Fixation biologique du diazote
La fixation biologique de l'azote, ou diazotrophie (du grec di, « deux », azo, « azote », et trophos, « alimentation »), est le processus du cycle de l'azote qui permet à un organisme de produire (indirectement) des substances protéiques à partir de l'azote gazeux (N2) présent dans l'atmosphère et l'environnement.
L'acronyme BNF est parfois utilisé par les anglophones pour résumer l'expression « biological N2-fixation ».
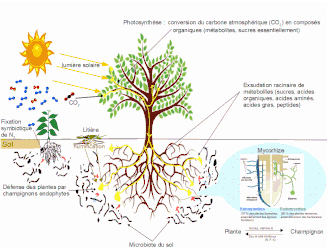

Elle peut être symbiotique lorsqu'elle est liée à la formation de nodosités, ou asymbiotique lorsqu'elle implique des bactéries diazotrophes qui restent libres dans le sol ou en association plus ou moins étroite avec les plantes, comme colonisateurs épi- ou endophytique[1]. La fixation par les bactéries symbiotiques, plus efficace, constitue le processus naturel dominant de fixation de l'azote dans la majorité des écosystèmes terrestres, permettant aux espèces pionnières de coloniser des milieux pauvres en azote. La fixation asymbiotique de l'azote qui joue un rôle important dans les milieux gorgés d'eau et les déserts, est estimée de 23 % à 32 % du total d'azote assimilé par les plantes[2].
Cette stratégie d'acquisition de nutriment met en jeu le processus de réduction enzymatique de N2 (azote moléculaire) en azote ammoniacal, ou ammoniac (NH3) : cette forme de N combiné, appelée intermédiaire-clé, représente la fin de la réaction de fixation et le début de l'incorporation de l'azote fixé dans le squelette carboné. Dans le système biologique fixateur de N2 les conditions optimales de la catalyse biologique correspondent à une pression de 0,2 à 1,0 atm de N2 et une température de 30-35 °C, alors que les conditions de la catalyse industrielle sont très sévères : pression de 250-1000 atm de N2 et température de 450 °C[3].
Généralités
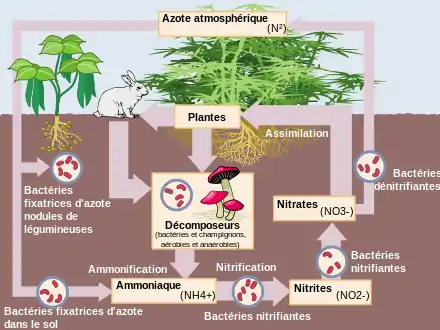
Ce processus est comparable à celui de la photosynthèse qui permet de produire des substances glucidiques à partir du dioxyde de carbone (CO2) de l'atmosphère. Mais, alors que la photosynthèse est le fait de tous les végétaux (sauf les végétaux saprophytes), la fixation de l'azote ambiant n'est réalisée que par certaines espèces de bactéries comme les cyanophycées. Toutefois, de nombreuses plantes, principalement de la famille des Fabaceae (légumineuses), la réalisent de façon indirecte, en symbiose avec des bactéries de leur rhizosphère, qui se localisent généralement dans des nodosités situées sur leurs racines.
Jusqu'à une date récente, on admettait que les champignons mycorrhiziens pouvaient fixer N2. On sait maintenant que la propriété de fixer N2 est strictement limitée aux procaryotes[4] - [5] et n’a jamais été montrée chez les champignons filamenteux. La fixation de N2 mesurée par la réduction de l’acétylène (C2H2) en éthylène (C2H4 ou CH2=CH2) par une racine mycorrhizée ne devrait pas être imputée au champignon lui-même mais aux bactéries associées de la mycorrhizosphère. Cette activité de fixation de N2 est d'ailleurs encore plus importante dans la litière forestière que dans la mycorrhizosphère elle-même[6].
Ces organismes procaryotes (bactéries) produisent une enzyme, la nitrogénase, qui permet de réaliser la synthèse de l'ammoniac par une réaction de réduction fortement exergonique et endothermique (en raison de la très forte énergie de la liaison triple de N2, 940,5 kJ/mol, elle exige 16 ATP pour la fourniture de l'énergie d'activation)[7] - [8] :
dans les conditions du sol, à l'opposé des 450 °C des procédés industriels qui requièrent aussi une pression moyenne de l'ordre de 400 atmosphères. L'ammoniac est ensuite transformé en acides aminés utilisables par les plantes.
La nitrogénase est en réalité un complexe enzymatique formé de sous-unités, dont le fonctionnement est in vitro inhibée par l'oxygène. Mais dans le processus de symbiose son fonctionnement est rendu possible en zone de microaérobiose (dans les nodules racinaires) voire en condition aérobie à l'intérieur des racines de plantes non légumineuses (avec l'aide de microorganismes dites « endophytes » selon Santi & al en 2013[9] qui semble ainsi bénéficier de condition [10] - [11].
Par ailleurs les micro-organismes diazotrophes (fixatrices d'azote) ont de leur côté développé plusieurs stratégies de protection de leur complexe nitrogénase[12] - [13].
L’inhibition de la nitrogénase par l'O2, que ce soit directement ou indirectement via des radicaux oxydants, constitue un régulateur fort de la teneur en oxygène dans l'atmosphère, dans le cycle de l'oxygène, par la rétroaction négative qu'il a sur la fixation de l'azote[14]. Une inhibition dans la fixation de l'azote conduirait à une chute de la productivité primaire, à la fois directement pour les cyanobactéries diazotrophes et indirectement pour l'ensemble de la productivité primaire de l'hydrosphère, induisant à plus long terme, pour le cycle du carbone, une réduction dans le rythme d'enfouissement du carbone organique. Sans enfouissement il n'y a pas de création nette d'O2 par la biosphère, et l'oxygène en excès espèce très réactive, disparaît progressivement par l'oxydation de la surface de la planète.
Les bactéries fixatrices d'azote
Il existe des bactéries libres qui vivent dans le sol et assurent la fixation de l'azote, soit seules, soit en symbiose avec d'autres bactéries. Ce sont souvent des bactéries auxiliaires de la mycorhization (en), d'où leur forte densité au niveau de la rhizosphère où elles peuvent recevoir les apports carbonés des exsudats racinaires. Elles sont caractérisées par la fixation asymbiotique de l'azote (l'association avec les racines n'est pas étroite alors que la symbiose au sens strict implique une interaction très intime). D'autres fixent l'azote atmosphérique, en relation plus ou moins étroite avec les plantes, comme colonisateur épi- ou endophytique[15]. Elles présentent une grande diversité de modes de productions d'énergie : métabolisme respiratoire aérobie (Azotobacter, Bacillus, cyanobactéries du genre Nostoc, Anabaena, Calothrix), associée à différentes stratégies pour éviter que leur nitrogénase soit en contact avec l'oxygène (augmentation de l'activité respiratoire pour réduire le taux d'oxygène environnant, développement d'hétérocystes) ; métabolisme anaérobie obligatoire (Clostridium, Chlorobium, Chromatium) ou facultative (Aerobacter, Klebsiella, Pseudomonas) ; métabolisme microaérophile (Azospirillum)[16] - [17].
D'autres espèces bactériennes développent une association symbiotique hautement spécialisée avec le système racinaire des plantes, faisant intervenir la création d'un nouvel organe, la nodosité :
- groupe des Rhizobia (Rhizobium, Bradyrhizobium, Mesorhizobium (en)…) : légumineuses (fabacées) et ulmacées ;
- Frankia : diverses espèces d'angiospermes, essentiellement arbres et arbustes (au sein des Casuarinacées, Eleagnacées, Myricacées, Rosacées, etc.), notamment les aulnes, l'argousier, les Casuarinaceae et le Myrica gale.
Dans les océans, ce sont les Cyanobacteria qui fixent le diazote. Il en existe deux types :
- Les cyanobactéries dites « filamenteuses », telles que Trichodesmium sp., qui vivent en colonie ;
- Les cyanobactéries unicellulaires, appelées UCYN, qui peuvent vivre libre ou en symbiose avec certaines espèces du phytoplancton.
Le cas des rhizobiums chez les légumineuses
Les différentes espèces de bactéries rhizobiums, qui sont capables d'infecter les racines des légumineuses (famille des Fabacées), sont plus ou moins spécifiques de certaines plantes-hôtes. Ainsi Rhizobium phaseoli infecte les haricots (Phaseolus sp).
Les bactéries provoquent la formation de nodosités sur les racines en pénétrant par les poils racinaires, et se transforment en « bacteroides » de plus grande taille. Les nodosités sont le siège d'une activité symbiotique dans laquelle la plante fournit les sucres et l'énergie issus de la photosynthèse, et bénéficie en retour des acides aminés qui y sont produits.
Cette activité peut produire annuellement jusqu'à 300 kg d'azote à l'hectare, qui se retrouvent en partie dans les récoltes exportées (protéines des graines et fourrages) et en partie enfoui dans le sol, utilisable par les cultures suivantes (engrais vert).
Au niveau mondial, on estime à 100 millions de tonnes par an la masse d'azote ainsi fixé, soit le même ordre de grandeur que la production d'azote de l'industrie chimique.
Le cas des cyanobactéries
Les cyanobactéries ont établi au cours de l'évolution des végétaux, des symbioses avec un large spectre de plantes hôtes : bryophytes, ptéridophytes, lichens, gymnospermes (cycas), angiospermes (Gunnera)[18].
Dialogue fabacées-rhizobiacés
La formation des nodules racinaires met en jeu un dialogue moléculaire complexe entre la plante-hôte et les rhizobiacées.
La plante sécrète des signaux de type flavonoïdes, bétaïnes. La perception de ces signaux par la bactérie induit l’expression de nombreux gènes bactériens, dont une centaine sont connus. Les plus étudiés sont les gènes nod (ou bien nol et noe). Les gènes Nod D expriment des protéines régulatrices en réponse aux inducteurs végétaux. Une fois activées les protéines Nod D se fixent sur des séquences nod-box, promoteur des gènes nod de structure et activent leur transcription. Ces gènes produisent des facteurs Nod essentiels dans l’établissement de la symbiose.
Les facteurs NOD activent une voie de signalisation faisant intervenir une série de gènes de la plante-hôte, et provoquent finalement des modifications morphologiques et cytologiques de la racine, pour aboutir à la formation des nodules.
La voie de signalisation NOD
De nombreux gènes ont été identifiés chez le pois (Pisum sativum), le lotier (Lotus japonica) et la luzerne faux-tribule (Medicago truncatula). Ce sont tous des récepteurs SYMRK (symbiosis receptor like kinase), possédant un domaine kinase côté intracellulaire (RLK, Receptor like kinase) et des motifs riches en LysmM (interaction avec les facteurs NOD) côté extracellulaire. Les gènes sont appelés Nfr (Nod factor receptor) ou Sym (symbiosis genes). Il pourrait y avoir deux types de récepteurs, un contrôlant l’infection de la racine par la bactérie, l’autre permettant la courbure des poils absorbants.
Quelques dizaines de secondes après l’application des facteurs Nod, on observe un influx rapide de calcium qui induit une dépolarisation de la membrane plasmique entrainant la courbure et la déformation de poils absorbants.
Dix minutes après des oscillations périodiques et transitoires de la concentration de calcium cytosolique se produisent à proximité de la région périnucléaire. Ces oscillations calciques (calcium spiking) sont dues à deux protéines membranaires DMI1 et DMI2. Ce dernier est un récepteur de type RKL (receptor like kinase) présentant trois domaines riches en Leucine(LRR).
La réponse à ces oscillations est due au gène DMI3 codant une protéine kinase calmoduline et calcium dépendante. Cette protéine perçoit les variations de concentration en calcium à la fois par liaison à des calmodulines liées au calcium et au calcium libre. DMI3 est activé par l’autophosphorylation de son domaine kinase. De plus, DMI3 exerce un retrocontrôle négatif sur DMI1 et DMI2, et donc sur les oscillations calciques.
DMI3 active l’expression des gènes ENOD (early noduline genes, gènes à nodulines précoces), protéines produites par la plante pendant les premières étapes de la symbiose, (comme la leghemoglobine) impliquées dans l’infection et l’organogenèse des nodules activant la division des cellules corticales. Cependant les substrats intermédiaires de DMI3 ne sont pas encore bien déterminés.
Évolution des symbioses fixatrices d’azote
Origine et apparition
Il semblerait que les rhizobiacées ont recruté la voie de signalisation des mycorhizes et que l'organogénèse du nodule soit dérivée du programme génétique de l'organogénèse des racines secondaires. Les endosymbioses fixatrices d'azote seraient apparues il y 65 millions d'années. L’ancêtre commun à toutes ces espèces aurait développé une endosymbiose bactérienne, grâce à une certaine prédisposition à l’organogénèse nodulaire (la formation de nodules étant un détournement du programme génétique de formation des racines secondaires). Il est probable que la capacité à former des nodosités soit apparue plusieurs fois et indépendamment au cours de l'évolution. Cette aptitude a aussi été perdue plusieurs fois, par exemple pour l’adaptation à des sols plus fertiles. Cela permet d'expliquer la présence de plantes sans nodules à l'intérieur du clade des plantes à nodules.
Spécificité
Deux « forces de sélection » antagonistes pourraient rendre compte de la spécificité des symbioses fixatrices d'azote.
- La première de ces forces, positive, est l'augmentation du taux d'azote fixé par la plante hôte colonisée par le micro-organisme fixateur d'azote.
- La seconde, négative, est l'invasion de la plante hôte par un pathogène non productif.
Une spécificité faible semble se rencontrer dans les genres de plantes primitives, alors que la spécificité des symbiotes augmente au cours de l'évolution.
Au niveau moléculaire on peut définir trois niveaux de spécificité lors du dialogue plante hôte rhizobium. Les gènes Nod sont portés par un plasmide bactérien. Des transferts horizontaux de gènes peuvent alors s'effectuer, ce qui transfère également les spectres d'hôtes. Ceci explique l'interaction entre des espèces de bactéries éloignées évolutivement et des plantes hôtes proches, et inversement. Ainsi il y a une coévolution entre les gènes de la symbiose et les plantes hôtes, et non entre les rhizobiacées et les plantes hôtes.
Autres plantes à la rhizosphère fixatrice d'azote

De nombreuses plantes ont des associations similaires :
- Allocasuarina ainsi que d'autres genres de la famille des Casuarinaceae
- Alnus ou Aulne (famille des Betulaceae)
- Azolla (famille des Azollaceae)
- Casuarina (famille des Casuarinaceae)
- Ceanothus (famille des Rhamnaceae)
- Cercocarpus (famille des Rosaceae)
- Cycadacée (famille des Cycadaceae)
- Elaeagnus umbellata (famille des Elaeagnaceae)
- Gunnera (famille des Gunneraceae)
- Lobaria - Lobaria cuneifolia (L.) Haw.; nom retenu: Saxifraga cuneifolia subsp. cuneifolia[20], Désespoir-du-peintre, Saxifrage à feuilles en coin (famille des Saxifragaceae) avec certains lichens.
- Myrica (famille des Myricaceae)
- Purshia tridentata (angl. antelope bitterbrush) (famille des Rosaceae)
- Robinia pseudoacacia, Leucaena leucocephala (famille des Fabaceae)
- Shepherdia argentea (angl. silver buffaloberry) (famille des Elaeagnaceae)
Plantes disposant de microorganismes « endophytes »
Depuis les années 2000 à 2015, plusieurs espèces végétales présentant des endophytes fixateurs d'azote ont été trouvées, dont :
- Azoarcus sp (herbacée) [21] ;
- Canne à sucre[22] ;
- riz sauvage[23] - [24] ;
- maïs[25],
- Sorghum halepense[26] ;
- Miscanthus[27] - [28] ;
- cactus Mammillaria fraileana[29] ;
- Patate douce[30] - [31] ;
- mousses des forêts boréales[32] ;
- herbacées dunaires (Ammophila arenaria et Elymus mollis[33] ;
- plants de café Coffea arabica L.[34] ;
- plants de conifères (Pinus flexilis et Picea engelmannii ) inoculé avec Paenibacillus polymyxa[35] - [36].
La capacité de fixation de l'azote a été directement quantifiés chez quelques graminées (canne à sucre, blé et riz[37] - [21] - [38] - [39]).
Mesure de la fixation biologique du diazote
Elle se fait par deux moyens :
- un premier test utilise le fait qu'il existe un isotope stable de l'azote : l'isotope 15N2 qui est chimiquement inerte et naturellement rare dans l'atmosphère (bien plus rare que l'autre isotope de l'azote : 14N). La mesure du taux d'incorporation de cet isotope dans les tissus vivants d'une plante est la méthode la plus directe de dosage de la fixation biologique du diazote (N2). Si une plante ou un tissu végétal contient plus cet isotope qu'un autre, c'est qu'il a bénéficié d'un moyen de capter des quantités plus importantes de diazote de l'air et de le transformer en azote moléculaire biodisponible (pour produire des acides aminés ou des protéines). Même une faible augmentations de l'isotope 15N par rapport à une plante-témoins indique une fixation de N2[40]. Il est possible pour les besoins d'expériences de laboratoire d'acheter des bombonnes d'azote 15N2 comprimé (98 % des atomes sont des isotopes 15N2)[41] ;
- un autre moyen (dit « ARA » par les anglophones ; acronyme de « Acetylene reduction assay ») est un « test de réduction de l'acétylène » ; c'est une évaluation cette fois indirecte de la fixation de l'azote atmosphérique N2[42].
Certaines études, par précaution, utilisent à la fois ces deux tests, comme l'étude de 2016 qui a pour la première fois montré l'existence de bactéries diazotrophes symbiotes d'espèces pionnières dans les branches d'arbre (saules ou peupliers pionniers s'épanouissant sur des milieux fortement carencés en azote)[41].
Notes et références
- (en) R. M. Boddey & J. Dobereiner, « Nitrogen fixation associated with grasses and cereals: Recent results and perspectives for future research », Plant and Soil, vol. 108, , p. 53–65.
- (en) K. J.Chaitanya, S. Meenu., « Plant growth promoting Rhizobacteria (PGPR): a review », E3 J. Agric.Res. Develop., vol. 5, no 2, , p. 110.
- R.W.F. Hardy, E.Jr. Knight. 1968. The biochenistry and postulated mechanisms of N2 fixation. In "Progress in Phytochemistry" (L. Reinhold, ed.), 387-469. Cité dans "Mycorrhizes et fixation d’azote" du 25 avril 1978 ; Y. Dommergues, O.R.S.T.O.M.
- G.D. Bowen 1973. Mineral nutrition of octomycorrhizae. Ectomycorrhyzae. Their Ecology and Physiology. G.C. Marks and T.T. Kozlowski, ed., Academic Press, New York and London, 151-197. Cité dans Dommergues 1978.
- J.R. Postgate. 1974. Evolution within nitrogen-fixing systems. Symposia of the Society for General Microbiology, 24, 263-292. Cité dans Dommergues 1978.
- W.B. Silvester, K.J. Bennett. 1973. Acetylene reduction by roots and associated soil of new zealand conifers. Soil Biol. Biochem, 5, 1’11-179. Cité dans Dommergues 1978
- Duhoux et Nicole 2004 (Dunod)
- Lee Chi Chung, Ribbe Markus W. et Hu Yilin, The Metal-Driven Biogeochemistry of Gaseous Compounds in the Environment, vol. 14, Springer, coll. « Metal Ions in Life Sciences », , 333 p. (ISBN 978-94-017-9268-4, PMID 25416393, DOI 10.1007/978-94-017-9269-1_6), « Chapter 7. Cleaving the N,N Triple Bond: The Transformation of Dinitrogen to Ammonia by Nitrogenases »
- Santi C, Bogusz D, Franche C (2013) Biological nitrogen fixation in non-legume plants. Ann Bot 111: 743–767. mct048 [pii]; doi: 10.1093/aob/mct048.
- Sessitsch A, Hardoim P, Doring J, Weilharter A, Krause A, Woyke T et al. (2012) Functional characteristics of an endophyte community colonizing rice roots as revealed by metagenomic analysis. Mol Plant Microbe Interact 25: 28–36. doi: 10.1094/MPMI-08-11-0204.
- ames EK, Olivares FL (1998) Infection and colonization of sugarcane and other graminacious plants by endophytic diazotrophs. Crit Rev Plant Sci 17: 77–119. doi: 10.1080/07352689891304195
- Kathleen Marchal et Jos Vanderleyden, « The "oxygen paradox" of dinitrogen-fixing bacteria », Biol Fertil Soils, vol. 30, , p. 363–373 (DOI 10.1007/s003740050017, lire en ligne)
- Boyd ES, Costas AM, Hamilton TL, Mus F, Peters JW, « Evolution of molybdenum nitrogenase during the transition from anaerobic to aerobic metabolism », J Bacteriol, vol. 197, , p. 1690–1699. JB.02611-14 [pii] (PMID 25733617, DOI 10.1128/JB.02611-14, lire en ligne)
- Feedbacks Between the Nitrogen, Carbon and Oxygen Cycles. Ilana Berman-Frank et al. In Nitrogen in the Marine Environment, 2008, Elsevier.
- (en) Carole Santi, Didier Bogusz, Claudine Franche, « Biological nitrogen fixation in non-legume plants », Ann Bot, vol. 111, no 5, , p. 743-767 (DOI 10.1093/aob/mct048).
- (en) Geeta Sumbali, R S Mehrotra, Principles of Microbiology, Tata McGraw-Hill, , p. 754-755.
- Jean-François Morot-Gaudry, Loïc Lepiniec, Roger Prat (dir.), Biologie végétale : Croissance et développement, Dunod, , p. 242-244.
- Lydie Suty, Les végétaux. Des symbioses pour mieux vivre, Quae, , p. 18
- (en) Allen Van Deynze et al., « Nitrogen fixation in a landrace of maize is supported by a mucilage-associated diazotrophic microbiota », PLoS Biol, vol. 16, no 8, (DOI 10.1371/journal.pbio.2006352).
- Tela-Botanica
- Hurek T, Handley LL, Reinhold-Hurek B, Piche Y (2002) Azoarcus grass endophytes contribute fixed nitrogen to the plant in an unculturable state. Mol Plant Microbe Interact 15: 233–242. doi: 10.1094/mpmi.2002.15.3.233
- [16]Welbaum GE, Sturz AV, Dong Z, Nowak J (2003) Endophytic nitrogen fixation in sugarcane: present knowledge and future applications. Plant Soil 252: 139–149. doi: 10.1023/a:1024152126541
- Peng G, Zhang W, Luo H, Xie H, Lai W, Tan Z (2009) Enterobacter oryzae sp. nov., a nitrogen-fixing bacterium isolated from the wild rice species Oryza latifolia. Int J Syst Evol Microbiol 59: 1650–1655. doi: 10.1099/ijs.0.65484-0.
- Elbeltagy A, Nishioka K, Sato T, Suzuki H, Ye B, Hamada T et al. (2001) Endophytic colonization and in planta nitrogen fixation by a Herbaspirillum sp. isolated from wild rice species. Appl Environ Microbiol 67: 5285–5293. doi: 10.1128/aem.67.11.5285-5293.2001
- Montanez A, Abreu C, Gill PR, Hardarson G, Sicardi M (2009) Biological nitrogen fixation in maize (Zea mays L.) by N-15 isotope-dilution and identification of associated culturable diazotrophs. Biology and Fertility of Soils 45: 253–263. doi: 10.1007/s00374-008-0322-2
- Rout ME, Chrzanowski TH (2009) The invasive Sorghum halepense harbors endophytic N2-fixing bacteria and alters soil biogeochemistry. Plant and Soil 315: 163–172. doi: 10.1007/s11104-008-9740-z
- Davis SC, Parton WJ, Dohleman FG, Smith CM, Del Grosso S, Kent AD et al. (2010) Comparitive biogeochemical cycles of bioenergy crops reveal nitrogen-fixation and low greenhouse gas emissions in a Miscanthus x giganteus agro-ecosystem. Ecosystems 13: 144–156. doi: 10.1007/s10021-009-9306-9
- Videira SS, Pereira e Silva MC, Galisa PS, Dias ACF, Nissinen R, Divan VLB et al. (2013) Culture-independent molecular approaches reveal a mostly unknown high diversity of active nitrogen-fixing bacteria associated with Pennisetum purpureum—a bioenergy crop. Plant Soil 373: 737–754. doi: 10.1007/s11104-013-1828-4
- [23]Lopez BR, Tinoco-Ojanguren C, Bacilio M, Mendoza A, Bashan Y (2012) Endophytic bacteria of the rock-dwelling cactus Mammillaria fraileana affect plant growth and mobilization of elements from rocks. Environ Exper Botany 81: 26–36. doi: 10.1016/j.envexpbot.2012.02.014
- Reiter B, Burgmann H, Burg K, Sessitsch A (2003) Endophytic nifH gene diversity in African sweet potato. Canadian Journal of Microbiology 49: 549–555. doi: 10.1139/w03-070
- Terakado-Tonooka J, Fujihara S, Ohwaki Y (2012) Possible contribution of Bradyrhizobium on nitrogen fixation in sweet potatoes. Plant and Soil 367: 639–650. doi: 10.1007/s11104-012-1495-x
- DeLuca TH, Zackrisson O, Gundale MJ, Nilsson MC (2008) Ecosystem feedbacks and nitrogen fixation in boreal forests. Science 320: 1181. 320/5880/1181 [pii]; doi: 10.1126/science.1154836.
- Dalton DA, Kramer S, Azios N, Fusaro S, Cahill E, Kennedy C (2004) Endophytic nitrogen fixation in dune grasses (Ammophila arenaria and Elymus mollis) from Oregon. FEMS Microbiol Ecol 49: 469–479. doi: 10.1016/j.femsec.2004.04.010.
- Jimenez-Salgado T, Fuentes-Ramirez LE, Tapia-Hernandez A, Mascarua-Esparza MA, Martinez-Romero E, Caballero-Mellado J (1997) Coffea arabica L., a new host plant for Acetobacter diazotrophicus, and isolation of other nitrogen-fixing acetobacteria. Applied and Environmental Microbiology 63: 3676–3683.
- Bal A, Chanway CP (2012) Evidence of nitrogen fixation in lodgepole pine inoculated with diazotrophic Paenibacillus polymyxa. Botany 90: 891–896. doi: 10.1139/b2012-044
- Carrell AA, Frank AC (2014) Pinus flexilis and Picea engelmannii share a simple and consistent needle endophyte microbiota with a potential role in nitrogen fixation. Front Microbiol 5: 333. doi: 10.3389/fmicb.2014.00333.
- Iniguez AL, Dong Y, Triplett EW (2004) Nitrogen fixation in wheat provided by Klebsiella pneumoniae 342. Mol Plant Microbe Interact 17: 1078–1085. doi: 10.1094/mpmi.2004.17.10.1078
- Elbeltagy A, Nishioka K, Sato T, Suzuki H, Ye B, Hamada T et al. (2001) Endophytic colonization and in planta nitrogen fixation by a Herbaspirillum sp. isolated from wild rice species. Appl Environ Microbiol 67: 5285–5293. doi: 10.1128/aem.67.11.5285-5293.2001
- Sevilla M, Burris RH, Gunapala N, Kennedy C (2001) Comparison of benefit to sugarcane plant growth and 15N2 incorporation following inoculation of sterile plants with Acetobacter diazotrophicus wild-type and Nif- mutant strains. Molecular Plant Microbe Interactions 14: 358–366. doi: 10.1094/mpmi.2001.14.3.358
- Pinto-Tomas AA, Anderson MA, Suen G, Stevenson DM, Chu FS, Cleland WW et al. (2009) Symbiotic nitrogen fixation in the fungus gardens of leaf-cutter ants. Science 326: 1120–1123. 326/5956/1120 [pii]; doi: 10.1126/science.1173036.
- Doty SL, Sher AW, Fleck ND, Khorasani M, Bumgarner RE, Khan Z, et al. (2016) Variable Nitrogen Fixation in Wild Populus. PLoS ONE 11(5): e0155979. doi:10.1371/journal.pone.0155979, licence creative commons CC-BY-SA 4.0
- Schollhorn R, Burris RH (1967) Acetylene as a competitive inhibitor of N-2 fixation. Proc Natl Acad Sci U S A 58: 213–216. doi: 10.1073/pnas.58.1.213