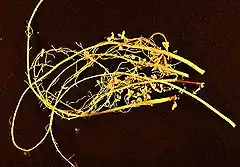
Nodosité
Les nodosités ou nodules sont des modifications des poils absorbants des racines, que certaines plantes développent pour améliorer leur nutrition d'azote sous l'action de certaines bactéries. Ces bactériocécidies se forment, sous l'action de bactéries symbiotiques fixatrices de l'azote atmosphérique, essentiellement sur les racines de nombreuses espèces de plantes (Fabacées appelées aussi Légumineuses, Ulmacées, Bétulacées, Casuarinacées, Eleagnacées, Myricacées, Rosacées, etc.), mais aussi sur leurs tiges (Aeschynomene et Sesbania rostrata de la famille des Fabacées) ou sur leurs feuilles (quelques genres chez les Rubiacées, Myrsinacées, Dioscoreacées)[1].
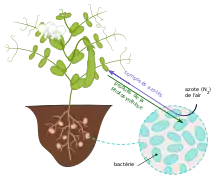
Dans cette symbiose très spécialisée, la plante fournit les substances carbonées et les bactéries les substances azotées synthétisées à partir de l'azote atmosphérique.
Les bactéries du genre Rhizobium et Bradyrhizobium forment des symbioses rhizobiennes. Les nodosités élaborées par l'actinomycète du genre Frankia sont appelées actinorhizes.
Dans les nodosités, les bactéries symbiotiques sont capables, grâce à leur synthèse de l'enzyme nitrogénase dont les plantes eucaryotes sont dépourvues, de convertir l’azote atmosphérique (N2) en ammoniac (NH3) assimilable par la plante hôte alors que séparément ces organismes ne peuvent pas utiliser l’azote présent dans l’air. L’ammoniac peut alors être utilisé pour produire des acides aminés (utilisés pour la synthétisation de protéines), nucléotides (utilisées pour la synthétisation d’ADN et ARN) et autres composés cellulaires essentiels à la plante. En échange de l’azote fixé par les bactéries, la plante colonisée fournit du malate provenant de sa photosynthèse, comme source de carbone qui servira au métabolisme du diazote en ammoniac[2]. La couleur rouge typique des nodosités est due à la présence de leghémoglobine, une hémoprotéine fixatrice de dioxygène (O2) qui compose 40 % du contenu protéique de ces structures. Alors que les rhizobia sont sensibles à l’oxygène, elles en ont tout de même besoin pour obtenir l’ATP nécessaire à la fixation de l’azote. L’heghémoglobine permet cette diffusion d’oxygène sans qu’il soit libre dans les cellules et donc toxique pour les bactéries[3]. Des nodosités vertes sont signes d’un problème de symbiose et sont incapables de fixer l’azote[4].
La fixation de l’azote au niveau des nodosités est un service écologique utile en agriculture. Cela permet de réduire l’utilisation de fertilisant azoté et de réduire les coûts monétaires et environnementaux de la culture de légumineuses, en particulier le soya.

Étapes du développement
Les activités coordonnées d'une légumiseuse et d'une bactérie du genre Rhizobium dépendent des réactions chimiques entre les deux partenaires symbiotiques.
Première étape : pénétration
Les racines sécrètent des substances chimiques de type flavonoïde (chrysoériol et lutéoline chez la luzerne, apigénine et ériodictyol chez le pois) et isoflavanoïde (daidzéine chez le soja), qui attirent les bactéries de type Rhizobium du voisinage[5]. En réponse, les bactéries synthétisent et émettent des facteurs de nodulation, dits facteurs nod. Sous leurs action, les poils absorbants changent leur direction de croissance et forment une structure en crosse de berger, qui enferme les rhizobiums. Les bactéries peuvent ainsi pénétrer dans l'écorce (parenchyme cortical), via un filet infectieux. Au même moment, la racine commence à répondre à cette pénétration par une division des cellules de l'écorce et du péricycle du cylindre central. Les vésicules contenant les bactéries bourgeonnent dans les cellules de l'écorce à partir de l'extrémité du filet infectieux ramifié.
Deuxième étape : développement de la nodosité
La croissance se poursuit dans les régions infectées de l'écorce et du péricycle, jusqu'à ce que ces deux masses de cellules fusionnent et forment la nodosité. Les bactéries provoquent des invaginations de la membrane plasmique qui vont fusionner avec l'appareil de Golgi, ce qui permet l'avancée de la bactérie vers le cortex racinaire.
Troisième étape : maturation de la nodosité
La nodosité continue sa croissance, alors que le tissu conducteur reliant la nodosité au xylème et au phloème du cylindre central se développe. Ce tissu conducteur apporte à la nodosité les glucides et autres substances organiques nécessaires au métabolisme cellulaire. Il transporte aussi les composés azotés produits dans la nodosité vers le cylindre central, qui les distribuera dans toute la plante. On ne parle donc pas d'infection mais de symbiose, chaque individu étant bénéfique à l'autre.
Bénéfices

En plus de la fixation d'azote, notamment avec les Fabaceae (autrefois légumineuses) par les bactéries des genres genere Rhizobium, Sinorhizobium, Bradyrhizobium, Mesorhizobium, Azorhizobium et Allorhizobium[6], ainsi que certains angiospermes, par actinorhize, principalement avec les bactéries Frankia, il y aura plus ou moins de bénéfices sur la production de la plante elle même.
Par exemple, sur 1 hectare (10 000 m²) de lupin, 2,2 tonnes sont produites sans nodosité, contre 4,2 tonnes avec.
Voir aussi
Articles connexes
Références
- (en) S.M. Reddy, S. Girisham, G. Narendra Babu, Applied Microbiology (Agriculture, Environmental, Food and Industrial Microbiology), Scientific Publishers, , p. 34-37
- Jean-François Morot-Gaudry, Assimilation de l'azote chez les plantes : aspects physiologique, biochimique et moléculaire, Inra, , 424 p.
- Singh, S, « Structure, Function, and Estimation of Leghemoglobin. », Soil Biology, , p. 309-330
- Becana, M., « Structure and function of leghemoglobins. », Anales de la Estación Experimental de Aula Dei, , p. 203-208
- Jean-Jacques Macheix, Annie Fleuriet, Christian Jay-Allemand, Les composés phénoliques des végétaux, Presses polytechniques et universitaires romandes, , p. 68-69.
- (en) Alfonso González-Sama, M. Mercedes Lucas, María R. de Felipe et José J. Pueyo, « An Unusual Infection Mechanism and Nodule Morphogenesis in White Lupin (Lupinus albus) », Pueyo The New Phytologist, vol. 163, no 2, 2004), p. 371-380 (DOI 10.1111/j.1469-8137.2004.01121.x, JSTOR 1514387, lire en ligne)
Bibliographie
- Bergersen, F.J. (1980). Methods for evaluating biological nitrogen fixation. New Jersey: John Wiley & Sons Ltd. 712 p.
- Elkan, G. H. (1987). Symbiotic nitrogen fixation technology. New York: Marcel Dekker, Inc. 464 p.
- Howieson, J. G. and Dilworth, M. J. (2016). Working with rhizobia. Canberra: Australian Centre for International Agricultural Research. 312 p.
- Arora, N. K. (2015). Plant Microbes Symbiosis: Applied Facets. New Delhi: Springer. 381 p.
- Lal, R. and Stewart, B. A. (2018). Soil Nitrogen Uses and Environmental Impacts. Boca Raton: CRC Press. 379 p.
- Boogerd, F. C. and Rossum, D. (1997). Nodulation of groundnut by Bradyrhizobium: a simple infection process by crack entry. FEMS Microbiology Reviews, 21, 5-27.
- Raven, P. H., Evert, R. F. et Eichhorn, S. E. (2014). Biologie végétale, 3e Édition. Paris: de Boeck. 880 p.