Leucine
La leucine (abréviations IUPAC-IUBMB : Leu et L), du grec ancien λευκός (blanc, éclatant[5])[6], est un acide α-aminé dont l'énantiomère L est l'un des 22 acides aminés protéinogènes, et l'un des 9 acides aminés essentiels pour l'humain. Elle fait partie des acides aminés branchés (BCAA branched-chain amino acid) avec la valine et l’isoleucine.
| Leucine | |
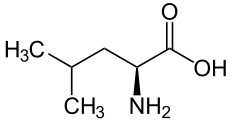 L ou S(+)-leucine 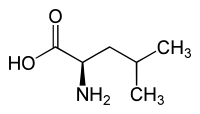 D ou R(–)-leucine |
|
| Identification | |
|---|---|
| Nom UICPA | Acide 2-amino-4-méthylpentanoïque |
| Synonymes |
L, Leu |
| No CAS | (racémique) L ou S(+) D ou R(–) |
| No ECHA | 100.000.475 |
| No CE | 200-522-0 L 206-327-7 D |
| No E | E641 L |
| FEMA | 3297 L |
| SMILES | |
| InChI | |
| Propriétés chimiques | |
| Formule | C6H13NO2 [Isomères] |
| Masse molaire[1] | 131,172 9 ± 0,006 5 g/mol C 54,94 %, H 9,99 %, N 10,68 %, O 24,39 %, |
| pKa | 2,4 et 9,6[2] |
| Propriétés biochimiques | |
| Codons | UUA, UUG, CUU, CUC, CUA, CUG |
| pH isoélectrique | 5,98[2] |
| Acide aminé essentiel | oui |
| Occurrence chez les vertébrés | 7,6 %[3] |
| Précautions | |
| SIMDUT[4] | |
Produit non contrôlé |
|
| Unités du SI et CNTP, sauf indication contraire. | |
Elle est encodée sur les ARN messagers par les codons UUA, UUG, CUU, CUC, CUA et CUG.
Une fois dans la cellule, la leucine peut[7] :
- Réguler les processus cellulaires, notamment stimuler la synthèse protéique musculaire, en activant le complexe 1 de mTOR (mTORC1), une protéine kinase ;
- Être incorporée dans une protéine (protéinogenèse) ;
- Être dégradée par transfert de son groupe amine au α-cétoglutarate, suivant une réaction de transamination.
Les produits finaux du métabolisme de la leucine dans cette troisième voie sont l’acétyl-CoA et l’acétoacétate. Elle fait donc partie des acides aminés exclusivement cétoformateurs, avec la lysine[8].
La L-leucine a une saveur sucrée (son seuil de détection est de 11–13 mmol·l-1) et est utilisée dans l'alimentation en Europe comme exhausteur de goût pour renforcer le goût d'une denrée alimentaire. C'est un additif alimentaire répertorié sous le numéro E641.
Sources alimentaires
L’institut Food and Nutrition Board a calculé un AJR (Apport journalier recommandé) de 42 mg/kg corps/jr pour un adulte de plus de 19 ans[9], soit 2,94 g/jr pour un individu de 70 kg.
Les protéines d’origine animale sont en général plus riches en leucine que les protéines végétales. Elles ont en général un contenu en leucine de 8,5 à 9 % et même plus de 10 % pour les produits laitiers alors que la plupart des protéines végétales en contiennent 6 à 8 %(van Vliet et al[10], 2015).
| Leucine en % du total des protéines (d’après van Vliet et al[10]) | |||||
| Plantes | maïs | spiruline | riz | soja | avoine |
| 12,2 | 8,5 | 8,2 | 8 | 7,7 | |
| Animal | petit-lait | lait | bœuf | œuf | morue |
| 13,6 | 10,9 | 8,8 | 8,5 | 8,1 | |
La base de données des compositions alimentaires (USDA Food Composition Database[11]) donne le contenu en leucine de 4 975 aliments. Nous en extrayons le tableau suivant :
| Aliment pour 100 g | Leucine contenu en g | Leucine contenu en % des protéines tot. |
| Œuf, blanc, en poudre | 7,17 | 8,49 % |
| Soja, isolat de protéines | 6,78 | 7,67 % |
| Morue de l’Atlantique, salée | 5,11 | 8,13 % |
| Spiruline, en poudre | 4,95 | 8,61 % |
| Gruyère | 3,1 | 10,40 % |
| Escalope de veau | 2,88 | 7,96 % |
| Lait entier, en poudre | 2,58 | 9,80 % |
| Germe de blé | 1,57 | 6,78 % |
| Protéine de lactosérum (whey) | 1,19 | 9,2 % |
| Yaourt entier, nature | 0,58 | 16,71 % |
.
La base USDA contient 178 produits commerciaux à base de lactosérum (whey), intitulés « Whey protein bar », « Pure whey protein shake », « Pure whey protein powder» etc. Nous avons retenu le n° 01115 « Whey, sweet, dried » qui donne une analyse en leucine qui est toutefois bien inférieure à celle donnée par van Vliet et al[10].
La colonne nommée « leucine (contenu en g) » est utile pour calculer l'apport en leucine d'un aliment. La colonne (% des protéines tot.) suivante donne une indication sur le meilleur assemblage d'acides aminés au sein d'une protéine conduisant à une moins grande conversion en urée et donc à une plus grande protéosynthèse. Car comme l'ont établi Tujioka et al.[12] (2011), la synthèse des protéines dans les tissus est en partie liée à la concentration relative des acides aminés.
Propriétés anaboliques
Dans les années 1970-80, plusieurs laboratoires ont montré que les acides aminés pouvaient stimuler la synthèse protéique musculaire [13] et inhiber la protéolyse. Ils ont alors pu mettre en évidence le rôle central joué par la leucine, dans ce processus de contrôle de l'équilibre protéosynthèse/protéolyse (voir l’étude classique de Buse et Reid[14], 1975).
Les chercheurs qui ont étudié l’effet de la prise de protéines possédant des taux de leucine plus ou moins élevés sur la protéosynthèse musculaire en ont conclu que c’était le contenu en leucine des protéines qui était le facteur déterminant. Ainsi Tang et al 2009[15], ont comparé trois groupes de jeunes hommes auxquels ils ont fait faire des exercices de résistance[n 1] suivis d’une prise d’une boisson possédant le même contenu en acides aminés essentiels (10 g) sous forme d’hydrolat de petit-lait, de caséine ou de protéines de soja. L’ingestion de protéines de petit-lait s’est traduite par une concentration plasmatique en leucine plus élevée et une stimulation de la synthèse protéique musculaire nettement plus grande, à savoir 33 % fois supérieure par rapport aux protéines de soja et 122 % fois supérieure par rapport à la caséine.
Toutefois la synthèse protéique musculaire à un moment donné n’est qu’un indicateur d’un potentiel de remodelage des muscles. Un gain de masse musculaire ne peut se faire que sur le long terme, de nombreux facteurs pouvant aussi intervenir sur l’importance relative de la protéosynthèse vis-à-vis de la protéolyse. Il a ainsi été montré que la supplémentation sur le long terme de leucine n’accroit pas la masse musculaire des personnes âgées[16].
Mais en 2013, Volek et al.[17] ont pu montrer que la qualité des protéines (caractérisée par un bon assemblage d’acides aminés) associée à des entraînements physiques en résistance jouaient un rôle déterminant. Ils ont comparé trois groupes de personnes prenant respectivement des suppléments iso-caloriques de protéines de petit-lait, de soja ou de glucides après des exercices de résistance et ceci durant 9 mois. Le gain de masse maigre fut plus important pour le petit-lait (3,3 ± 1,5 kg) que pour les glucides (2,3 ± 1,7 kg) et le soja (1,8 ± 1,6 kg). La masse grasse décrut légèrement pour les trois groupes sans différences notoires. Ainsi les trois groupes eurent des résultats contrastés malgré un apport calorique et protéique semblable : les protéines de petit-lait plus riches en leucine et s’absorbant plus rapidement, accroissent plus fortement la protéosynthèse musculaire. L’activité de musculation vient renforcer l’activité de protéosynthèse[18] et en améliorant la sensibilité à l'insuline, elle ralentit la protéolyse. Une étude portant sur des personnes âgées (74± 1 ans) a pu établir que les protéines de lactosérum stimulaient plus efficacement l'accrétion[n 2] protéique que celles de la caséine.
Les muscles ne sont pas les seuls tissus à répondre à une administration orale de leucine. La synthèse protéique s’accroît aussi dans les tissus adipeux blancs, le foie, le cœur, les reins et le pancréas[19].
Mécanismes de régulation moléculaires
La digestion des aliments protéinés libère dans l’intestin des acides aminés libres. Après avoir traversé les entérocytes de l’épithélium intestinal, ils se dirigent vers le foie par la veine porte. Mais contrairement aux acides aminés non branchés, la leucine n’y est pas métabolisée et passe ainsi dans la circulation générale. Cette caractéristique est unique chez les acides aminés. Aussi, après un repas, la concentration plasmique en leucine augmente. La leucine est alors transportée à travers les membranes cellulaires par une famille de transporteurs d’acides aminés de type L[7](nommés LAT1, LAT2, LAT3, LAT4). Il existe aussi une synthèse de novo (endogène) d'acides aminés mais ce n’est pas le cas de la leucine puisque c’est un acide aminé essentiel.
Malgré le caractère obligatoire de l’apport exogène d’acides aminés sous forme d’aliments protéinés, il faut se souvenir que la source principale d’acides aminés pour l’organisme est la protéolyse (ou catabolisme protéique) qui représente 75 % de l’apport total[20]. La dégradation intracellulaire des protéines (anormales ou détériorées) peut se faire dans les lysosomes ou les protéasomes. Ce sont eux qui produisent les trois quarts des acides aminés. Il ne faut pas confondre cette dégradation des protéines (protéolyse) avec la dégradation des acides aminés (ou transamination) que nous allons voir ci-dessous.
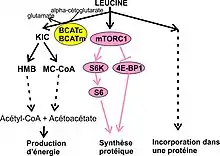
Une fois dans le cytosol de la cellule, la leucine peut[7] :
- Être catabolisée : le transfert du groupe α-aminé de la leucine à l’α-cétoglutarate pour former du glutamate et de l’α-cétoisocarproate (KIC) est catalysée par BCATm et BCATc. Environ 20 % de la leucine est convertie en KIC[7]. Cette étape est appelée la transamination ;
- Réguler les processus cellulaires, notamment stimuler la synthèse protéique, en activant la voie de signalisation mTORC1 ;
- Être incorporée dans une protéine lors de la traduction de l'ARN messager par les ribosomes.
Le démarrage de la dégradation de la leucine se fait principalement dans les mitochondries des muscles squelettiques et d’autres tissus[7]. L’enzyme BCATm s’exprime dans la plupart des tissus sauf les hépatocytes du foie. La BCATc par contre, s’exprime essentiellement dans le système nerveux. Les métabolites de cette voie donnent de l'acétyl-CoA et de l’acétoacétate, utilisés pour la production d’énergie[7] (sous forme d’ATP). La transamination de la leucine par BCATm permet de réguler l’apport de leucine demandé par ses autres fonctions (synthèse protéique et voie mTORC1). Elle permet aussi de prévenir les excès de concentration en leucine qui peuvent être très dommageables; ils peuvent en effet se manifester par une progression tumorale comme l'a montré une étude de 2014 sur des souris, supplémentées en leucine et souffrant d’un cancer du pancréas[21]. Chez les patients cancéreux souffrant de cachexie, la supplémentation de leucine renforce la progression de la tumeur[7].
Régulation de mTORC1 par la leucine et les acides aminés
Le mécanisme moléculaire de la stimulation de la synthèse protéique se joue au niveau d’un complexe enzymatique nommé mTOR[n 3], à activité sérine-thréonine kinase, découvert en 1994[22]. Lorsque la kinase mTOR est associée à d’autres protéines (dont la protéine RAPTOR), elle forme le complexe mTORC1, qui fonctionne comme l’intégrateur des signaux de disponibilité des nutriments, de l'énergie ou du potentiel redox et de contrôle de la protéosynthèse. Pour être actif ce complexe doit se localiser à la surface du lysosome où se trouve son coactivateur Rheb. Une fois activé, le complexe mTORC1 inhibe l’autophagie et augmente la synthèse protéique et la croissance cellulaire.
Le complexe mTORC1 phosphoryle de nombreux substrats, en particulier ceux qui sont impliqués dans la traduction des ARN messager en protéines. La dérégulation de la voie mTOR caractérise des maladies telles que l’obésité, le diabète de type 2, le cancer et les maladies neurodégénératives[23].
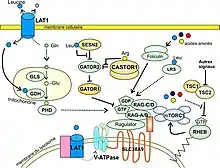
La régulation du complexe mTORC1 par les acides aminés est connue depuis la fin des années 1990 par les travaux de Hara et al.[25] démontrant qu’une carence en acides aminés, notamment en leucine, supprime la phosphorylation des deux voies aval, S6K1 et 4E-BP1 (voir fig. 1), même en présence de facteurs de croissance. Le mécanisme de cette régulation a commencé à être compris en 2008 quand les protéines RAG[n 4] et leurs rôles ont été identifiés. Le complexe des RAG, associé au RAGULATOR, est responsable de l’ancrage du complexe mTORC1 à la membrane du lysosome[24].
La connaissance du mécanisme d’activation de mTOR par la leucine a maintenant beaucoup progressé. Trop complexe pour être exposé ici, nous renvoyons les lecteurs à Yann Cormerais[24] ou Jacques Robert[22]. La leucine intervient au niveau d’un super-complexe composé de GATOR1 et GATOR2. En 2015 et 2016, deux études[26] - [27] ont montré que la protéine Sestrin2 se fixe sur GATOR2 et l’inhibe de façon dépendante de la leucine. En présence de leucine, GATOR2 se fixe sur GATOR1 et l’inhibe ce qui pour effet d’entrainer le recrutement de mTORC1 et son activation par Rheb. Ces résultats expliquent, presque 20 ans plus tard, les premières observations de Hara et al. sur la régulation de mTORC1 par la leucine.
En plus de l’explication par l’axe Castor1/Sestrin2-GATOR2/1-Rags, une étude de 2011 a suggéré que la détection des acides aminés se faisait à l’intérieur du lysosome[28] où les acides aminés doivent s’y accumuler pour pouvoir activer mTORC1. Le transporteur transmembranaire SLC38A9 de l’arginine interagit avec le super complexe Rags-RAGULATOR et son invalidation supprime l’activation de mTORC1 par l’arginine.
mTORC1 peut aussi être activé indépendamment par l’insuline et IGF-1, via le complexe TSC.
Régulation de la synthèse protéique par mTORC1
mTORC1 joue un rôle majeur dans la coordination entre d’une part la synthèse des ribosomes et la traduction des ARNm et d’autre part le statut énergétique et la disponibilité en nutriments.
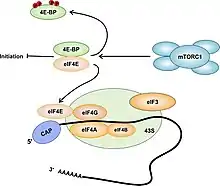
Une voie de contrôle de la synthèse protéique passe par la phosphorylation de la protéine eIF4E (en), effectuée par mTORC1 (fig. 1). La protéine 4E-BP1 est liée étroitement au facteur d'initiation eIF4E qui est responsable du recrutement du complexe d’initiation au niveau de la coiffe des ARNm (fig. 3). Une fois le complexe mTORC1 activé, il phosphoryle 4E-BP1 qui se désolidarise alors de eIF4E, permettant le recrutement d’une autre protéine, eIF4G et par incidence de eIF4A à l’extrémité 5’ d’un ARNm. Le complexe ainsi formé, constitue avec le ribosome le complexe 48S qui permet le démarrage (initiation en anglais) de la traduction des ARN messagers Cap-dépendants[29] - [n 5].
L’autre voie de contrôle de la synthèse protéique passe par la phosphorylation des protéines S6K1 et S6K2[30] (toujours par mTORC1). Lorsqu’elle n’est pas phosphorylée, la S6K se fixe et inhibe le complexe avec eIF3, bloquant ainsi l’initiation de la traduction. Mais une fois activée par mTORC1, la S6K se sépare de eIF3 lui permettant de se fixer au complexe d’initiation[24].
Notes
- exercices de musculation
- une synthèse protéique supérieure à une protéolyse, conduit à un gain protéique ou « accrétion protéique »
- mTOR= mechanistic target of rapamycin « cible mécanique de la rapamycine », un médicament immunosuppresseur
- Ras related GTPases
- les protéines ribosomiques (constituant des ribosomes) sont issues de la traduction d’ARNm particuliers qui possèdent tous un élément 5’ Terminal OligoPyrimidine (5’TOP) à l’extrémité 5’ de leur 5’UTR
Références
- Masse molaire calculée d’après « Atomic weights of the elements 2007 », sur www.chem.qmul.ac.uk.
- (en) Francis A. Carey, « Table of pKa and pI values », sur Département de chimie de l'université de Calgary, (consulté le )
- (en) M. Beals, L. Gross, S. Harrell, « Amino Acid Frequency », sur The Institute for Environmental Modeling (TIEM) à l'université du Tennessee (consulté le )
- « Leucine (l-) » dans la base de données de produits chimiques Reptox de la CSST (organisme québécois responsable de la sécurité et de la santé au travail), consulté le 25 avril 2009
- Abrégé du dictionnaire Grec-Français A. Bailly, p. 531.
- Étymol. et Hist. 1832 (Raymond). Composé de leuc(o)-* et de -ine*
- Elitsa A Ananieva, Jonathan D Powell, Susan M Hutson, « Leucine Metabolism in T Cell Activation: mTOR Signaling and Beyond », Advances in Nutrition, vol. 7, no 4, , p. 798-805 (lire en ligne)
- Denise Ferrier, Biochemistry (Sixth Edition), Wolters Kluwer, Lippincott William & Wilkins,
- Food and Nutrition Board, Institute of Medicine of the National Academies, Dietary Reference Intakes, The National Academies Press, (lire en ligne)
- Stephan van Vliet, Nicholas A Burd, and Luc JC van Loon, « The Skeletal Muscle Anabolic Response to Plant - versus Animal - Based Protein Consumption », Journal of Nutrition, vol. 145, , p. 1981-91
- « Leucine » (consulté le )
- Kazuyo Tujioka et al., « Effects of the Quality of Dietary Amino Acids Composition on Urea Synthesis in Rats », J. Nutr. Sci. Vitaminol, vol. 57, , p. 48-55
- Garlick PJ, Grant I., « Amino acid infusion increases the sensitivity of muscle protein synthesis in vivo to insulin. Effect of branched-chain aminoacids », Biochim. J, no 254, , p. 579-584
- Buse MG, Reid SS., « Leucine. A possible regulator of protein turnover in muscle. », J. Clin. Invest., vol. 56, , p. 1250-1261
- Tang JE, Moore DR, Kujbida GW, Tarnopolsky MA, Phillips SM., « Ingestion of whey hydrolysate, casein, or soy protein isolate: effects on mixed muscle protein synthesis at rest and following resistance exercise in young men », J Appl Physiol, vol. 107, , p. 987-92
- Verhoeven S, Vanschoonbeek K, Verdijk LB, Koopman R, Wodzig WK, Dendale P, van Loon LJ, « Long-term leucine supplementation does not increase muscle mass or strength in healthy elderly men. », Am. J. Clin. Nutr., vol. 89, no 5, , p. 1468-75
- Volek JS et al, « Whey protein supplementation during resistance training augments lean body mass », J Am Coll Nutr., vol. 32, no 2, , p. 122-35
- Xavier Bigard CERIN, « Quelles stratégies pour minimiser la sarcopénie du sujet âgé, septembre 2013 »
- João A.B. Pedroso, Thais T. Zampieri, Jose Donato Jr, « Reviewing the Effects of l-Leucine Supplementation in the Regulation of Food Intake, Energy Balance, and Glucose Homeostasis », Nutrients, vol. 7, no 5, , p. 3914-3937
- Stéphane Schneider, Nutrition, Collège des Enseignants de Nutrition, Elsevier Masson, , 534 p.
- K.A. Liu et al., « Leucine supplementation differentially enhances pancreatic cancer growth in lean and overweight mice. », Cancer Metab, vol. 2, no 1,
- Jacques Robert, « mTOR une protéine multifonctionnelle impliquée (entre autres) dans l’angiogenèse », VEGF Actu, vol. 33, (lire en ligne)
- Nicolas Saucisse, Dissection du rôle de la voie intracellulaire de mTORC1 dans les circuits hypothalamiques à la mélanocortine régulant la prise alimentaire, Thèse, Neuroscience, Université de Bordeaux, (lire en ligne)
- Yann Cormerais, Acides aminés et cancer: LAT1, un transporteur essentiel à l’activité mTORC1 et la croissance tumorale, Thèse, Université Nice Sophia Antipolis, (lire en ligne)
- Hara, Yonezawa et al., « Amino acid sufficiency and mTOR regulate p70 S6 kinase and eIF-4E BP1 through a common effector mechanism », J Biol Chem, vol. 273, no 23, , p. 14484-94
- Kim, J.S., S.H. Ro et al., « , Sestrin2 inhibits mTORC1 through modulation of GATOR complexes », Sci Rep, vol. 5, no 9502,
- Wolfson, Chandranupong et al., « Sestrin2 is a leucine sensor for mTORC1 pathway », Science, vol. 351, no 6268, , p. 43-48
- Zoncu, Bar-Peled et al., « mTORC1 senses lysosomal amino acids through an inside-out mechanism that requires vacuolar H+ ATPase », Science, vol. 334, no 6056, , p. 678-683
- Blagden, S.P., Willis, A.E., « The biological and therapeutic relevance of mRNA translation in cancer », Nat Rev Clin Oncol, vol. 8, , p. 280-91
- Thoreen C.C., Chantranupong H.R et al., « A unifying model for mTORC1-mediated regulation of mRNA translation », Nature, vol. 485, no 7593, , p. 109-113