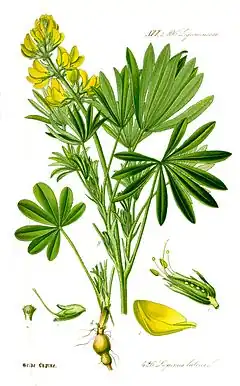
Fabaceae
Les Fabaceae, ou Leguminosae[2] (Légumineuses), sont une famille de plantes dicotylédones de l'ordre des Fabales. C'est l'une des plus importantes familles de plantes à fleurs, la troisième après les Orchidaceae et les Asteraceae par le nombre d'espèces. Elle compte environ 765 genres regroupant plus de 19 500 espèces[3]. Sur le plan économique, les Fabaceae sont la deuxième famille en importance après les Poaceae et constituent une source de protéines végétales indispensable pour l'alimentation humaine et animale.
| Règne | Plantae |
|---|---|
| Sous-règne | Tracheobionta |
| Division | Magnoliophyta |
| Classe | Magnoliopsida |
| Sous-classe | Rosidae |
| Ordre | Fabales |
- Leguminosae
- Papilionaceae

Étymologie


Le nom de cette famille, Fabaceae, est formé d'après le nom de genre Faba (fève en latin), de l’indo-européen commun *bhabha, « fruit sec »[4]. Le nom de genre Faba n'est plus utilisé, ayant laissé place au genre Vicia. Un représentant de l'ancien genre Faba (du latin faba, fève) est la fève, anciennement Faba vulgaris, renommée en Vicia faba.
Le nom des légumineuses vient du bas latin legumen. L'étymologie populaire rapproche ce terme du verbe latin legere « cueillir », « choisir » et par extension « lire ». Cependant, l'étymologie exacte reste problématique car on est plutôt en présence d'un mot non indoeuropéen, emprunté à une langue inconnue[5].
Description
Les Fabacées, au sens large, sont des plantes herbacées, des arbustes, des arbres ou des lianes. C'est une famille à répartition cosmopolite, présente dans tous les continents (à l'exception de l'Antarctique), des zones froides aux zones tropicales. La fonction chlorophyllienne est parfois transférée aux tiges.
De nombreuses espèces, principalement chez les Faboideae et Mimosoideae ont la particularité de puiser l'azote à la fois dans le sol et l'air. Elles se caractérisent par une activité symbiotique de fixation de l'azote atmosphérique grâce aux bactéries du genre Rhizobium, qui produisent des nodosités sur les racines et permettent des échanges d'azote avec la terre. L'agriculture exploite cette particularité naturelle en alternant la culture de Faboideae avec celle d'autres végétaux cultivés bénéficiant de cet apport.
Plusieurs légumineuses sont d'importantes plantes cultivées parmi lesquelles le soja, les haricots, les pois, le pois chiche, l'arachide, la lentille cultivée, la luzerne cultivée, différents trèfles, les fèves, le caroubier, la réglisse, etc. Comme les céréales, certains fruits et des légumes-racines tropicaux, un certain nombre de légumineuses sont des aliments de base pour l'homme depuis des millénaires et sont intimement liés à l'évolution humaine[6]. Les graines appelées légumes secs de certaines espèces telles que les gesses, les fèves, les lentilles et les pois font partie des premières espèces cultivées par l'homme dans le Croissant fertile, dès le Néolithique, pour son alimentation[7]. Grâce à leur aptitude à former des relations symbiotiques avec des bactéries fixatrices d'azote, de nombreuses légumineuses sont riches en protéines (teneur variant entre 17 et 27 %, deux à trois fois plus que les graines des céréales majeures) et ont un profil nutritionnel en acides aminés comparable à un steak de bœuf et plus intéressant que celle des poacées (elles sont déficitaires en acides aminés soufrés — méthionine, cystéine — alors que les céréales ont des teneurs d'acides aminés inférieurs, à l'exception de la méthionine, et sont déficitaires en lysine)[8]. Ces caractéristiques rendent la consommation de leurs graines complémentaires des régimes à base de céréales et expliquent qu'elles font partie des cultures fondatrices du Néolithique qui voit la lente domestication des plantes dans plusieurs foyers[9]. Les travaux de l'historien Fernand Braudel[10] sur les rapports entre la culture vivrière dominante, la densité du peuplement et le régime sociopolitique, suggèrent que la quête des aliments et leurs différents apprêts sont à l'origine de la civilisation du blé associée aux lentilles au Moyen-Orient et en Europe, la civilisation du riz associée au soja en Asie, la civilisation du maïs associée aux haricots en Amérique, et la civilisation de la houe dans d'autres régions du monde[11] - [12].
Nomenclature et systématique
Synonymes
Liste des synonymes selon ITIS[13] :
- Acaciaceae E. Mey.
- Aspalathaceae Martinov
- Astragalaceae Bercht. & J. Presl
- Bauhiniaceae Martynov
- Caesalpiniaceae
- Cassiaceae Vest
- Ceratoniaceae Link
- Ciceraceae W. Steele
- Coronillaceae Martynov
- Dalbergiaceae Martinov
- Daleaceae Berchtold & J. Presl
- Detariaceae Hess
- Galedupaceae Martynov
- Geoffroeaceae Martius
- Hedysaraceae Bercht. & J. Presl
- Inocarpaceae Zoll.
- Leguminosae Juss., nom. cons.
- Lotaceae Oken
- Mimosaceae R. Br., nom. cons.
- Papilionaceae Giseke, nom. cons.
- Phaseolaceae Mart
- Swartziaceae Bartl.
- Tamarindaceae Martinov
- Trifoliaceae Berchtold & J. Presl
- Viciaceae Oken
En français, cette famille est appelée couramment Légumineuses (Leguminosae, Jussieu, 1789) ou Papilionacées (Papilionaceae, Tournefort, 1694), mais ce ne sont pas de vrais synonymes. Chaque nom s'applique à un caractère particulier du fruit ou de la fleur de cette famille de plantes.
Schématiquement, le terme Légumineuses désigne des plantes dont le fruit est une gousse (legumen désignant en latin les « légumes », c'est-à-dire les végétaux dont les fruits sont des gousses[14]) et Papilionacées des légumineuses à fleurs irrégulières comportant cinq pétales inégaux ayant quelque ressemblance avec un papillon en position de vol[15].
Classification
Selon les classifications, la composition de cette famille varie :
- Le nom Fabaceae, au sens limité, est adopté en classification classique de Cronquist (1981). Ce groupe est nommé Fabaceae (stricto sensu) ou Papilionaceae. Cette famille comprend 12 000 espèces réparties en plus de 400 genres. (En classification phylogénétique, ce groupe des plantes est la sous-famille Faboideae.)
- Le nom Fabaceae, au sens large, est adopté en classification phylogénétique APG II (2003). Ce groupe est nommé Fabaceae (lato sensu) ou Leguminosae. Cette famille comprend 18 000 espèces réparties dans trois sous-familles (en classification classique, ce groupe des plantes est l'ordre des Fabales avec trois familles).
Les trois sous-familles reconnues classiquement sont les suivantes :- Mimosoideae, aux fleurs régulières (actinomorphes) ;
- Caesalpinioideae, aux fleurs pseudo-papilionacées (généralement zygomorphes) ;
- Faboideae ou Papilionoideae, aux fleurs typiques en « papillon » (zygomorphes).
Caractéristiques botaniques
Appareil végétatif

Au niveau de l'appareil végétatif, les Fabaceae sont représentées par des plantes à port très variable, herbacées annuelles ou vivaces, dressées, lianes ou plantes volubiles, arbustes et même arbres. Les plantes ligneuses (arbres, arbrisseaux), ordinairement regroupées dans les Caesalpinioideae et les Mimosoideae, se trouvent préférentiellement dans les régions chaudes (tropicales ou méditerranéennes). Les herbacées, représentées généralement par les Faboideae, prédominent plutôt dans les régions tempérée. Les lianes volubiles (Phaseolus, Physostigma) peuvent s'enrouler par des vrilles[17].
Les feuilles alternes, stipulées (les stipules sont parfois peu visibles : Anthyllis, Lotus), sont primitivement composées imparipennées (cependant paripennées chez les Mimosideae et Cesalpiniodeae)[18]. L'évolution foliaire conduit à des feuilles réduites : disparition de la foliole terminale (feuilles paripennées des fèves) ; transformation des folioles terminales en vrille (vesces, pois) ; réduction à 3 folioles comme le trèfle mais en compensation, par un phénomène dit de « surévolution », développement de stipules (Lotier), de stipelles (Faboideae : Phaseolus, Vigna), ou subdivision des folioles latérales (feuille palmée des lupins) ; réduction à une seule foliole chez les feuilles supérieure des Genêts à balais ; réduction à deux folioles latérales (Lathyrus sativus) qui peuvent même totalement disparaître, tandis que par compensation, les stipules acquièrent la taille de folioles (Lathyrus aphaca) ; transformation des stipules en épines (Robinia, Gleditsia)[19]. Cette réduction peut être telle que les fonctions photosynthétiques sont transférées aux tiges ou aux pétioles transformés en phyllodes. Les Fabacées ont typiquement un pulvinus à la base des pétioles impliqués dans le mouvement de repli des feuilles ou folioles répondant à l'alternance du jour et de la nuit (nyctinastie) ou à une stimulation tactile (thigmonastie)[20].
On observe normalement la présence de nodules racinaires (plus rarement caulinaires) fixateurs de l’azote atmosphérique sur les racines chez les Faboideae et les Mimosoideae, alors qu'ils sont absents chez la plupart des Caesalpinioideae. Ces nodosités présentes chez 88 % des Légumineuses[21] sont le résultat d'une symbiose entre des bactéries fixatrices d'azote, essentiellement les rhizobiums, et ces différentes espèces de légumineuses. Cette caractéristique biologique correspond à une particularité métabolique des Fabacées, la présence d'une hémoprotéine fixatrice de dioxygène, la léghémoglobine (ou LegHb), très proche de l’hémoglobine. Cette protéine permet de fixer l'oxygène pour former un milieu anaérobie favorable à la fixation du N2 par le rhizobium. Le système racinaire est généralement pivotant, permettant une association fréquente des Légumineuses aux Graminées dotées d'un système racinaire fasciculé. Ce système permet une meilleure exploration du sol grâce à la racine pivotante qui pompe l'eau à de grandes profondeurs et aux nombreuses racines secondaires (ces racines et radicelles se déploient à quelques centimètres de la surface du sol, mais à plusieurs mètres de la plante) et contrebalancent le phénomène de feutrage (les racines superficielles des Graminées s'entremêlent et finissent par former à la surface du sol un tapis serré appelé « feutre »)[22].
Du point de vue phytochimique, de nombreuses parties de l'appareil végétatif synthétisent des métabolites secondaires qui ont un rôle de protection contre l'herbivorie mais aussi d'attraction vis-à-vis des animaux pollinisateurs ou disperseurs de graines. Les légumineuses constituent une source majeure de matières riches en protéines pour l'alimentation animale (source primaire d'azote assimilable pour les animaux herbivores, dont les êtres humains, qui les consomment). Aussi leurs organes de stockage et leurs graines produisent une très grande diversité de métabolites toxiques qui participent à l'arsenal de défenses chimiques contre les herbivores. Ces métabolites secondaires peuvent être regroupés en trois classes principales dérivant de voies de synthèses différentes : les composés à base azotée (alcaloïdes, amines, peptides, glucosinolates cyanogénétiques), les composés phénoliques (anthocyanines, coumarines, tannins), les terpénoïdes[23].
Appareil reproducteur


Les fleurs sont groupées en inflorescences de formes variées : grappe souvent allongée, épi, glomérule sphérique ou cylindrique (parfois pendant: Mimosoideae)[19]. L'inflorescence peut être réduite à une seule fleur ou regrouper et condenser les fleurs pour former un pseudanthe, structure mimant une fleur unique (Mimosoideae du genre Dichrostachys, Neptunia et Parkia)[25]. Les fleurs hermaphrodites, encore régulières chez les Mimosoideae, deviennent très spécifiques chez les autres sous-familles et marquées par une forte zygomorphie[26]. Le périanthe hétérochlamyde comporte un calice gamosépale réduit à cinq dents, parfois bilabié (deux lèvres chez les Genêts). La corolle dialypétale est dite « papilionacée » : le pétale supérieur, en position adaxiale (dorsale), appelé étendard en raison de sa position dressée, est soit très développé et recouvrant les pétales latéraux (Faboideae en raison de leur préfloraison descendante), soit discret et recouvert par les pétales latéraux (Caesalpinioideae en raison de leur préfloraison ascendante). Dans certains cas, il peut se retrouver en position abaxiale par résupination de la fleur. Les deux pétales latéraux, appelés ailes, sont libres et identiques entre eux ou parfois légèrement adhérents aux pétales inférieurs qu'ils recouvrent (Faboideae) ou dont ils sont recouverts (Caesalpinioideae). Les deux pétales ventraux sont semblables et souvent connés entre eux (simulant un pétale unique) pour former la carène (terme en référence à la coque d'un bateau) entourant les étamines et le pistil, permettant la présentation secondaire du pollen (dispositif similaire chez les Polygalacées, résultant du phénomène de convergence évolutive)[27]. Ces deux pétales comprimés sont à l'origine d'une fécondation par cléistogamie[28]. Les racèmes très contractés des Trèfles en forme de glomérule sont associés à une corolle gamopétale[29]. La pollinisation est entomophile, très rarement anémophile, les fleurs pouvant également se féconder par autopollinisation[30].
- L'androcée généralement diplostémone est formé de 10 étamines soudées en tube (Faboideae à l'androcée monadelphe avec un tube fendu du côté de l'étendard, ou diadelphe avec 9 étamines soudées et une libre ou partiellement libre au-dessus) ou parfois libres (Mimosoideae, Caesalpinioideae). Les deux cycles d'étamines sont à anthères basifixes, introrses, à déhiscence longitudinale (parfois poricide apicale). Le gynécée monocarpellé forme un ovaire supère à placentation marginale (contenant deux à plusieurs ovules anatropes ou campylotropes, bitégumentés, avec peu ou pas d’albumen), surmonté par un style allongé de forme variable. Un anneau nectarifère est souvent présent autour de l'ovaire (disque intrastaminal) ou sur la surface interne de l'hypanthium, assurant une pollinisation entomophile[26]. Le fruit est typiquement une gousse multiséminée mais il existe de nombreuses modifications morphologiques. La dissémination est également variée[31] : autochorie de la glycine, zoochorie (notamment ornithochorie par les oiseaux, chiroptérochorie par les chauve-souris)[32].
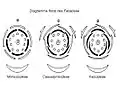 Diagramme floral des Fabaceae.
Diagramme floral des Fabaceae. Fleur de gesse aphylle vue de profil : calice (avec, sur le dessus, une bosse qui correspond au nectaire), étamines et style.
Fleur de gesse aphylle vue de profil : calice (avec, sur le dessus, une bosse qui correspond au nectaire), étamines et style. Gousses de différentes espèces de fabacées tropicales.
Gousses de différentes espèces de fabacées tropicales.
Phytochimie
Les Fabaceae produisent de nombreux métabolites secondaires qui jouent un rôle soit pour lutter contre les animaux herbivores ou phytophages et les micro-organismes, soit pour attirer les animaux pollinisateurs et disperseurs de fruits et graines. Certains métabolites secondaires (flavonoïdes, triterpènes, pinitol) ont une très large distribution et sont présents dans la quasi-totalité des tribus, tandis que d'autres ne sont présents que dans un faible nombre de taxons. Beaucoup d'espèces ayant la capacité de fixer l'azote atmosphérique, la famille des légumineuses produit davantage de métabolites secondaires contenant de l'azote que d'autres familles de plantes[23].
Les composés azotés comprennent[23] :
- des alcaloïdes et amines (quinolizidine, pyrrolizidine, indolizidine, pipéridine, pyridine, pyrrolidine, indole simple, érythrine, isoquinoléine simple et alcaloïdes d'imidazole, polyamines, phényléthylamine, tyramine et dérivés de tryptamine),
- des acides aminés non protéinogènes (canavanine, albiziine, carboxyéthylcystéine, willardiine, homoarginine, mimosine) ;
- des glucosides cyanogènes (prunasine, linamarine, lotaustraline, proacacipétaline) ;
- des peptides (lectines, inhibiteurs de protéases (de la trypsine) et d'amylases, peptides antimicrobiens, cyclotides).
Les métabolites secondaires sans azote comprennent[23] :
- des composés phénoliques (phénylpropanoïdes, flavonoïdes, isoflavones, catéchines, anthocyanines, tanins, lignanes, coumarines et furanocoumarines),
- des polycétides (anthraquinones)
- des terpénoïdes (notamment des triterpénoïdes, des saponines stéroïdiennes et des tétrapeptides).
Importance économique
Cette famille de plantes se présente au deuxième rang de production mondial, derrière les céréales, pour l’alimentation humaine. En 2014 selon la FAO, plus de 300 millions de tonnes de légumineuses (soja, arachide, haricots, pois, fèves et lentilles) sont produites dans le monde (dont les deux-tiers pour le soja) sur une surface de 190 million ha (soit 13 % des surfaces cultivées, contre 70 % pour les céréales)[33]. 670 des 750 genres et 18 000 des 19 000 espèces de la famille des légumineuses ont un rôle important en tant que grains, pâturages et en agroforesterie[34]. La complémentarité nutritionnelle entre les familles des céréales et des légumineuses pourrait expliquer que dans de nombreux foyers de domestication, elles aient été domestiquées ensemble[35].
Elle est tout d'abord une source essentielle de protéines végétales pour l'alimentation humaine ou animale. Elle fournit également des matières grasses (huile de soja et d'arachide), du bois (nombreuses espèces exotiques et précieuses comme le palissandre), et sert à bien d'autres usages.
Comme ces plantes ne demandent pas d'engrais azotés, du fait de leur capacité à fixer l'azote atmosphérique grâce aux bactéries symbiotiques du genre rhizobiums présentes dans leurs racines, elles tiennent une place particulière dans les rotations culturales.
La famille des Fabacées comprend de nombreuses plantes cultivées d'intérêt économique : l'arachide, le fenugrec, la fève, la féverolle, les gesses, les haricots, le haricot mungo (pousses de soja germé), les lentilles, les lotiers, les lupins, les luzernes, le palissandre, les pois, le pois chiche, la réglisse, le rooibos, le sainfoin, le soja, les trèfles, les vesces…
Une multitude de plantes herbacées ou ligneuses de cette famille présentent un intérêt ornemental, comme la glycine, les lupins, les mimosas, les pois de senteur, etc.
Différentes plantes de cette famille fournissent aussi des substances diverses pour tanner des cuirs, fabriquer des colles, soigner les fièvres…
Intérêt dans l'alimentation
Les légumineuses jouent un rôle très important dans l'alimentation humaine et animale grâce à leur haute teneur en protéines (comprise entre 20 et 42 %[36] - [37] - [38] - [39]) et en acides aminés essentiels (souffrant d'un déficit général en acides aminés soufrés, elles ont une teneur très intéressante en lysine alors que les protéines de céréales sont pauvres en lysine)[40]. Les produits dérivés peuvent en contenir plus encore, comme le tourteau de soja (50 % environ). Les autres nutriments importants sont l'amidon, et éventuellement les lipides pour les légumineuses qui sont également des oléagineux (soja qui contient environ 20 % d'huile, arachide qui en contient presque 50 %)[41]. Après cuisson, la teneur en protéines est comprise entre 5 %[42] et 18 %[43], ce qui est un peu inférieur à la viande cuite.
Les légumineuses sont riches en minéraux (fer, calcium, zinc, cuivre, manganèse)[44] et en fibres, mais relativement pauvres en vitamines, sauf celles du groupe B - celles-ci étant de plus partiellement détruites à la cuisson. Les légumineuses contiennent également des facteurs anti-nutritionnels[45], comme les tanins, la lectine, les facteurs anti-trypsiques, l'acide phytique, les alpha-galactosides (des FODMAP), la vicine-convicine, et nécessitent souvent une préparation technique avant consommation (trempage, cuisson, fermentation, toastage du tourteau de soja, par exemple).
Riches en protéines végétales dites « incomplètes », les céréales et les légumineuses s'associent pour apporter des protéines complètes comparables à celles de la viande, du poisson ou des œufs, l'apport recommandé étant de 2⁄3 de légumineuses et 1⁄3 de céréales[46].
Intérêt dans l'alimentation animale
Le soja est l'une des principales sources de protéines en alimentation animale. Le pois protéagineux et la féverolle sont cultivés dans ce but. Différentes espèces comme la luzerne, les trèfles, le sainfoin, le lotier… jouent un rôle essentiel dans la valeur des prairies.
Intérêt dans l'alimentation humaine
Les légumineuses, riches en glucides complexes, micronutriments, protéines, vitamines du groupe B et fibres sont une partie essentielle d'un régime alimentaire sain[47].
Les légumineuses fournissent des protéines très bon marché, et en particulier de la lysine. En les combinant à d'autres protéines végétales comme celles des céréales, on obtient une source équilibrée de protéines.
La soixante-huitième session (en) de l'Assemblée générale des Nations Unies a déclaré 2016 année internationale des légumineuses « pour sensibiliser et pour célébrer le rôle des haricots, pois chiches, lentilles et autres légumineuses dans l'alimentation mondiale ».
Commerce
La production mondiale de légumineuses cultivées pour leurs graines dépasse les 300 millions de tonnes annuelles, dont les deux tiers pour le seul soja[48]. Cette plante ainsi que l'arachide (ou cacahuète) est souvent rattachée aux oléagineux, ou encore aux protéo-oléagineux.
L'Inde est à la fois le premier producteur et importateur mondial de légumineuses à graines au sens strict (pois, haricots, fèves, lentilles). Le Canada, la Birmanie, l'Australie et les États-Unis sont des exportateurs importants (Nota : ce sont, dans cet ordre, les quatre premiers fournisseurs de l'Inde). La très grande majorité des cultures de légumineuses des États-Unis sont des cultures de cacahuètes et de soja destinées à l'alimentation du bétail et à l'extraction d'huile végétale, considérées comme des oléagineux en raison de leur mise en valeur sous forme d'huile avant tout.
Intérêt agronomique et environnemental

Grâce à leur capacité à fixer l'azote atmosphérique, les légumineuses sont, d'une manière générale, des plantes « pionnières » capables de coloniser des milieux pauvres en azote, et jouent de ce fait un rôle important dans la végétalisation des sols. Cela leur permet de se développer les premières dans des sols pauvres[49]. Elles facilitent par la suite le développement d'autres espèces. Cette caractéristique de fixation de l'azote leur permet d'accumuler des concentrations importantes d'azote, d'où leur utilisation comme engrais verts : la restitution au sol d'une partie de leur biomasse favorise le rétablissement de la fertilité des sols après la culture de plantes plus exigeantes, telles que les céréales. En outre, leur présence dans la rotation des cultures contribue à la réduction du besoin d'engrais à base d'azote synthétique et des émissions de gaz à effet de serre[50]
Beaucoup d'espèces de cette famille sont utiles pour les pollinisateurs. Qu'elles soient prairiales ou arbustives, nombre d'entre elles sont très favorables aux abeilles[51].
Quand elles sont introduites dans l'alimentation animale, elles contribuent à améliorer le taux de conversion des aliments en réduisant simultanément les émissions de gaz à effet de serre[52].
À l'inverse, certains membres de cette famille sont des adventices particulièrement difficiles à contrôler et peuvent devenir envahissantes.
Phylogénie

La phylogénie des légumineuses a fait l'objet de nombreuses études par des groupes de recherche du monde entier. Ces études ont utilisé des données de morphologie, l'ADN (l'intron des chloroplastes trnL, les gènes chloroplastiques rbcL et matK, ou les espaceurs ribosomaux ITS) et l'analyse cladistique, afin d'étudier les relations entre les différentes lignées de la famille. Toutes les études ont démontré systématiquement le caractère monophylétique des Fabaceae[53].
Les études ont en outre confirmé que les sous-familles traditionnelles des Mimosoideae et des Papilionoideae étaient toutes deux monophylétiques mais qu'elles étaient toutes deux nichées dans la sous-famille des Caesalpinioideae qui est, elle, paraphylétique[54] - [53].
Toutes les différentes approches ont donné des résultats similaires en ce qui concerne les relations entre les principaux clades de la famille[55] - [56] - [57] - [58] - [59] - [60] - [61] - [62] - [63].
Après de longues discussions dans la communauté phylogénétique des légumineuses, le Legume Phylogeny Working Group (groupe de travail sur la phylogénie des légumineuses) a subdivisé les Fabaceae en six sous-familles au lieu de trois, ce qui a nécessité la recirconscription des Caesalpinioideae en une sous-famille réduite (Caesapinioideae, sensu stricto), incluant l'ancienne sous-famille des Mimosoideae, et quatre nouvelles sous-familles[64]
| Fabales |
| |||||||||||||||
Liste des genres
Principaux genres
Les genres les plus importants sont les suivants : Astragalus (plus de 2 400 espèces), Acacia (plus de 950 espèces), Indigofera (environ 700 espèces), Crotalaria (environ 700 espèces) et Mimosa (environ 500 espèces)[65].
Dans cette famille, on peut citer les genres suivants :
(Le genre-type Faba est un synonyme de Vicia, et c'est sous ce nom qu'il est repris dans la liste des genres ci-dessous.)
Sous-famille Mimosoideae
(liste non exhaustive)
- Albizia avec l'arbre de soie
- Anadenanthera
- Entada avec le cœur de la mer
- Inga ;
- Pithecellobium ;
- Acacia, avec le mimosa ;
- Mimosa.
Sous-famille Caesalpinioideae
(liste non exhaustive)
- Afzelia
- Amherstia
- Bauhinia ;
- Brachystegia ;
- Caesalpinia ;
- Cassia ;
- Ceratonia, avec le Caroubier (Ceratonia siliqua) ;
- Cercis, avec l'arbre de Judée (Cercis siliquastrum) ;
- Colophospermum ;
- Copaifera ;
- Delonix ;
- Gleditsia avec le févier épineux ;
- Haematoxylum ;
- Melanoxylon ;
- Parkinsonia ;
- Paubrasilia, le pernambouc ou bois-brésil ;
- Peltophorum ;
- Schizolobium parahyba ;
- Sclerolobium ;
- Senna ;
- Tamarindus avec le tamarinier ;
- etc.
Sous-famille Faboideae ou Papilionoideae
(liste non exhaustive)
- Abrus precatorius ; la liane réglisse ;
- Aeschynomene ;
- Amorpha ;
- Anthyllis, avec la vulnéraire ;
- Arachis, l'arachide ;
- Aspalathus linearis, le rooibos ;
- Astracantha ;
- Astragalus, les astragales ;
- Bowdichia;
- Brongniartia;
- Canavalia, avec le pois sabre et le pois-bord-de-mer ;
- Cicer, avec le pois chiche ;
- Cyclopia, le Honeytea ;
- Cytisus, certains genêts comme le genêt à balais ;
- Coronilla, certaines coronilles ;
- Dalbergia, avec les palissandres ;
- Derris
- Desmodium adscendens ;
- Dorycnium, les dorycnies et bonjeanies ;
- Ebenus L., 1753[note 3]
- Eutaxia, genre australien;
- Genista, c'est le genêt poilu ou le genêt des teinturiers ;
- Glycine, qui donne le soja (ne pas confondre avec les « glycines » du genre Wisteria) ;
- Glycyrrhiza glabra, la réglisse ;
- Hippocrepis avec le fer à cheval ;
- Lablab, avec le pois antaque ou dolique d'Égypte ;
- Laburnum, avec le cytise faux ébénier ;
- Lathyrus, avec les gesses et le pois de senteur;
- Lens, avec la lentille cultivée ;
- Lotus, les lotiers ;
- Lupinus, les lupins ;
- Medicago, les luzernes et la minette ;
- Melilotus, les mélilots ;
- Onobrychis les sainfoins ;
- Ononis les bugranes ;
- Phaseolus, les nombreux haricots dont le haricot d'Espagne ;
- Physostigma, avec la fève de Calabar ;
- Pisum, avec le petit pois ou pois potager, le pois protéagineux et le pois fourrager ;
- Robinia, avec le robinier faux-acacia au bois dur imputrescible ;
- Sophoreae ;
- Trifolium, les nombreux trèfles ;
- Trigonella, le fenugrec ;
- Ulex, ce sont les ajoncs ;
- Vicia, les vesces, la fève et la féverolle ;
- Vigna, avec les doliques, le haricot azuki, le haricot mungo qui donne les « pousses de soja », le niébé ;
- Wisteria, avec les « glycines ».
Plantes emblèmes
Plusieurs espèces de Fabaceae ont été choisies comme plantes emblèmes par différents pays ou territoires :
- Erythrina crista-galli, l'érythrine crête-de-coq ou ceibo, est la fleur nationale de l'Argentine (depuis 1942) et de l'Uruguay[66] - [67].
- Enterolobium cyclocarpum, le guanacaste, est l'arbre national du Costa Rica depuis 1959[68].
- Paubrasilia echinata, le pernambouc ou bois du Brésil, est l'arbre national du Brésil depuis 1978[69].
- Acacia pycnantha, le mimosa doré, est la fleur nationale de l'Australie[70].
- Swainsona formosa, le pois du désert, est l'emblème floral de l'Australie-Méridionale depuis 1961[71].
- Bauhinia blakeana, l'arbre aux orchidées, est la fleur nationale de Hong Kong depuis 1965[72]. Cette espèce est également l'arbre emblématique de la ville de Zhuhai dans la province de Guangdong (Chine)[73].
- Le lupin est la fleur officielle du Texas. Il s'agissait à l'origine, en 1901, de Lupinus subcarnosus, mais la décision a été amendée en 1971 pour ajouter Lupinus texensis et « toute autre variété de lupin non encore enregistrée »[74].
- Trifolium pratense, le trèfle violet, est la fleur officielle du Vermont depuis 1895[75].
- Parkinsonia florida, le palo verde, est l'arbre officiel de l'Arizona depuis 1954[76].
- Intsia bijuga, le faux gaïac, est l'arbre officiel de Guam depuis 1969[77].
- Delonix regia, le flamboyant, est l'arbre officiel de îles Mariannes du Nord depuis 1979[78]. Cette espèce est également la fleur officielle de Saint-Christophe-et-Niévès[79].
- Cercis canadensis, le gainier du Canada, est l'arbre officiel de l'Oklahoma depuis 1937[80].
- Genista pilosa, le genêt poilu, est la fleur officielle de la province du Halland (Suède)[81].
- Cassia fistula, le cassier commun, est la fleur officielle de l'État du Kerala (Inde)[82].
- Senna auriculata, l’avaram, est la fleur officielle de l'État du Telangana (Inde)[83].
- L'ajonc (genre Ulex) a été choisi en 2016 comme emblème floral de la Bretagne à la suite d'un vote organisé par l'institut culturel de Bretagne[84].
- Le garapuvu (Schizolobium parahyba, Caesalpinioideae) a été désigné « arbre symbole » de la ville de Florianópolis (État de Santa Catarina, Brésil) par décision municipale du 25 mai 1992[85].
Le trèfle irlandais ou shamrock, symbole de l'Irlande, est en fait une feuille d'oxalis petite oseille, Oxalis acetosella, et non une feuille de trèfle Trifolium repens[86].
Notes et références
Notes
- « La structure de la brosse au sommet du style varie d'un genre à un autre ce qui indique que cette brosse à pollen est apparue plusieurs fois indépendamment (au moins huit fois semble-t-il) dans plusieurs lignées de Fabacées ». Cf Gérard Guillot, La planète fleurs, Quæ, , p. 115.
- « Le seul point d’appui dont bénéficie l’insecte, c’est le milieu de la carène qui lui sert de « centre de pression » ; ses pattes arrière poussent tandis que les pattes médianes et antérieures tirent, laissant au passage des traces de griffures sur les ailes et la carène. La carène descend sous la pression et comme la brosse à pollen reste immobile, elle se trouve poussée contre le corps velu de l’abeille qu’elle frotte sur le côté, tout en bloquant l’insecte dans cette position ». Cf Gérard Guillot, « Le xylocope et le pois vivace : une histoire d’amour musclée », sur zoom-nature.fr (consulté le ).
- Ne pas confondre avec Ebenus Kuntz, 1882 successivement appelé Maba puis Diospyros et qui a donné son nom à la famille des Ebenaceae
Références
- (en) Susana Magallon, Peter R. Crane and Patrick S. Herendeen, « Phylogenetic pattern, diversity, and diversification of Eudicots », Annals of the Missouri Botanical Garden, vol. 86, no 2, , p. 297-372 (DOI 10.2307/2666180)
- Le nom « Leguminosae » (nomen alternativum) est considéré comme validement publié, en raison d'un « long usage » par le Code international de nomenclature pour les algues, les champignons et les plantes, article 18.5
- (en) « New Subfamily Classification of the Leguminosae and Insights into Plastomes of the Mimosoid Clade », sur Chinese Academy of Sciences (consulté le ).
- (en) J. P. Mallory, Douglas Q. Adams, Encyclopedia of Indo-European Culture, Taylor & Francis, (lire en ligne), p. 55.
- Alain Rey, Dictionnaire historique de la langue française, Nathan, (lire en ligne), p. 2000.
- (es)Burkart, A. Leguminosas. In: Dimitri, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires. Pages : 467-538.
- Michel Pitrat et Claude Foury Histoire de légumes des origines à l'orée du XXIe siècle INRA 2003 (ISBN 2-7380-1066-0) p. 340.
- (en) Beryl Brintnall Simpson, Molly Conner-Ogorzaly, Economic Botany. Plants in Our World, McGraw-Hill, , p. 187-190.
- La consommation initiale de graines de légumineuses vient probablement du fait qu'elles étaient disponibles comme ressources, qu'elles avaient bon goût et qu'elles satisfaisaient la faim. La domestication des légumineuses se révélant complémentaires au niveau nutritionnel relève certainement d'un « phénomène accidentel particulièrement heureux ». Cf (en) Charles Bixler Heiser, Seed to Civilization. The Story of Food, Harvard University Press, , p. 117.
- Fernand Braudel, Civilisation matérielle, économie et capitalisme, XVe-XVIIIe siècle, Paris, Armand Colin, 1979, volume 1, chapitre « Le pain de chaque jour », p. 81-152.
- Outil agricole qui déterre les tubercules d'igname et de manioc en Afrique, de pomme de terre dans les Andes, de taro dans les îles du Pacifique.
- Jean Paul Charvet, L'alimentation dans le monde, Larousse, , p. 65.
- « Fabaceae Système mondial d'information sur la biodiversité (SMIB) », sur Système canadien d’information sur la biodiversité (SCIB) - Système d'information taxonomique intégré (SITI) (consulté le ).
- Informations lexicographiques et étymologiques de « légumineuse » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales.
- Informations lexicographiques et étymologiques de « papilionacée » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales
- Romaric Forêt, Dictionnaire des sciences de la vie, De Boeck Supérieur, , p. 4.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, (lire en ligne), p. 599.
- Marjorie Blamey, C. Grey-Wilson, La flore de France et d'Europe occidentale, Eclectis, , p. 199.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 602.
- William G. Hopkins, Physiologie végétale, De Boeck Supérieur, (lire en ligne), p. 408.
- (en) SM de Faria, SM Lewis, JI Sprent, JM Sutherland, « Occurrence of nodulation in the leguminosae », New Physiologist, vol. 111, no 4, , p. 607–619 (DOI 10.1111/j.1469-8137.1989.tb02354.x).
- Charles Victor Garola, Plantes fourragères, J.-B. Baillière et fils, , p. 178.
- (en) M. Wink, « Evolution of secondary metabolites in legumes (Fabaceae) », South African Journal of Botany, vol. 89, , p. 164–175 (lire en ligne).
- (en) Christian Westerkamp, « The co-operation between the asymmetric flower of Lathyrus latifolius (Fabaceae-Vicieae) and its visitors », Phyton, vol. 33, no 1, , p. 121-137 (lire en ligne).
- (en) Rodolphe Spichiger, Systematic botany of flowering plants : a new phylogenetic approach to angiosperms of the temperate and tropical regions, Science Publishers, , p. 202.
- Rodolphe-Edouard Spichiger, Vincent V. Savolainen, Murielle Figeat-Hug, Daniel Jeanmonod, Botanique systématique des plantes à fleurs : une approche phylogénétique nouvelle des angiospermes des régions tempérées et tropicales, Presses Polytechniques et Universitaires Romandes, , p. 182.
- (en) C. Westerkamp, A. Weber, « Keel flowers of the Polygalaceae and Fabaceae: a functional comparison », Botanical Journal of the Linnean Society, vol. 29, no 3, , p. 207-221 (DOI 10.1111/j.1095-8339.1999.tb00501.x).
- (en) Theresa M. Culley, Matthew R. Klooste, « The cleistogamous breeding system: a review of its frequency, evolution, and ecology in angiosperms », Botanical Review, vol. 73, no 1, , p. 1–30.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 604.
- (en) SC Tucker, « Floral development in legumes », Plant Physiology, vol. 131, no 3, , p. 911-926.
- Michel Botineau, Botanique systématique et appliquée des plantes à fleurs, Lavoisier, , p. 607.
- (en) L. Van der Pijl, Principles of dispersal in higher plants, Springer, , p. 162.
- (en) « Food and agriculture data », sur fao.org, .
- (en) RM Polhill, PH Raven, CH Stirton, « Evolution and systematics of the Leguminosae », in Advances in Legume Systematics Part 1 (eds RM Polhill, PH Raven), Royal Botanic Gardens, 1981, p. 1–26.
- (en) Paul Gepts, « Crop Domestication as a Long‐Term Selection Experiment », Plant breeding reviews, vol. 24, no 2, , p. 1-44 (DOI 10.1002/9780470650288.ch1).
- teneur en protéines de plusieurs aliments courants[PDF].
- Les protéines végétales[PDF].
- (en) USDA Database 16001, Beans, adzuki, mature seeds, raw.
- (en) USDA Database 16108, Soybeans, mature seeds, raw.
- (en) TL Wang, C Domoney, CL Hedley, R Casey, MA Grusak, « Can we improve the nutritional quality of legume seeds? », Plant Physiology, vol. 131, no 3, , p. 886–89 (DOI 10.1104/pp.102.017665).
- (en) Marcello Duranti, Cristina Gius, « Legume seeds: protein content and nutritional value », Field Crops Research, vol. 53, nos 1–3, , p. 31-45 (DOI 10.1016/S0378-4290(97)00021-X).
- (en) USDA Database 16006, Beans, baked, canned, plain or vegetarian.
- (en) USDA Database 16109, Soybeans, mature cooked, boiled, without salt.
- (en) MA Grusak, « Enhancing mineral content in plant food products », Journal of the American College of Nutrition, vol. 21 (3 Suppl), , p. 178-183.
- (en) YP Gupta, « Anti-nutritional and toxic factors in food legumes: a review », Plant Foods for Human Nutrition, vol. 37, no 3, , p. 201-228.
- (en) P. H. Graham & C. P. Vance, « Legumes: Importance and Constraints to Greater Use », Plant Physiol., vol. 131, no 3, , p. 872–877 (DOI 10.1104/pp.017004).
- Anne Schneider et Christian Huygue, Les légumineuses pour des systèmes agricoles et alimentaires durables, Quae, , 512 p. (ISBN 978-2-7592-2334-3, lire en ligne), p. 264
- source FAOSTAT.
- Eric Giraud, « Symbiose rhizobium/ légumineuse : un nouveau sésame », Médecine/Sciences, Inserm, no 23, , p. 663-664 (lire en ligne [PDF])].
- (en) Catherine Hénault & Cécile Revellin, « Inoculants of leguminous crops for mitigating soil emissions of the greenhouse gas nitrous oxide », Plant and Soil, vol. 346, , p. 289–296 (DOI 10.1007/s11104-011-0820-0).
- Liste des plantes attractives pour les abeilles [PDF], gnis.fr.
- « Les légumineuses et le changement climatique », sur fao.org, (consulté le ).
- (en) Martin F. Wojciechowski, Johanna Mahn et Bruce Jones, « Fabaceae », The Tree of Life Web Project, .
- (en) M. F. Wojciechowski, M. Lavin et M. J. Sanderson, « A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported sub clades within the family », American Journal of Botany, vol. 91, no 11, , p. 1846–62 (PMID 21652332, DOI 10.3732/ajb.91.11.1846)
- (en) P. F. Stevens, « Fabaceae », sur Angiosperm Phylogeny, (consulté le )
- (en) « Molecular evolution of the Leguminosae: phylogeny of the three subfamilies based on rbcL sequences », Biochemical Systematics and Ecology, vol. 24, no 5, , p. 365–378 (DOI 10.1016/0305-1978(96)00032-4, lire en ligne).
- (en) « Phylogenetic relationships in the Papilionoideae (Family Leguminosae) based on nucleotide sequences of cpDNA (rbcL) and ncDNA (ITS1 and 2) », Molecular Phylogenetics and Evolution, vol. 8, no 1, , p. 65–88 (DOI 10.1006/mpev.1997.0410, lire en ligne).
- (en) « A phylogeny of the chloroplast gene rbcL in the Leguminosae: taxonomic correlations and insights into the evolution of nodulation », American Journal of Botany, vol. 84, no 4, , p. 541–554 (DOI 10.2307/2446030, lire en ligne).
- (en) « Evolutionary significance of the loss of the chloroplast-DNA inverted repeat in the Leguminosae subfamily Papilionoideae », Evolution (journal), vol. 44, no 2, , p. 390–402 (DOI 10.2307/2409416).
- (en) « Diversification rates in a temperate legume clade: are there "so many species" of Astragalus (Fabaceae)? », American Journal of Botany, vol. 83, no 11, , p. 1488–1502 (DOI 10.2307/2446103).
- (en) Chappill JA., Advances in Legume Systematics, Part 7 : Phylogeny, Royal Botanic Gardens, Kew, UK, , 371 p. (ISBN 978-0-947643-79-9), « Cladistic analysis of the Leguminosae: the development of an explicit hypothesis ».
- (en) « Phylogenetic patterns and diversification in the caesalpinioid legumes », Botany (journal), vol. 86, no 7, , p. 697–718 (DOI 10.1139/B08-058).
- (en) « Reconstructing the deep-branching relationships of the papilionoid legumes », S. Afr. J. Bot., vol. 89, , p. 58–75 (DOI 10.1016/j.sajb.2013.05.001, lire en ligne).
- (en) The Legume Phylogeny Working Group (LPWG)., « A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny », Taxon (journal), vol. 66, no 1, , p. 44–77 (DOI 10.12705/661.3, lire en ligne).
- (en) CTI Reviews, Plant Systematics, Cram101 Textbook Reviews, , 80 p. (ISBN 978-1-4784-2888-6).
- (es) « El ceibo: flor nacional », sur Todo Uruguay, (consulté le ).
- (es) « INTA Chicos - Día de la Flor Nacional: el Ceibo », sur INTA Chicos (consulté le ).
- Gilbert Vargas Ulate. 1997. Geografía turística de Costa Rica. EUNED, 180 p. (ISBN 9977-64-900-6), 9789977649009.
- "Lei Nº 6.607, de 7 de dezembro de 1978. O Presidente da República, faço saber que o Congresso Nacional decreta e eu sanciono a seguinte Lei: Art. 1º- É declarada Árvore Nacional a leguminosa denominada Pau-Brasil (Caesalpinia echinata, Lam), cuja festa será comemorada, anualmente, quando o Ministério da Educação e Cultura promoverá campanha elucidativa sobre a relevância daquela espécie vegetal na História do Brasil."
- (en) Boden, Anne, « Golden Wattle: Floral Emblem of Australia », Australian National Botanic Gardens, (lire en ligne [http], consulté le ).
- (en) Anne Boden, « Floral Emblems - Australian Plant Information », sur Floral emblems of Australia, Australian National Botanic Gardens, Parks Australia, (consulté le ).
- (en) Williams, Martin, « Golden Enigmatic Beauty », Bahuninia, (lire en ligne [http], consulté le ).
- (zh) 珠海 sur Baidu.com
- (en) « Texas State Flower », sur State Symbols USA (consulté le ).
- (en) Victoria Hughes, « Red Clover », sur Vermont History Explorer (consulté le ).
- (en) « Arizona State Tree », sur statesymbolsusa.org (consulté le ).
- (en) Thomas E. Marler & John H. Lawrence, « Ifit: Intsia bijuga - the territorial tree of Guam », Guam Department of Agriculture, Forestry and Soil Resources Division, (consulté le ).
- (en) « Flamboyant - Delonix regia - Extant & Habitat resource - Details », sur Encyclopedia of Life (consulté le ).
- (en) « How We Are Governed, St. Kitts & Nevis - National Symbols », sur www.stkittsnevis.net (consulté le ).
- (en) « Oklahoma State Tree - Redbud », sur State Symbols USA (consulté le ).
- (en) « Halland », sur Swedensite (consulté le ).
- (en-US) « Cassia fistula L. – MSSBG », (consulté le ).
- (en) « State Symbols », sur Telangana State Portal (consulté le ).
- « L'ajonc devient la plante emblème de la Bretagne », Ouest-France, (lire en ligne).
- (pt) Rodrigo L. S. Aguiar, « Arqueologia e História indígena no Litoral de Santa Catarina », sur Academia, (consulté le ).
- (en) « The truth behind the shamrock. », sur BBC News,
Voir aussi
Bibliographie
- Fabuleuses légumineuses - Actes du colloque SNHF - 2016
- Rôle des légumineuses sur la fertilité des sols ferrugineux tropicaux des zones guinéenne et soudanienne du Burkina Faso Thèse - Boubie Vincent .
- Normes Codex pour les Céréales, légumes secs, légumineuses et matières protéiques végétales, Première édition, FAO/OMS, 2007, (ISBN 978-92-5-205842-7), 128 p., consultable sur le site de la FAO[PDF].
- (en) O N Allen et E K Allen, The Leguminosae, University of Wisconsin Press/Macmillan Publishing Company, , 812 p.
- (en) Wojciechowski MF, Lavin M, Sanderson MJ, « A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family », American Journal of Botany, vol. 91, no 11, , p. 1846-1862 (DOI 10.3732/ajb.91.11.1846).
Articles connexes
Liens externes
- Ressources relatives au vivant :
- Global Biodiversity Information Facility
- Tela Botanica
- VASCAN
- (en) Australian Plant Name Index
- (sv) Dyntaxa
- (en) EPPO Global Database
- (en) Flora of China
- (en) FloraBase
- (en) Paleobiology Database
- (en) Germplasm Resources Information Network
- (mul + en) iNaturalist
- (en) Interim Register of Marine and Nonmarine Genera
- (en) International Plant Names Index
- (en) NBN Atlas
- (nl) Nederlands Soortenregister
- (en + en) New Zealand Organisms Register
- (ru) Plantarium
- (en) Plants of the World Online
- (en) Plazi
- (en) Système d'information taxonomique intégré
- (en) Tropicos
- (en) VicFlora
- (en) World Register of Marine Species
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- (fr) Référence Belles fleurs de France : Fabaceae
- (fr) Référence Belles fleurs de France 2 : Fabaceae
- (fr) Référence Tela Botanica (France métro) : Fabaceae
- (en) Référence Flora of China : Fabaceae
- (en) Référence Madagascar Catalogue : Fabaceae
- (en) Référence Flora of Chile : Fabaceae
- (en) Référence Angiosperm Phylogeny Website : Fabaceae ou Leguminosae
- (en) Référence Jardin botanique du Missouri : Fabaceae (images de Madagascar)
- (en) Référence Tree of Life Web Project : Fabaceae (consulté le )
- (en) Référence Catalogue of Life : Fabaceae (consulté le )
- (fr+en) Référence ITIS : Fabaceae stricto sensu
- (en) Référence NCBI : Fabaceae (taxons inclus) lato sensu
- (en) Référence GRIN : famille Fabaceae Lindl. (+liste des genres contenant des synonymes)
- (en) Référence DELTA Angio : Leguminosae Fabaceae lato sensu