Gymnosperme
Gymnospermae

Taxons de rang inférieur
- Cycadopsida
- Ginkgoopsida
- Pinopsida
- Cupressidae
- Araucariales
- Cupressales
- Pinidae
- Gnetidae
- Cupressidae
Les Gymnospermes (Gymnospermae, du grec γυμνός / gymnós, « nu » et σπέρμα / spérma, « graine ») sont un groupe de Spermatophytes (plantes à graines) qui comprend les conifères, les cycadales, le Ginkgo et les gnetophytes. Ce sont des plantes dont l'ovule est à nu (non enclos dans un ovaire, à la différence des angiospermes) et porté par des pièces foliaires groupées sur un rameau fertile (cône). Le terme « gymnosperme » est souvent utilisé en paléobotanique pour désigner toutes les plantes à graines non angiospermes. Dans ce cas, pour spécifier le groupe monophylétique moderne des gymnospermes, le terme Acrogymnospermae est parfois utilisé.
La plupart des Gymnospermes sont des conifères (Coniferophyta, ou Pinophyta), tels que les sapins (Abies), les épicéas (Picea), les mélèzes (Larix), les séquoias géants (Sequoiadendron) , les pins (Pinus), les genévriers (Juniperus), les cyprès (Chamaecyparis et Cupressus), les ifs (Taxus), ou les thuyas (Thuja). Les cycas et les ginkgos sont également des Gymnospermes, mais appartiennent respectivement aux Cycadophyta et aux Ginkgophyta, et non aux conifères.
Description
Il existe de soixante-quinze à quatre-vingt genres et environ huit cents à mille espèces actuellement, répartis en quatorze familles. Mais il a existé jusqu'à 20 000 espèces. La flore actuelle est principalement représentée par les conifères qui comptent sept familles. On ne rencontre que des plantes ligneuses c’est-à-dire des arbres et des arbustes (on trouve quelques espèces herbacées à l'état fossile). Elles ont un cambium classique bifacial qui va donner vers l'intérieur du xylème et vers l'extérieur du phloème. Le bois est homoxylé (que des trachéides) sauf chez les Gnetales et leurs organes reproducteurs (qui ne sont pas des fleurs) sont toujours unisexués.
Reproduction
Chez les espèces dioïques, les sexes sont séparés alors que chez les espèces monoïques, les structures reproductrices mâles et femelles sont portées par un même pied. Les organes reproducteurs sont localisés au niveau des cônes. Les grains de pollen sont présents dans les cônes mâles et les cônes femelles abritent les ovules qui se transforment en graines chez les conifères et les chlamydospermes. La graine libérée contient un embryon et peut germer lorsqu'elle se pose dans un habitat propice.
Chez les Gymnospermes, la fécondation est simple, c'est-à-dire qu'un seul spermatozoïde féconde l'oosphère. Il s'agit d'une reproduction par siphonogamie, n'ayant pas besoin d'eau. Ce sont des grains de pollen (et non des spores), produits dans les cônes mâles, qui sont disséminés par le vent. Les grains de pollen peuvent voler grâce à deux petits ballonets qu'ils possèdent. Ils sont protégés du milieu extérieur par leur paroi d'exine. Le grain de pollen contient deux cellules : la cellule végétative et la cellule anthéridiale. La cellule anthéridiale se divise l'année suivante en une cellule socle et une cellule spermatogène. La cellule spermatogène se divise en deux et forme alors deux spermatozoïdes. L'ovule est composé d'un nucelle (tissu sporogène), d'un tégument (paroi) et d'une cellule mère. La cellule mère subit des mitoses et donne quatre mégaspores dont trois dégénèrent et une seule se développe en endosperme. La mégaspore restante contient un noyau qui se divise de nombreuses fois, ce qui donne finalement une cellule contenant plusieurs noyaux et une grande vacuole. On nomme cette cellule l'endosperme coenocytique. Cet endosperme évolue ensuite en prothalle femelle. Ce dernier met en place un archégone dans lequel se retrouve un oosphère. Toujours un seul oosphère par ovule peut être fécondé.
Une fois que tous les éléments précédents sont mis en place, l'année suivante il peut y avoir fécondation. Les grains de pollen arrivent au niveau de l'ovule, puis il y a formation d'un tube pollinique par la cellule végétative, capable d'atteindre l'endosperme coenocytique (le gamétophyte femelle). Ce tube pollinique déverse ensuite ses spermatozoïdes : il y en a deux de formés mais un seul peut féconder. L'un d'entre eux va jusqu'à l'oosphère, contenu dans l'archégone, où se forme le zygote. Ce zygote subit ensuite deux divisions successives qui mènent à la formation de quatre étages de quatre noyaux. À la suite de cela, une différenciation cellulaire s'effectue pour chaque étage de cellule : des cellules du suspenseur, des cellules de la rosette, des cellules ouvertes et des cellules embryonnaires. La particularité des Gymnospermes est qu'ils forment quatre embryons indépendants à partir des cellules embryonnaires, mais seul l'un d'eux se différencie, se développe et produit une graine.
La graine des gymnospermes est composée d'un embryon au milieu, entouré de réserves organiques (l'endosperme) et d'une enveloppe, nommée le tégument, provenant de l'ovule. Cette graine est nue : elle n'est pas enfermée dans un fruit. Elle peut rester en dormance et ne germer que lorsque les conditions sont favorables.
Répartition géographique
Les Gymnospermes sont surtout abondantes dans les milieux froids où elles sont très largement dominantes, grâce à leur adaptation à la sécheresse et au froid, en particulier leurs feuilles fines et aciculaires[alpha 1] (nommées « aiguilles » pour cette raison) qui limitent la surface transpirante de la plante, à cuticule épaisse, à stomates enfoncés dans des puits ou des sillons, grâce aussi à leur endurcissement hivernal (fermeture des stomates, épaississement des tissus) et la permanence du feuillage qui permet au printemps une assimilation rapide sans attendre le développement de nouvelles aiguilles, préjudiciable pour une saison végétative courte[3].
Cette physiologie permet aux plantes de devenir dominantes dans les régions bio-géographiques caractérisées par un climat froid :
- zonation en altitude : l'étagement et la composition sont liés à de nombreux facteurs comme la position de la chaîne de montagnes, le versant, les vents dominants. En France on peut distinguer 4 étages :
- collinéen (environ jusqu'à 800 m) : composé principalement de feuillus tels que les chênes,
- montagnard (de 800 à 1 600 m) : on constate l'apparition de quelques résineux tels que les Sapins et les Pins, le tout mélangé à des hêtres,
- subalpin (de 1600 à 2 400 m) : dominance absolue des conifères (Pin à crochets, Epicea, Mélèze) ; on rencontre tout de même quelques angiospermes tels que l'Aulne vert,
- alpin (à partir de 2 400 m) : toundra d'altitude où la végétation se raréfie ; on ne rencontre que quelques ligneux petits, tortueux,
- zonation en latitude : Près d'un tiers des forêts du globe sont des forêts nordiques (Taïga) composées principalement de conifères. En Europe, les forêts boréales sont dominées par les sapins, Picea et les pins, alors qu'en Amérique ce sont les Tsuga. Plus au nord, il y a disparition des formes ligneuses ; c'est la toundra, qui est dominée par les lichens et les mousses.
Il existe également d'autres types de forêts naturelles, ce sont les forêts édaphiques (liées à la nature du sol), telles que les zones marécageuses du golfe du Mexique où on rencontre le Cyprès chauve (Taxodium) ou bien les tourbières qui peuvent dans certains cas être colonisées par les Pins à crochets.
Il existe également de nombreuses forêts artificielles telles que la forêt des Landes, ou celles de la Méditerranée qui étaient autrefois peuplées par du Chêne vert.
Origine et diversité
Dès le Dévonien moyen, deux groupes de végétaux, les Lycophytes et les Progymnospermes (Gymnospermes archaïques), acquièrent le port arborescent. Les Gymnospermes sont déjà représentées dans les forêts carbonifères (Cordaitales) où la végétation est dominée par les Ptéridophytes (fougères, prêles …)[4]. Au Permo-Trias marqué par un climat plus chaud et sec, la flore à larges feuilles (Ptéridospermales ou fougères à graines, Cordaitales) fait place à une nouvelle flore cosmopolite et oligotrophe à Progymnospermes (Ginkgo, Cycadales), à nouvelles Ptéridospermales (Dicroidium (en)) et Ptéridophytes, mais aussi à Gymnospermes avec des Conifères comme Voltzia[5].
Le groupe des Gymnospermes connaît une grande extension et un maximum de diversification au Mésozoïque qui est souvent appelée « ère des Gymnospermes » [6] (simultanément, pour les vertébrés, c'est l'« ère des reptiles » car elle voit l'apogée des Dinosaures, les Gymnospermes et les fougères étant la source principale d'alimentation des dinosaures herbivores)[7], depuis les régions tropicales jusqu'aux habitats plus secs dont s'emparent les conifères et cycadales[8].
Alors que le Crétacé voit l'avènement des Angiospermes, de nombreuses Gymnospermes s'éteignent et sont supplantées dans les régions tropicales, les survivantes possédant probablement l'aptitude physiologique requise pour s'adapter à des habitats xérophytes (feuilles en écaille de Cupressacées adaptées au climat sec, résineux qui constituent les forêts sempervirentes boréale et subalpine grâce à leur adaptation à la sécheresse et au froid, en particulier leurs feuilles fines et aciculaires[alpha 1] nommées « aiguilles » pour cette raison)[8].
Au cénozoïque, les principales Gymnospermes modernes se répandent, en occupant leurs habitats actuels (Conifères adaptés au froid et aux sols pauvres, Cycadales dans les régions tropicales, Gingko sauvé de l'extinction par des moines bouddhistes chinois qui les ont élevé dans leurs temples, Gnetales à large répartition mais surtout en climat tropical)[9].
|
Gymnospermae |
Systématique
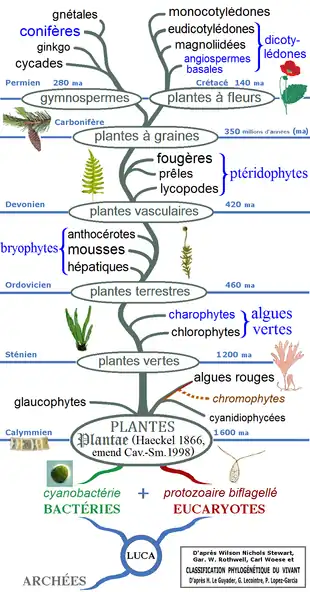
Taxonomie
C'est sans doute Théophraste (v. 374 av. J.-C. – v.286 av. J.-C.) qui, le premier, distingue les Angiospermes des Gymnospermes. John Ray utilise, à la fin du XVIIe siècle, ces différences dans sa classification qui sont les premières tentatives de classification naturelle de l'époque moderne.
Classification
| Phylogénie des Acrogymnospermae[10] - [11] - [12] - [13] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Les Gymnospermes comprennent les six ordres en quatre classes
- Sous-classe Pinidae
- Ordre des Pinales (une famille)
- Famille des Pinacées
- Ordre des Araucariales (deux familles)
- Famille des Araucariacées
- Famille des Podocarpacées
- Ordre des Cupressales (trois familles)
- Famille des Sciadopityacées
- Famille des Cupressacées
- Famille des Taxacées (inclus la famille des Céphalotaxacées)
- Ordre des Pinales (une famille)
- Sous-classe Ginkgoidae
- Ordre des Ginkgoales (une famille)
- Famille des Ginkgoacées
- Ordre des Ginkgoales (une famille)
- Sous-classe Cycadidae
- Ordre des Cycadales (trois familles)
- Famille des Cycadacées
- Famille des Stangeriacées
- Famille des Zamiacées
- Ordre des Cycadales (trois familles)
- Sous-classe Gnetidae
- Ordre des Gnétales (une famille)
- Famille des Gnetaceae
- Ordre des Ephédrales (une famille)
- Famille des Ephedraceae
- Ordre des Welwitschiales (une famille)
- Famille des Welwitschiaceae
- Ordre des Gnétales (une famille)
Phylogénie

Il s'agit d'un groupe considéré comme paraphylétique[alpha 2], regroupant Cycadophytes, Ginkgoales et Coniférophytes avec les Gnétophytes supposées apparentées aux Angiospermes sur des critères morphologiques. Mais les analyses moléculaires les plus récentes attestent plutôt la monophylie du groupe, les seules Gymnospermes plus proches des Angiospermes étant des groupes fossiles. Les Gnétales seraient le groupe frère des Pinales, voire des Pinacées.
Records
Les Gymnospermes sont les arbres de tous les records :
- de hauteur : 111,50 m pour le Sequoia sempervirens et jusqu'à 110 m pour le Douglas (Pseudotsuga menziesii) ; les séquoias géants, eux, ne font « que » 95 m. En Europe, les conifères sont plus petits avec comme arbre le plus grand un séquoia de 53 m. Le plus grand arbre connu à ce jour n'appartient pas aux Gymnospermes mais est un eucalyptus de 147 m de haut (vrai il y a 50 ans) ;
- de circonférence : 42 m pour un Taxodium mexicain et 38 m pour le Général Grant, un séquoia géant ;
- d'âge : l'arbre le plus vieux est un Pinus longaeva de 4 900 ans qui ne fait que 5 m de haut et qui se trouve à 3 000 m d'altitude. En France, il y a des ifs communs (Taxus baccata) d'environ 1 400 ans dans le Calvados.
Galerie


 Encephalartos lebomboensis - Zamiaceae
Encephalartos lebomboensis - Zamiaceae

Notes et références
Notes
- Se dit d'une feuille linéaire, rigide et pointue.
- Selon Lecointre et al., les Gymnospermes ne forment pas un groupe monophylétique sauf s'ils sont réduits aux Pinophytes[15].
Références
- Tropicos.org. Missouri Botanical Garden., consulté le 3 août 2014
- Yang Y, Ferguson DK, Liu B, Mao KS, Gao LM, Zhang SZ, Wan T, Rushforth K, Zhang ZX, « Recent advances on phylogenomics of gymnosperms and a new classification », Plant Diversity, vol. 44, no 4, , p. 340–350 (ISSN 2468-2659, DOI 10.1016/j.pld.2022.05.003, S2CID 249117306)
- Monique Schuler, Environnements et paléoclimats paléogènes, Editions du BRGM, , p. 307.
- Nicholas Polunin, Éléments de géographie botanique, Gauthier-Villars, , p. 86.
- Francis Lethiers, Évolution de la biosphère et événements géologiques, Taylor & Francis, (lire en ligne), p. 178.
- Jean-Claude Roland, Françoise Roland, François Bouteau, Hayat El Maarouf Bouteau, Atlas de biologie végétale, Dunod, , p. 116.
- (en) Karen Chin and Bruce D. Gill, « Dinosaurs, Dung Beetles, and Conifers: Participants in a Cretaceous Food Web », PALAIOS, vol. 11, no 3, , p. 280-285 (DOI 10.2307/3515235).
- (en) Stanley A. Rice, Encyclopedia of Evolution, Infobase Publishing, (lire en ligne), p. 182.
- (en) Stanley A. Rice, Encyclopedia of Evolution, Infobase Publishing, , p. 182-183.
- Andrew B. Leslie, Jeremy Beaulieu, Garth Holman, Christopher S. Campbell, Wenbin Mei, Linda R. Raubeson et Sarah Mathews, « An overview of extant conifer evolution from the perspective of the fossil record », American Journal of Botany, vol. 105, no 9, , p. 1531–1544 (PMID 30157290, DOI 10.1002/ajb2.1143, S2CID 52120430, lire en ligne)
- Andrew B. Leslie, « ajb21143-sup-0004-AppendixS4 », American Journal of Botany, (DOI 10.1002/ajb2.1143, S2CID 52120430, lire en ligne)
- Gregory W. Stull, Xiao-Jian Qu, Caroline Parins-Fukuchi, Ying-Ying Yang, Jun-Bo Yang, Zhi-Yun Yang, Yi Hu, Hong Ma, Pamela S. Soltis, Douglas E. Soltis, De-Zhu Li, Stephen A. Smith et Ting-Shuang Yi, « Gene duplications and phylogenomic conflict underlie major pulses of phenotypic evolution in gymnosperms », Nature Plants, vol. 7, , p. 1015–1025 (DOI 10.1101/2021.03.13.435279, S2CID 232282918, lire en ligne)
- Gregory W. Stull, « Gene duplications and genomic conflict underlie major pulses of phenotypic evolution in gymnosperms », Nature Plants, vol. 7, , p. 1015–1025 (DOI 10.1038/s41477-021-00964-4, lire en ligne)
- (en) Jaume Pellicer et al, « Genome Size Diversity and Its Impact on the Evolution of Land Plants », Genes, vol. 9, no 2, , p. 88 (DOI 10.3390/genes9020088).
- François Cariou, André Duco, Gérard Guillot, Claude Lebas, Guillaume Lecointre (dir.), Marie-Laure Le Louarn, Patrick Mardelle et Éric Nicol (préf. André Giordan, ill. Thomas Haessig et Dominique Visset, sous la direction de Guillaume Lecointre), Comprendre et enseigner la classification du vivant, Paris, Belin, , 2e éd. (1re éd. 2004), 351 p. [détail des éditions] (ISBN 978-2-7011-4798-7), chap. 9 (« La classification du vivant en classes préparatoires »), p. 228.
Voir aussi
Bibliographie
- (en) Charles B. Beck, Origin and evolution of Gymnosperms, Columbia University Press, , 504 p.
- (en) C.M. Govil, Gymnosperms, extinct and extant, Krishna Prakashan Media (P) Ltd, , 348 p.
Articles connexes
Références taxinomiques
- (en) Référence Flora of North America : Gymnospermae (consulté le )
- (en) Référence Trees and shrubs of the Andes of Ecuador : Gymnospermae (consulté le )
- (fr+en) Référence ITIS : Gymnospermae Non valide (consulté le )
- (en) Référence Tree of Life Web Project : Gymnospermae (consulté le )
- (en) Référence Tropicos : Gymnospermae Prantl, 1831 (+ liste sous-taxons) (consulté le )