut être utilisé pour la néoglucogenèse dans le foie, ou être converti en L-malate et rejoindre les mitochondries. L'acétyl-CoA cytosolique est utilisé pour la biosynthèse des acides gras et du cholestérol. Ce dernier peut, à son tour, être utilisé pour produire des hormones stéroïdiennes, des acides biliaires et de la vitamine D.
Lors de la néoglucogenèse, l'oxaloacétate mitochondrial est réduit en malate, qui est par la suite transporté hors de la mitochondrie pour être à nouveau oxydé en oxaloacétate dans le cytosol. Ce dernier est alors décarboxylé en phosphoénolpyruvate par la phosphoénolpyruvate carboxykinase, qui est l'étape cinétiquement déterminante de la conversion en glucose de pratiquement tous les précurseurs glucoformateurs — tels que les acides aminés glucoformateurs et le lactate — par le foie et les reins.
Production d'acides aminés
La chaîne carbonée de nombreux acides aminés non essentiels provient de métabolites du cycle de Krebs. L'aspartate et l'asparagine dérivent ainsi de l'oxaloacétate, tandis que la glutamine, la proline et l'arginine dérivent de l'α-cétoglutarate. Le groupe amine provient du glutamate par transamination sur un α-cétoacide à l'aide d'enzymes utilisant du phosphate de pyridoxal comme cofacteur ; au cours de ces réactions, le glutamate est converti en α-cétoglutarate, qui est un métabolite du cycle de Krebs.
Production de bases nucléiques
L'aspartate et la glutamine interviennent par ailleurs dans la biosynthèse des bases nucléiques puriques entrant dans la composition des acides nucléiques — ADN et ARN — ainsi que des nucléotides tels que l'ATP, l'AMP cyclique, le NAD, le FAD et la coenzyme A. Les bases nucléiques pyrimidiques, quant à elles, sont dérivées de l'aspartate, lui-même issu de l'oxaloacétate.
Production de porphyrines
La plupart des atomes de carbone des porphyrines proviennent du succinyl-CoA, un métabolite du cycle du Krebs. Les porphyrines constituent les groupes prosthétiques des hémoprotéines, telles que l'hémoglobine, la myoglobine et les cytochromes.
Notes et références
- (en) « The Nobel Prize in Physiology or Medicine 1937 : Albert Szent-Györgyi », (consulté le ) : « The Nobel Prize in Physiology or Medicine 1937 was awarded to Albert Szent-Györgyi "for his discoveries in connection with the biological combustion processes, with special reference to vitamin C and the catalysis of fumaric acid". ».
- (en) « The Nobel Prize in Physiology or Medicine 1953 : Hans Krebs, Fritz Lipmann » (consulté le ) : « The Nobel Prize in Physiology or Medicine 1953 was divided equally between Hans Adolf Krebs "for his discovery of the citric acid cycle" and Fritz Albert Lipmann "for his discovery of co-enzyme A and its importance for intermediary metabolism". ».
- (en) H. Gest, « Evolutionary roots of the citric acid cycle in prokaryotes », Biochemical Society Symposium, vol. 54, , p. 3-16 (PMID 3332996)
- (en) Enrique Meléndez-Hevia, Thomas G. Waddell et Marta Cascante, « The puzzle of the Krebs citric acid cycle: Assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution », Journal of Molecular Evolution, vol. 43, no 1, , p. 293-303 (PMID 8703096, DOI 10.1007/BF02338838, lire en ligne)
- (en) Oliver Ebenhöh et Reinhart Heinrich, « Evolutionary Optimization of Metabolic Pathways. Theoretical Reconstruction of the Stoichiometry of ATP and NADH Producing Systems », Bulletin of Mathematical Biology, vol. 63, no 1, , p. 21-55 (PMID 11146883, DOI 10.1006/bulm.2000.0197, lire en ligne)
- (en) Takehiko SAHARA, Yasuhiro TAKADA, Yoji TAKEUCHI, Naoto YAMAOKA et Noriyuki FUKUNAGA, « Cloning, Sequencing, and Expression of a Gene Encoding the Monomeric Isocitrate Dehydrogenase of the Nitrogen-fixing Bacterium, Azotobacter vinelandii », Bioscience, Biotechnology, and Biochemistry, vol. 66, no 3, , p. 489-500 (PMID 12005040, DOI 10.1271/bbb.66.489, lire en ligne)
- (en) Michel E. van der Rest, Christian Frank et Douwe Molenaar, « Functions of the Membrane-Associated and Cytoplasmic Malate Dehydrogenases in the Citric Acid Cycle of Escherichia coli », Journal of bacteriology, vol. 182, no 24, , p. 6892-6899 (PMID 11092847, PMCID 94812, DOI 10.1128/JB.182.24.6892-6899.2000, lire en ligne)
- (en) R. K. Porter et M. D. Brand, « Mitochondrial proton conductance and H+/O ratio are independent of electron transport rate in isolated hepatocytes », Biochemical Journal, vol. 310, , p. 379–382 (lire en ligne)
- (en) David O. Lambeth, Kristin N. Tews, Steven Adkins, Dean Frohlich et Barry I. Milavetz, « Expression of Two Succinyl-CoA Synthetases with Different Nucleotide Specificities in Mammalian Tissues », Journal of Biological Chemistry, vol. 279, no 35, , p. 36621-36624 (PMID 15234968, DOI 10.1074/jbc.M406884200, lire en ligne)
- (en) Elwood A. Mullins, Julie A. Francois et T. Joseph Kappock, « A Specialized Citric Acid Cycle Requiring Succinyl-Coenzyme A (CoA):Acetate CoA-Transferase (AarC) Confers Acetic Acid Resistance on the Acidophile Acetobacter aceti », Journal of Bacteriology, vol. 190, no 14, , p. 4933-4940 (PMID 18502856, PMCID 2447011, DOI 10.1128/JB.00405-08, lire en ligne)
- (en) Irène E. Corthésy-Theulaz, Gabriela E. Bergonzelli, Hughes Henry, Daniel Bachmann, Daniel F. Schorderet, André L. Blum et L. Nicholas Ornston, « Cloning and Characterization of Helicobacter pylori Succinyl CoA:Acetoacetate CoA-transferase, a Novel Prokaryotic Member of the CoA-transferase Family », Journal of Biological Chemistry, vol. 272, no 41, , p. 25659-25667 (PMID 9325289, DOI 10.1074/jbc.272.41.25659, lire en ligne)
- (en) Anthony D. Baughn, Scott J. Garforth, Catherine Vilchèze et William R. Jacobs Jr., « An Anaerobic-Type α-Ketoglutarate Ferredoxin Oxidoreductase Completes the Oxidative Tricarboxylic Acid Cycle of Mycobacterium tuberculosis », PLoS Pathogens, vol. 5, no 11, , e1000662 (PMID 19936047, PMCID 2773412, DOI 10.1371/journal.ppat.1000662, lire en ligne)
- (en) Shuyi Zhang et Donald A. Bryant, « The Tricarboxylic Acid Cycle in Cyanobacteria », Science, vol. 334, no 6062, , p. 1551-1553 (PMID 22174252, DOI 10.1126/science.1210858, lire en ligne)
- (en) Maxim V. Ivannikov et Gregory T. Macleod, « Mitochondrial Free Ca2+ Levels and Their Effects on Energy Metabolism in Drosophila Motor Nerve Terminals », Biophysical Journal, vol. 104, no 11, , p. 2353-2361 (PMID 23746507, PMCID 3672877, DOI 10.1016/j.bpj.2013.03.064, lire en ligne)
- (en) Richard M. Denton , Philip J. Randle, Barbara J. Bridges, Ronald H. Cooper, Alan L. Kerbey, Helen T. Pask, David L. Severson, David Stansbie et Susan Whitehouse, « Regulation of mammalian pyruvate dehydrogenase », Molecular and Cellular Biochemistry, vol. 9, no 1, , p. 27-53 (PMID 171557, DOI 10.1007/BF01731731, lire en ligne)
- (en) Inhibition of Hypoxia-inducible Factor (HIF) Hydroxylases by Citric Acid Cycle Intermediates. POSSIBLE LINKS BETWEEN CELL METABOLISM AND STABILIZATION OF HIF, « Peppi Koivunen, Maija Hirsilä, Anne M. Remes, Ilmo E. Hassinen, Kari I. Kivirikko et Johanna Myllyharju », Journal of Biological Chemistry, vol. 282, no 7, , p. 4524-4532 (PMID 17182618, DOI 10.1074/jbc.M610415200, lire en ligne)
- (en) Premjit P. Halarnkar et Gary J. Blomquist, « Comparative aspects of propionate metabolism », Comparative Biochemistry and Physiology Part B: Comparative Biochemistry, vol. 92, no 2, , p. 227-231 (PMID 2647392, DOI 10.1016/0305-0491(89)90270-8, lire en ligne)
Voir aussi
Articles connexes
- Chaîne respiratoire : le devenir des NADH,H+
- Cycle de l'urée
- Respiration cellulaire
- Glycolyse
- Les principales voies du métabolisme (sur wikibooks)
- Hans Adolf Krebs
- Cycle de Calvin, Chloroplaste : réactions biochimiques de la photosynthèse