Anoures
Les anoures, Anura, sont un ordre d'amphibiens. C'est un groupe, diversifié et principalement carnivore, d'amphibiens sans queue comportant notamment des grenouilles et des crapauds. Le plus ancien fossile de « proto-grenouille » a été daté du début du Trias à Madagascar, mais la datation moléculaire suggère que leur origine pourrait remonter au Permien, il y a 265 Ma. Les anoures sont largement distribués dans le monde, des tropiques jusqu'aux régions subarctiques, mais la plus grande concentration d'espèces se trouve dans les forêts tropicales. Il y a environ 4 800 espèces d'anoures recensées, représentant plus de 85 % des espèces d'amphibiens existantes. Il est l'un des cinq ordres de vertébrés les plus diversifiés.
Anura

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Amphibia |
| Sous-classe | Lissamphibia |
| Clade | Paratoidia |
| Super-ordre | Batrachia |
| Clade | Salientia |
Sous-ordres de rang inférieur
- Archaeobatrachia*
- clade Pipanura :
L'anoure adulte a communément un corps robuste et sans queue, des yeux exorbités, une langue bifide, des membres repliés sous le reste du corps, prêts pour des bonds à bonne distance. Vivant ordinairement en eau douce et sur le sol, certaines espèces vivent, au stade adulte, sous terre ou dans les arbres. La peau des grenouilles est glandulaire, avec des sécrétions lui donnant un mauvais goût ou la rendant toxique. Certaines espèces, appelées par convention crapauds, ont une peau dite verruqueuse dont les renflements contiennent des toxines glandulaires ; mais par la taxinomie et la phylogénie, certains crapauds sont plus proches des grenouilles que des autres crapauds. La couleur de la peau des grenouilles varie du marron tacheté, gris et vert, propices au mimétisme, à des motifs vifs de rouge ou jaune et noir signalant aux prédateurs leur toxicité.
Les œufs que ces amphibiens pondent dans l'eau donnent des larves aquatiques appelées têtards possédant des pièces buccales adaptées à un régime herbivore, omnivore ou planctonophage. Quelques espèces déposent leurs œufs sur terre ou n'ont pas de stade larvaire. Les grenouilles adultes ont généralement un régime carnivore à base de petits invertébrés, mais il existe des espèces omnivores et certaines mangent des fruits. Les grenouilles sont extrêmement efficaces pour convertir ce qu'elles mangent en biomasse formant une ressource alimentaire majeure pour des prédateurs secondaires, leur donnant ainsi le rôle de clé de voûte du réseau trophique d'un grand nombre d'écosystèmes de la planète.
Leur peau semi-perméable les rendant sensibles à la déshydratation, leur habitat est le plus souvent humide, mais certains ont connu une profonde adaptation à des habitats secs. Les grenouilles ont une riche gamme de cris et chants, en particulier en période de reproduction, ainsi qu'une large palette de comportements sophistiqués pour communiquer entre elles ou se débarrasser d'un prédateur.
Les populations de grenouilles ont considérablement diminué partout dans le monde depuis les années 1950. Plus d'un tiers des espèces sont considérées comme menacées d'extinction et plus de 120 sont soupçonnées d'avoir disparu depuis les années 1980. Le nombre de malformations chez les grenouilles est à la hausse et une maladie fongique émergente, la chytridiomycose, s'est propagée dans le monde entier. Les spécialistes de la conservation des espèces cherchent les causes de ces problèmes. Les grenouilles sont consommées par les humains et ont aussi une place notable dans la culture à travers la littérature, le symbolisme et la religion.
Étymologie
_tight_crop.jpg.webp)

Le nom anoure vient du grec οὐρά (oura), signifiant « queue », et du préfixe privatif a signifiant « sans », soit « sans queue »[1] - [2], car chez ces animaux la queue ne persiste pas après la métamorphose de passage à l'âge adulte, contrairement par exemple aux urodèles.
L'utilisation des termes « grenouilles » et « crapaud » n'a pas de justification taxonomique. Le terme « grenouille » se réfère généralement à des espèces aquatiques ou semi-aquatiques à la peau lisse et humide, tandis que l'on désigne par le terme « crapaud » des espèces terrestres à la peau sèche et verruqueuse[3] - [4]. Cependant, il y a de nombreuses exceptions à cette règle. Ainsi, le Crapaud sonneur à ventre de feu (Bombina bombina) n'a une peau que très légèrement verruqueuse et préfère les habitats aquatiques[5], tandis que la Grenouille dorée (Atelopus zeteki) est placée dans la famille des Bufonidae, qui rassemble les « vrais crapauds », et a une peau lisse[6].
Morphologie et physiologie
Aspect général et squelette
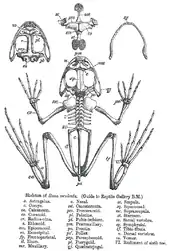
Les anoures n'ont pas de queue, excepté sous leur forme larvaire, et la plupart ont de longues pattes arrière, des os des chevilles allongés, des orteils palmés et démunis de griffes, de grands yeux et une peau lisse ou verruqueuse. Le crâne est élargi, et les arcs de la mâchoire supérieure délimitent de grandes orbites sans plancher[7]. Ils ont une colonne vertébrale courte, avec pas plus de 10 vertèbres libres et se terminant par une vertèbre sacrée unique sur laquelle s'articule un urostyle issu de la fusion des vertèbres postérieures[8] - [9]. Comme d'autres amphibiens, ils peuvent respirer à travers leur peau qui est perméable à l'oxygène. Cette caractéristique unique leur permet de rester dans l'eau, sans accès à l'air libre, en respirant par la peau[10]. Les côtes sont peu développées, généralement limitées à deux à trois paires[8], et les poumons sont remplis par pompe buccale. Il a été prouvé qu'une grenouille privée de ses poumons pouvait tout de même maintenir ses fonctions vitales[10]. Pour que la peau puisse servir d'organe de respiration, elle doit rester humide. Les anoures sont donc sensibles à diverses substances toxiques présentes dans l'environnement qui les entoure, qui peuvent se dissoudre dans le film d'eau qui recouvre leur peau et s'infiltrer dans leur sang. Ce pourrait être une des causes du déclin global de leurs populations[11] - [12] - [13] - [14].
Les anoures peuvent varier en taille entre 7,7 mm pour la récemment découverte Paedophryne amauensis de Papouasie-Nouvelle-Guinée[15] - [16] et 300 mm pour la Grenouille goliath (Conraua goliath) au Cameroun. Leur peau est lâche car démunie de tissu conjonctif. Les grenouilles ont trois paupières : une transparente qui protège les yeux sous l'eau, et deux autres qui sont translucides ou opaques. Les anoures présentent un tympan de chaque côté de la tête qui joue un rôle dans leur audition, et qui peut être couvert de peau chez certaines espèces. Les « vrais crapauds » sont totalement démunis de dents mais la plupart des grenouilles ont des dents pédicellées avec une couronne séparée de la racine par du tissu fibreux[7], placées dans les coins de la mâchoire supérieure. Elles ont également des dents vomériennes en haut de la bouche. Les anoures ne présentent en revanche jamais de dents sur la mâchoire inférieure, et avalent généralement leur nourriture entière. Les dents ont principalement comme fonction de maintenir leur proie le temps qu'ils l'avalent[17]. Les Pyxicephalus, qui se nourrissent de proies de grande taille comme les souris ou d'autres grenouilles, ont des processus osseux coniques appelés odontoïdes au niveau de la mâchoire inférieure, qui jouent le rôle de dents[18].
Pattes et membres
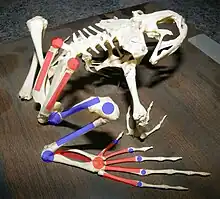
La structure des membres et des pattes varie fortement d'une espèce à l'autre, et dépend notamment de l'environnement dans lequel l'espèce évolue : sur le sol, dans l'eau, dans les arbres ou dans des terriers. Les anoures doivent être capables de se déplacer rapidement dans leur environnement pour échapper à leurs prédateurs et attraper leurs proies, et de nombreuses adaptations morphologiques les aident dans ce sens. La plupart des espèces sont bien adaptées au saut, ou du moins leurs ancêtres l'étaient, et leur morphologie y est bien adaptée. Ainsi, les différents segments du membre postérieur sont très allongés et disposés en Z, tel un ressort[19]. Le tibia, le fibula et le tarse sont fusionnés pour former un os solide, et il en est de même pour le radius et l'ulna au niveau des membres antérieurs, qui doivent absorber l'impact à la réception du saut. Le métatarse est allongé pour que les pattes postérieures soient plus longues et permettent à l'animal d'appuyer sur le sol plus longtemps. L'ilium est allongé et a une articulation mobile avec le sacrum qui, chez les sauteurs spécialisés comme les ranidés et les hylidés, participe à donner de la puissance à l'animal au moment du saut. Les vertèbres de la queue sont fusionnées pour former un urostyle qui est compris dans le pelvis. Cela permet à la force d'être transmise des pattes vers le corps durant un saut[9]. Les membres antérieurs constituent la ceinture scapulaire, rattachée à la colonne vertébrale par des muscles. Elle est également constituée de deux petits os dermiques, la clavicule et le cleithrum, ainsi que de constituants enchondraux plus importants, la scapula et le coracoïde[19].

Les adaptations au saut sont également visibles au niveau des muscles. Ainsi, les pattes postérieures des grenouilles primitives présentaient une paire de muscles antagonistes (l'un permettait de plier le genou, l'autre de l'étendre), comme chez la plupart des animaux munis de membres. Toutefois, chez les grenouilles modernes, tous les muscles participent à l'action de sauter, et seuls quelques petits muscles ont la charge de ramener la patte dans sa position de départ et maintenir cette posture. Les muscles sont également beaucoup plus gros, et les principaux muscles des pattes postérieures représentent 17 % du poids total d'une grenouille[20].
De nombreux anoures ont des pieds palmés, et le degré de palmure dépend généralement de la proportion de temps passé par la grenouille dans l'eau[21]. Les Hymenochirus, complètement aquatiques, ont des orteils totalement palmés, tandis que ceux de la Rainette de White (Litoria caerulea), une espèce arboricole, ne sont que partiellement palmés[22].

Les grenouilles arboricoles ont des disques adhésifs situés à l'extrémité de leurs orteils pour les aider à se maintenir à des surfaces verticales. Il ne s'agit pas de ventouses, mais de disques pourvus de cellules très nombreuses au sommet plat et avec de petits interstices entre chacune, lubrifiées par des glandes à mucus. Quand la grenouille fait pression avec ses pattes, les cellules adhèrent aux irrégularités de la surface et l'animal se tient au support par capillarité. Cela lui permet de grimper sur des surfaces lisses, mais cette méthode fonctionne mal si les ventouses sont trop humides[23].
Chez plusieurs grenouilles arboricoles, une « structure intercalaire » sur chaque orteil augmente la surface adhérant au substrat. Par ailleurs, comme il est dangereux de sauter dans la canopée, certaines grenouilles arboricoles ont des articulations de la hanche leur permettant à la fois de sauter et de marcher. Certaines grenouilles qui vivent en haut des arbres possèdent également une importante palmure entre les orteils. Cela permet aux grenouilles de se « parachuter » ou de planer d'un point à l'autre à travers la canopée[24].
Les grenouilles vivant sous terre ne présentent généralement pas les adaptations des grenouilles aquatiques ou arboricoles. La plupart ont de plus petites ventouses, si elles en sont pourvues, et une très légère palmure. Certains de ces animaux, comme Scaphiopus couchii, ont un tubercule kératinisé à l'extrémité des orteils des pattes postérieures, qui les aide à creuser[25].
Parfois, durant le stade larvaire, un des membres en développement du têtard est mangé par un prédateur tel qu'une larve de libellule. Dans certains cas, le membre parvient à se développer complètement, mais ce n'est pas toujours le cas, et certains anoures vivent ensuite avec seulement trois membres. Occasionnellement, un plathelminthe parasite (Ribeiroia ondatrae) s'installe dans le train arrière du têtard, ce qui provoque un réarrangement des cellules à l'origine des membres, et l'animal peut ensuite développer une ou deux pattes supplémentaires[26].
Peau

La peau des anoures, en plus de les protéger, joue un rôle dans leur respiration[27], peut absorber de l'eau et les aide à maîtriser leur température corporelle. Elle présente de nombreuses glandes, en particulier sur la tête et le dos, qui produisent généralement des substances désagréables et toxiques. La sécrétion de la peau est souvent poisseuse et contribue à garder la peau humide, protège l'animal des bactéries et le rend glissant et donc difficile à saisir pour les prédateurs[28]. La peau se régénère régulièrement, au bout de quelques semaines. L'ancienne peau se détache au milieu du dos et sous le ventre, et l'animal peut libérer ses membres de cette vieille enveloppe, qu'il consomme aussitôt[29].
Les anoures sont des animaux à sang froid et doivent adopter un comportement adéquat pour réguler leur température. Pour se réchauffer, ils se placent au soleil ou sur une surface chaude, et s'ils ont trop chaud, ils s'abritent à l'ombre ou adoptent une posture leur permettant d'exposer le moins de surface possible à l'air. Cette posture est également utilisée pour éviter de se déshydrater. L'animal est plaqué près du sol et les pattes sont rentrées sous son corps[30]. La coloration de la peau de l'anoure joue un rôle dans la thermorégulation, et l'animal peut légèrement la faire varier pour s'adapter à la température. Ainsi, dans des conditions froides et humides, la peau sera plus sombre que lors d'une journée chaude et sèche. Chiromantis xerampelina est même capable de devenir blanche pour limiter au maximum les risques de chaleur excessive[31].
Plusieurs grenouilles sont capables d'absorber de l'eau et de l'oxygène directement à travers leur peau, notamment au niveau de la région pelvique, mais la perméabilité de la peau des grenouilles accroît également les risques de déshydratation. Les glandes situées partout sur le corps sécrètent un mucus qui permet à la peau de rester constamment humide et de limiter l'évaporation. Certaines glandes sur les pattes avant et la poitrine des mâles sont spécialisées dans la production de substances poisseuses qui jouent un rôle au moment de l'amplexus. Des glandes similaires chez les grenouilles arboricoles produisent une substance collante au niveau des ventouses de leurs pattes. Certaines grenouilles arboricoles réduisent leurs pertes d'eau grâce à une couche de peau étanche, et plusieurs espèces sud-américaines ont la peau recouverte d'une sécrétion cireuse. D'autres grenouilles adaptent leur comportement pour ne pas se déshydrater : elles sont nocturnes et se reposent dans une posture limitant l'évapotranspiration. Certaines espèces se reposent en groupe comprenant un grand nombre d'individus qui se serrent les uns contre les autres. Cela limite la surface de peau exposée à l'air et donc la déshydratation[30]. Le Crapaud de Woodhouse (Bufo woodhousii), si on lui donne un accès à l'eau après qu'il a demeuré dans un endroit sec, s'assied dans l'eau peu profonde pour se réhydrater[32]. Le mâle Trichobatrachus robustus a une papille dermale proéminente à la base du dos et des cuisses, ce qui lui donne un aspect hérissé. Elle contient des vaisseaux sanguins et on pense qu'elle permet d'accroître la surface de peau disponible pour la respiration cutanée[33].
Le camouflage est une stratégie de défense commune chez les anoures. Les grenouilles les mieux camouflées sont nocturnes. Durant la journée, elles cherchent une position où elles peuvent se fondre dans le décor et y restent sans se faire repérer. Certaines grenouilles ont la capacité de changer de coloration, mais la palette de couleurs est alors restreinte. Par exemple, Litoria caerulea a une couleur qui varie entre le vert pâle et le chamoisé suivant la température, et la Rainette du Pacifique (Pseudacris regilla) peut aller du vert au brun, uni ou tacheté, suivant la période de l'année et la couleur de l'environnement proche[34]. Des caractéristiques comme des verrues ou des plis de peau sont fréquentes chez les anoures souterrains, où une peau lisse ne permet pas un camouflage efficace. Certaines grenouilles changent de couleur selon que l'on est le jour ou la nuit, la lumière et l'humidité stimulent les cellules pigmentaires et les font s'étendre ou se contracter[10].

Respiration et circulation
La peau des anoures est perméable à l'oxygène et au dioxyde de carbone, ainsi qu'à l'eau. Ils possèdent des vaisseaux sanguins près de la surface de la peau et quand une grenouille est sous l'eau, l'oxygène passe directement dans le sang. À l'air libre, les anoures respirent par pompe buccale. Leurs poumons sont similaires à ceux des humains mais les muscles de la poitrine ne jouent aucun rôle dans la respiration, et ils n'ont pas de côtes ni de diaphragme pour participer à l'entrée et la sortie de l'air. Ils gonflent leur gorge et aspirent l'air par leurs narines. Chez certaines espèces, les narines sont obturées par des valves[35]. Barbourula kalimantanensis a été découverte dans une zone isolée de l'Indonésie en 2007. Elle est entièrement aquatique et est la première espèce de grenouille connue à vivre sans poumons[36] - [37].
Les anoures ont un cœur à trois chambres, caractéristique qu'ils partagent avec les lézards[38]. Le sang oxygéné des poumons et le sang désoxygéné revenant des tissus arrivent dans deux oreillettes différentes. Quand ces chambres se contractent, les deux afflux de sang arrivent dans le ventricule commun avant qu'il ne soit pompé via une valve spiralée vers le vaisseau approprié, l'aorte pour le sang oxygéné et l'artère pulmonaire pour le sang désoxygéné. Le ventricule est partiellement divisé en cavités étroites qui minimisent le mélange des deux types de sang[39]. Cette caractéristique permet aux anoures d'avoir un métabolisme plus élevé[38].
Certaines espèces de grenouilles disposent d'adaptations qui leur permettent de survivre dans des eaux pauvres en oxygène. La grenouille du lac Titicaca (Telmatobius culeus) fait partie de ces espèces, et elle a une peau ridée qui augmente sa surface de contact pour faire des échanges de gaz avec le milieu environnant. Elle n'utilise généralement pas ses poumons rudimentaires, mais il lui arrive de se lever et se coucher successivement au fond du lac pour accroître le flux d'eau autour d'elle[40].
Digestion et excrétion
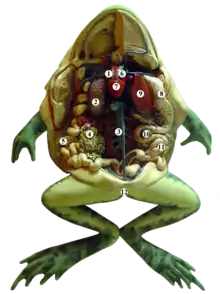
Les anoures ont des dents maxillaires sur la mâchoire supérieure qui sont utilisées pour retenir la nourriture tandis qu'ils l'avalent. Ces dents sont fragiles et ne peuvent être utilisées pour mâcher ou attraper des proies rapides. C'est pourquoi les grenouilles utilisent plutôt leur longue langue gluante pour attraper les insectes et autres petites proies rapides. La langue est normalement enroulée dans la bouche, libre à l'arrière et fixée à la mandibule à l'avant. Elle peut être lancée et rétractée à grande vitesse[21]. Certaines grenouilles n'ont pas de langue et amènent les aliments dans leur bouche avec leurs mains[21]. Les yeux aident à avaler la nourriture car ils peuvent être escamotés par des trous dans le crâne et participent à pousser la nourriture dans la gorge[21]. La nourriture traverse ensuite l'œsophage pour arriver dans l'estomac où elle est mise en contact avec les enzymes digestives. Le tube digestif se poursuit avec l'intestin grêle qui comprend le duodénum et l'iléon où se passe en grande partie la digestion. Les enzymes produites par le pancréas, et la bile produite par le foie et stockée dans la vésicule biliaire, sont sécrétées dans l'intestin grêle, où les aliments sont digérés et les nutriments absorbés. Le résidu de nourriture passe dans le gros intestin où l'excès d'eau est réabsorbé et les déchets sont évacués à travers le cloaque[41].
Bien adaptées à la vie terrestre, les grenouilles ressemblent à des poissons d'eau douce dans leur incapacité à conserver l'eau du corps efficacement. Quand ils sont sur terre, beaucoup d'eau est perdue par évaporation à travers la peau. Le système excréteur est similaire à celui des mammifères et ils disposent de deux reins qui capturent et excrètent les déchets azotés contenus dans le sang. Les grenouilles produisent de grandes quantités d'urine diluée afin d'évacuer les produits toxiques des tubules rénaux[42]. L'azote est excrété sous forme d'ammoniaque par les têtards et les grenouilles aquatiques, mais principalement sous forme d'urée, un produit moins toxique, par la plupart des adultes terrestres. Quelques espèces de grenouille arboricoles ayant moins souvent accès à l'eau excrètent de l'acide urique, encore moins toxique[42]. L'urine passe le long des uretères jumelés pour se diriger vers la vessie à partir de laquelle elle est évacuée périodiquement dans le cloaque. Tous les déchets corporels quittent le corps par le cloaque[43].
Système reproducteur
Chez les anoures mâles, les deux testicules sont attachés aux reins et la semence passe dans les reins à travers de fins tubes appelés canaux efférents. Elle passe ensuite par les uretères, qui sont des conduits urogénitaux. Il n'y a pas de pénis et le sperme est éjecté via le cloaque directement sur les œufs quand la femelle les a pondus. Les ovaires de la femelle sont derrière les reins et les œufs passent à travers une paire d'oviductes puis par le cloaque[43].
Quand la grenouille copule, le mâle monte sur le dos de la femelle et la saisit avec ses pattes avant soit au niveau des pattes avant soit au niveau des pattes arrière, voire dans le cas de Epipedobates tricolor autour du cou. Cette position est appelée amplexus et les deux partenaires peuvent la conserver pendant plusieurs jours[44]. Le mâle présente un certain nombre de caractères sexuels secondaires comme la présence de callosités rugueuses sur ses pattes en période de reproduction, lui permettant d'avoir une prise ferme[45]. L'étreinte exercée par le mâle durant l'amplexus stimule la femelle à pondre ses œufs, généralement enveloppés de gel[43]. Chez plusieurs espèces, le mâle est plus petit que la femelle. Les mâles ont des cordes vocales et peuvent émettre une grande variété de croassement, notamment pendant la période de reproduction, et certaines espèces disposent d'un sac vocal pour amplifier le son[43].
Système nerveux
Les anoures ont un système nerveux bien développé constitué d'un cerveau, d'une moelle épinière et de nerfs. Plusieurs parties du cerveau des grenouilles sont semblables à celles que l'on observe dans le cerveau humain. On rencontre en effet deux lobes olfactifs bien développés, deux hémisphères cérébraux, une glande pinéale, deux lobes optiques, un cervelet et une medulla oblongata. La coordination musculaire et les postures sont contrôlées par le cervelet, et la medulla oblongata contrôle la respiration, la digestion et d'autres fonctions automatiques[43]. Le cervelet des grenouilles est en proportion plus petit que celui de l'Homme. La moelle épinière est tubulaire et semblable à celle des autres vertébrés. Elle est courte et se prolonge dans l'urostyle uniquement par un phylum terminal[7]. Sa structure est fondamentalement tubulaire. Les anoures ont dix paires de nerfs crâniens qui transmettent les informations recueillies dans le milieu extérieur directement au cerveau, et dix paires de nerfs spinaux qui transmettent les informations venues du cerveau au reste du corps en passant par la colonne vertébrale[43] - [7]. Tous les autres amniotes (mammifères, oiseaux et reptiles) ont douze paires de nerfs crâniens[46].
Organes des sens

Les yeux de la plupart des anoures sont situés de chaque côté de la tête près de son sommet et sont bien proéminents. Ils procurent à l'animal une vision binoculaire sur un champ de 100° devant et une vision totale à presque 360°[47]. Parfois ils constituent l'unique partie submergée d'une grenouille cachée dans l'eau. Chaque œil a des paupières situées au-dessus et au-dessous et une membrane nictitante qui procure une protection supplémentaire, notamment lorsque la grenouille est en train de nager[48]. Les membres de la famille des Pipidae, qui vivent majoritairement dans l'eau, ont les yeux situés au sommet de la tête, une position plus adaptée pour repérer les proies au-dessus d'eux quand ces animaux sont partiellement immergés[47]. L'iris peut prendre diverses colorations et la pupille adopte différentes formes. Ainsi, le Crapaud commun (Bufo bufo) a des iris dorés et des pupilles fines et horizontales, la Grenouille aux yeux rouges (Agalychnis callidryas) a des pupilles verticales, Phyllobates a des iris sombres, les Crapauds sonneurs (Bombina) ont des pupilles triangulaires et le Crapaud rouge de Madagascar (Dyscophus antongilii) en a des circulaires. Les iris d'Anaxyrus terrestris sont modelés de façon à se fondre avec la peau du reste du corps[48].
La vision distante d'un anoure est meilleure que sa vision proche. Les grenouilles en train de chanter deviennent rapidement silencieuses quand elles voient un intrus, ou rien qu'une ombre, mais plus proche est l'intrus et moins bien il sera détecté[48]. Quand une grenouille lance sa langue pour attraper un insecte elle réagit au mouvement d'un petit objet qu'elle distingue mal et dont elle doit anticiper la trajectoire car elle ferme les yeux au moment où la langue est étendue[21]. La vision des couleurs par les anoures est sujet à débat, mais on a montré qu'ils réagissaient positivement à la lumière bleue, peut-être parce qu'ils l'associent aux points d'eau qui leur procurent un abri en cas de danger[49].
Les anoures peuvent entendre à l'air libre comme dans l'eau. Ils n'ont pas d'oreilles externes, leurs tympans sont directement exposés ou simplement couverts d'une couche de peau. Ils sont visibles et forment une zone circulaire juste derrière les yeux. La taille et la distance séparant les tympans dépendent de la fréquence des cris de ces animaux. Chez certaines espèces comme les Ouaouarons, la taille des tympans permet de déterminer le sexe de l'animal, car les mâles ont des tympans plus grands que leurs yeux tandis que les yeux et les tympans des femelles sont à peu près de la même taille[50]. Les bruits font vibrer le tympan et le son est transmis à l'oreille interne. L'oreille interne comprend également des canaux semi-circulaires qui jouent un rôle dans l'équilibre et l'orientation de l'animal. Les cellules ciliées qui permettent l'audition sont localisées dans deux zones de la cochlée : la papille basilaire et la papille amphibienne. La première détecte les hautes fréquences et la seconde les basses fréquences[51]. Du fait de la cochlée très courte, la grenouille utilise le réglage électrique (en) pour étendre la gamme de fréquences qu'elle peut entendre et l'aider à différencier certains sons[52]. Cet arrangement rend possible la détection des appels territoriaux de leurs congénères. Chez certaines espèces qui vivent dans les régions arides, le son de la foudre ou d'une forte pluie peut sortir les animaux de leur état de dormance[51]. Une grenouille peut être effrayée par un bruit inattendu, mais elle ne va généralement pas réagir tant que la source du bruit n'est pas repérée par la vue[50]. La morphologie particulière de leur canal auditif montre qu'il y a des composants ultrasoniques dans leurs vocalisations et que, comme certaines aloses, les anoures peuvent détecter ces ultrasons[53]. Dans le cas des grenouilles, les mâles utilisent cette capacité pour jauger leur rivaux et moduler leur comportement territorial et leur agressivité, engendrant des ajustements, triggering acoustiques complexes[54].
Pour l'odorat, les anoures disposent d'un sac nasal qui s'ouvre vers l'extérieur de chaque côté du nez par une narine externe, et s'ouvre sur la cavité buccale par un orifice en avant de la voûte buccale. Les anoures disposent également d'un organe de Jacobson, pour capter les odeurs venant de la qualité buccale[7]. Les capteurs sensoriels du goût se limitent à la face dorsale de la langue, et sont regroupés chez les anoures à l'extrémité distale de papilles fongiformes[7]. Les larves, ainsi que certains adultes aquatiques et primitifs comme les xénopes, possèdent une ligne latérale, organe sensoriel spécifique aux animaux aquatiques. Elle est constituée d'alignements céphaliques et troncaux, de neuromastes toujours superficiels, reliés à une paire de neurones bulbaires[7].
Écologie et comportement
Communication
.jpg.webp)
Le cri ou coassement des anoures est spécifique à ces espèces. Les anoures créent ce son en faisant passer l'air à travers leur larynx. Chez la plupart des grenouilles qui coassent, ce cri est amplifié par un ou plusieurs sacs vocaux, des membranes de peau placées sous la gorge ou dans le coin de la bouche, qui se détendent au cours du cri. Certains coassements sont si bruyants qu'ils peuvent être entendus à plus d'un kilomètre[55].
Les grenouilles des genres Heleioporus et Neobatrachus n'ont pas de sac vocal mais sont tout de même capables d'émettre un cri puissant. Leur bouche est élargie et à la forme d'un dôme, et agit comme une caisse de résonance qui amplifie le son. Les espèces de grenouilles qui n'ont pas de coassement bruyant sont généralement celles qui vivent dans les zones proches d'eau coulant en provoquant un bruit continu. Elles ont besoin d'utiliser d'autres stratégies pour communiquer. La Grenouille-à-queue côtière (Ascaphus truei) vit dans les ruisseaux de montagne en Amérique du Nord et n'émet pas de chant[56].
Le but du coassement est d'attirer une partenaire pour les mâles. Ils peuvent chanter seuls ou en groupe lorsque de nombreux mâles ont convergé vers un même site de reproduction. Les femelles de plusieurs espèces de grenouilles, comme Polypedates leucomystax, répondent aux cris du mâle, ce qui renforce l'activité reproductive dans une colonie[57]. Les femelles grenouilles préfèrent les mâles qui produisent des sons de grande intensité et de basse fréquence, des caractéristiques qui les font sortir du lot dans un chœur[58].
Un cri différent est émis par une grenouille mâle ou une femelle non réceptive lorsqu'un autre mâle tente de la chevaucher. Il s'agit d'un gazouillis caractéristique qui est accompagné d'une vibration du corps[59]. Les grenouilles arboricoles et certaines espèces non-aquatiques ont un chant particulier qu'elles entonnent avant une pluie, qu'elles anticipent sur la base de l'humidité de l'air ambiant[59]. Certaines espèces ont également un chant territorial qui permet de chasser les autres mâles. Les anoures émettent ces chants avec la bouche fermée[59]. Un appel de détresse, émis par certaines grenouilles quand elles sont en danger, est produit avec la gueule ouverte, ce qui permet un cri un peu plus fort. Il est généralement utilisé lorsque la grenouille est attrapée par un prédateur, et peut servir pour faire diversion et déconcerter le prédateur afin qu'il relâche sa proie[59].
Plusieurs espèces de grenouilles ont un cri profond. Le coassement du ouaouaron (Lithobates catesbeianus) est parfois décrit comme un « jug o' rum »[60]. La Rainette du Pacifique (Pseudacris regilla) émet un onomatopéique « ribbit » que l'on écoute souvent dans les films[61]. D'autres chants sont décrits comme un « brekekekex koax koax », le cri de la Grenouille rieuse (Pelophylax ridibundus) dans Les Grenouilles, un ancien drame comique grec d'Aristophane[62].
Torpeur
Lors de conditions extrêmes, certaines grenouilles entrent dans un état de torpeur, et restent inactives pendant des mois. Dans les régions les plus froides, plusieurs espèces de grenouilles hibernent en hiver. Celles qui vivent sur terre comme le Crapaud d'Amérique (Bufo americanus) creusent un terrier et bâtissent un hibernaculum où elles peuvent entrer en dormance. D'autres, moins efficaces pour creuser, trouvent une crevasse ou s'enterrent dans les feuilles mortes. Des espèces aquatiques comme la Grenouille taureau (Rana catesbeiana) coulent au fond de la mare ou de l'étang dans lequel ils vivent, moitié ensevelis dans la vase mais toujours capables d'absorber l'oxygène dissous dans l'eau. Leur métabolisme se ralentit et ils vivent sur leurs réserves énergétiques. Certaines grenouilles peuvent même survivre à la congélation. Des cristaux de glace se forment sous leur peau et dans la cavité de leur corps mais les organes essentiels sont protégés des dégâts de la congélation par leur forte concentration en glucose. Une grenouille apparemment sans vie, congelée, peut reprendre sa respiration et voir son cœur repartir quand la température remonte[63].
À l'autre extrême, Cyclorana alboguttata entre en estivation régulièrement durant la saison chaude et sèche en Australie, survivant dans un état de dormance sans accès à la nourriture et à l'eau pendant neuf à dix mois de l'année. Elle s'enterre sous le sol et s'enferme dans un cocon protecteur formé par sa mue. Les chercheurs de l'Université du Queensland ont établi que durant l'estivation, le métabolisme de la grenouille est modifié et l'efficacité opérationnelle des mitochondries est améliorée. Cela signifie que les quantités limitées d'énergie disponible pour la grenouille en torpeur sont utilisées de manière plus efficace. Ce mécanisme de survie est seulement utile aux animaux qui restent complètement inconscients pendant une très longue période de temps et dont les besoins en énergie sont faibles car ils sont à sang froid et n'ont pas besoin de générer de chaleur[64]. D'autres recherches ont montré que, pour pourvoir à ces besoins énergétiques, les muscles s'atrophiaient, mais que les muscles des pattes postérieures étaient souvent épargnés par ce phénomène[65].
Locomotion
Les différentes espèces de grenouilles utilisent diverses méthodes de locomotion comme le saut, la course, la marche, la nage, et peuvent aussi creuser des tunnels, grimper et planer.
Le saut

Les grenouilles sont généralement reconnues comme étant bien adaptées au saut, et sont dans ce sens les vertébrés qui sautent le plus loin par rapport à leur taille[66]. Litoria nasuta peut sauter à plus de 2 m, une distance qui représente plus de 50 fois la longueur de son corps, qui est de 5,5 cm[67]. Il y a de grandes différences d'une espèce à l'autre quant à leurs aptitudes au saut. Au sein d'une même espèce, plus l'animal est grand et plus il saute loin, mais le rapport entre la distance sautée et la longueur diminue. Euphlyctis cyanophlyctis a la capacité de sauter en dehors de l'eau à partir d'une position flottante à la surface[68]. La toute petite Rainette grillon (Acris crepitans) peut traverser une mare avec une série de courts sauts rapides sur sa surface[69].
Le visionnage de ce mouvement au ralenti montre que les muscles ont une certaine flexibilité. Ils sont d'abord étendus quand la grenouille est en position accroupie, puis ils sont contractés avant d'être étendus à nouveau pour permettre à la grenouille de se lancer dans l'air. Les pattes antérieures sont repliées contre la poitrine et les pattes postérieures restent dans la position étendue pendant le saut[20]. Chez certaines espèces particulièrement bien adaptées au saut, comme la Rainette de Cuba (Osteopilus septentrionalis) et la Grenouille léopard (Rana pipiens), la puissance produite durant un saut peut dépasser celle que le muscle peut théoriquement produire. Lorsque le muscle se contracte, l'énergie est tout d'abord transférée vers le tendon étiré qui entoure l'os de la cheville. Ensuite, les muscles s'étirent à nouveau tandis que dans le même temps le tendon libère son énergie telle une catapulte afin de produire une puissance d'accélération dépassant les limites du muscle[70]. Un mécanisme similaire est connu chez les locustes et les sauterelles[71].
Marche et course
Les anoures de la famille des Bufonidae, des Rhinophrynidae et des Microhylidae ont de courtes pattes postérieures et sont plus enclins à marcher qu'à sauter[72]. Quand ils essaient de se déplacer plus rapidement, ils accélèrent la vitesse des mouvements de leurs membres ou adoptent une démarche sautillante. La démarche de Gastrophryne olivacea a été décrite comme une « combinaison entre de la course et de petits bonds de deux à cinq centimètres de long »[73]. Dans une expérimentation, un Bufo fowleri était placé sur un ergomètre tournant à différentes vitesses. En mesurant la quantité d'oxygène consommée par le crapaud on s'est rendu compte que ce sautillement utilisait de manière inefficace l'énergie lors de longs trajets, mais était une stratégie payante lors de courtes périodes d'activité intense[74].
Kassina maculata a de courtes et frêles pattes postérieures mal adaptées au saut. Elle peut se déplacer rapidement en « courant » en utilisant ses deux pattes alternativement. Au ralenti, on s'en rend compte qu'à la différence d'un cheval qui peut trotter ou galoper, la démarche de cette grenouille est strictement identique quelle que soit sa vitesse d'avancement[75]. Cette espèce peut également grimper dans des arbres et des arbustes, ce qu'elle fait souvent la nuit à la recherche d'insectes[76]. Euphlyctis cyanophlyctis a de larges pattes et peut courir à la surface de l'eau sur quelques mètres[69].
Nage

Les anoures qui vivent ou vont parfois dans l'eau disposent d'adaptations pour améliorer leur faculté à nager. Les pattes postérieures ont notamment une forte musculature, et les orteils sont intégralement palmés, ce qui augmente la surface des pattes et aide l'animal à se propulser dans l'eau. Les membres de la famille des Pipidae sont totalement aquatiques et sont les plus spécialisés dans ce domaine. Ils ont une colonne vertébrale très peu flexible, un corps aplati, un système de ligne latérale et de puissantes pattes postérieures aux orteils palmés[77]. Les têtards ont une grande nageoire caudale qui leur permet d'avancer en remuant la queue d'un côté à l'autre[78].
Vie souterraine
Certains anoures se sont adaptés à la vie dans des galeries souterraines et peuvent creuser. Ils ont généralement un corps arrondi, de courts membres, une petite tête aux yeux renflés et des pattes de derrière adaptées pour creuser. Un exemple extrême est Nasikabatrachus sahyadrensis qui vit dans le sud de l'Inde et se nourrit de termites, et qui passe presque l'intégralité de sa vie sous la surface du sol. Elle en émerge brièvement durant la mousson pour se reproduire dans des mares temporaires. Elle a une minuscule tête avec un museau pointu et un corps globuleux. Du fait de son existence souterraine, elle a été décrite scientifiquement pour la première fois en 2003, et n'était connue auparavant que par la population locale[79].

Les Scaphiopodidae d'Amérique du Nord sont également bien adaptés à la vie souterraine. Le Crapaud des Plaines (Spea bombifrons) en est un exemple typique avec un volet d'os kératinisé relié à l'un des métatarses qui est utilisé pour creuser en reculant. Quand il creuse, le crapaud tortille ses hanches d'un côté à l'autre pour s'enfoncer dans le sol. Il dispose ainsi d'un terrier peu profond en été duquel il émerge la nuit pour aller se nourrir. En hiver il creuse plus profondément et on l'a retrouvé jusqu'à 4,5 m de profond[80]. Le tunnel est rempli de terre et le crapaud hiverne dans une petite chambre située à son extrémité. Pendant ce temps, l'urée s'accumule dans les tissus et l'eau du sol est absorbée par le crapaud par osmose pour subvenir à ses besoins[80]. Les Scaphiopodidae se reproduisent en masse. Ils émergent tous de leurs terriers au même moment et se dirigent vers des mares temporaires, attirés par le chant du premier mâle à trouver une telle opportunité pour se reproduire[81].
Les grenouilles australiennes vivant sous le sol ont un mode de vie semblable. Heleioporus albopunctatus creuse un terrier à côté d'une rivière ou dans le lit d'un ruisseau temporairement à sec, et en émerge régulièrement pour se nourrir. L'accouplement a lieu et les œufs sont pondus dans nid d'écume dans le terrier. Les œufs se développent partiellement ici mais n'éclosent pas tant qu'ils ne sont pas immergés par de fortes précipitations. Les têtards se dirigent alors vers l'étendue d'eau la plus proche et terminent rapidement leur développement[82]. Certaines espèces de Madagascar s'enterrent généralement simplement dans la litière de feuille. Parmi elles Scaphiophryne marmorata a une tête aplatie, un museau court et des tubercules bien développés au niveau du métatarse, en faisant un animal bien adapté pour creuser. Par ailleurs ses orteils des pattes antérieures se terminent par des disques qui lui permettent de grimper dans les buissons[83]. Elle se reproduit dans des points d'eau temporaires après la pluie[84].
Mœurs arboricoles

Les grenouilles arboricoles vivent haut dans la canopée, où elles naviguent entre les branches, les brindilles et les feuilles, parfois ne descendant jamais à terre. Les grenouilles de la famille des Hylidae sont un exemple typique de grenouilles arboricoles, mais certaines autres espèces ont développé un mode de vie similaire, comme les Centrolenidae, les Hyperoliidae, certaines Microhylidae et les Rhacophoridae[72]. La plupart des grenouilles arboricoles mesurent moins de 10 cm de long, avec de longues pattes arrière et de longs orteils équipés de ventouses à leur extrémité. La surface de ces ventouses est constituée d'une couche de cellules épidermiques à sommet plat hexagonales, très rapprochées et seulement séparées par des sillons au sein desquels des glandes sécrètent du mucus. Ces ventouses, maintenues humides par le mucus, permettent à ces animaux de s'agripper à des surfaces sèches comme mouillées, y compris du verre. Elles utilisent la force de tension superficielle et la viscosité[85]. Certaines grenouilles sont très acrobatiques et peuvent attraper des insectes tandis qu'elles se tiennent à une brindille par un orteil ou qu'elles serrent la tige d'un roseau balayé par le vent[86]. Certains membres de la sous-famille des Phyllomedusinae ont des orteils en position opposés. La Phyllomedusa ayeaye a un doigt opposé sur chacune de ces pattes avant et deux doigts opposés sur ses pattes arrière. Cela lui permet de saisir les tiges des plantes quand elle se déplace en grimpant dans la végétation en bord de rivière[87].
Déplacement en planant
Durant l'histoire évolutive de la grenouille, différents groupes ont développé un déplacement par la voie des airs[88]. Certaines grenouilles dans la forêt tropicale sont bien adaptées pour planer d'arbres en arbres ou pour retomber sur le sol de la forêt. La Grenouille volante de Wallace (Rhacophorus nigropalmatus) en est un exemple typique que l'on trouve en Malaisie et à Bornéo. Elle a de larges pattes avec l'extrémité des doigts formant de larges disques adhésifs et une palmure complète. Les membres sont bordés de bandes de peau libre. En écartant les doigts, étendant les membres et déployant ses bandes de peau, la grenouille peut planer sur des distances considérables[89]. Elle peut changer sa direction pendant qu'elle plane et naviguer ainsi entre les arbres sur des distances pouvant atteindre 15 m[90].
Reproduction
Principalement deux types de reproduction sont utilisés par les anoures : la reproduction prolongée et la reproduction explosive. Dans le premier cas, les adultes se rassemblent certaines périodes de l'année dans les étangs, lacs ou ruisseaux pour se reproduire. La plupart des animaux retournent vers le lieu où ils sont nés et ont grandi. Il en résulte d'importantes migrations de centaines d'individus. Dans l'autre cas, les grenouilles adultes arrivent aux points de reproduction en réponse à certains facteurs, comme des averses dans les régions extrêmement arides. Chez les espèces concernées, la copulation et la ponte ont lieu très rapidement après l'arrivée des animaux sur le site de reproduction, et les têtards se développent très rapidement afin d'utiliser les points d'eau temporaires avant qu'ils ne s'assèchent à nouveau[91].
Chez les espèces qui se rassemblent saisonnièrement, les mâles arrivent généralement les premiers sur les sites de reproduction, et y restent quelque temps avant l'arrivée des femelles, ces dernières quittant le site rapidement après la ponte. Cela implique que les mâles sont plus nombreux que les femelles à un instant donné, et ils défendent un territoire duquel ils excluent les autres mâles. Ils indiquent leur présence en coassant, alternant souvent leurs appels avec les coassements des animaux voisins. Les grenouilles les plus grosses et les plus fortes émettent les coassements les plus profonds, et elles peuvent garder les meilleurs territoires. Les femelles choisissent leur partenaire en partie sur la profondeur de leur chant[92]. Chez certaines espèces certains mâles n'ont pas de territoire et ne chantent pas. Ils interceptent des femelles qui se dirigent vers des mâles coassant ou prennent un territoire lorsqu'il devient vacant. Le chant est une activité qui demande beaucoup d'énergie. Parfois il y a des échanges de rôle, et un mâle disposant d'un territoire le laisse pour graviter librement autour de celui des autres[91].
.jpg.webp)
Chez les espèces adeptes d'une reproduction explosive, le premier mâle qui trouve un lieu approprié pour se reproduire appelle d'un coassement bruyant, attirant ses congénères des deux sexes. Les animaux chantent ensuite à l'unisson, formant un chœur que l'on peut entendre de très loin. Les Scaphiopus d'Amérique du Nord entrent dans cette catégorie. La sélection du partenaire et la cour n'ont que peu d'importance par rapport à la rapidité avec laquelle les animaux s'accouplent. Certaines années, les conditions favorables ne se produisent pas et les animaux peuvent rester deux années ou plus sans se reproduire[91]. Certaines femelles Spea multiplicata pondent uniquement la moitié de leurs œufs à la fois, peut-être pour en conserver si une meilleure opportunité pour se reproduire se produisait ultérieurement[93].
Sur le site de reproduction, le mâle monte sur la femelle et s'accroche solidement sur son corps. Typiquement, l'amplexus a lieu dans l'eau, la femelle relâchant ses œufs et le mâle les couvrant avec son sperme : la fécondation est donc externe. Chez certaines espèces comme le Crapaud des steppes (Bufo cognatus), le mâle retient les œufs avec ses pattes arrière, les gardant en place pendant environ trois minutes[91]. Les membres du genre Nimbaphrynoides que l'on trouve en Afrique de l'Ouest sont uniques parmi les anoures du fait de leur viviparité. Les membres du genre tanzanien Nectophrynoides sont les seuls anoures ovovivipares. Dans les deux cas, la fertilisation est interne et les femelles donnent naissance à de jeunes grenouilles complètement développées[94] - [95].
Cycle de vie



Comme d'autres amphibiens, le cycle de vie de la grenouille commence dans l'eau avec un œuf qui éclos pour donner une larve sans membres munie de branchies, que l'on appelle têtard. Après une période de croissance, durant laquelle le têtard développe des membres et des poumons, il se métamorphose et son aspect général et l'arrangement de ses organes internes sont modifiés. Il peut quitter l'eau sous la forme d'une grenouille miniature.
Œufs
Les embryons des anoures sont généralement entourés de plusieurs couches d'une substance gélatineuse. La gélatine protège l'œuf tout en permettant le passage de l'oxygène, du dioxyde de carbone et de l'ammoniac. Elle absorbe l'humidité et gonfle en contact avec l'eau. Après la fertilisation, la portion la plus interne se liquéfie pour permettre les libres mouvements de l'embryon en développement. Chez certaines espèces, comme la Grenouille à pattes rouges (Rana aurora) et la Grenouille des bois (Rana sylvatica), une algue verte unicellulaire symbiotique est présente dans la gélatine. On pense qu'elle est utile à l'embryon en développement en lui pourvoyant de l'oxygène supplémentaire à travers la photosynthèse[96]. Les œufs sont noirs ou marron foncé, et ont l'avantage d'absorber la chaleur du soleil qui est retenue par la capsule. L'intérieur des amas d'œufs de la Grenouille des bois (Rana sylvatica) est 6 °C plus chaud que l'eau environnante et cela accélère le développement de la larve[97].
La forme et la taille des œufs sont caractéristiques de l'espèce concernée. Les ranidés produisent des amas d'œufs globulaires comprenant un grand nombre d'œufs, tandis que les bufonidés produisent des amas d'œufs formant de longues chaînes cylindriques. La minuscule Eleutherodactylus limbatus pond des œufs isolés, les enterrant dans le sol humide[98]. Leptodactylus pentadactylus fait un nid de mousse dans un arbre creux. Les œufs éclosent lorsque le nid est inondé, ou les têtards terminent leur développement dans la mousse s'il n'y a jamais d'arrivée d'eau[99]. La Grenouille aux yeux rouges (Agalychnis callidryas) dépose ses œufs sur une feuille au-dessus d'un point d'eau et quand elles éclosent les larves tombent dans l'eau en dessous[100]. Les larves en développement dans les œufs peuvent détecter les vibrations causées par des guêpes ou serpents prédateurs, et peuvent éclore plus précocement pour éviter d'être mangés[101]. En général, la longueur d'incubation dépend de l'espèce et des conditions environnementales. Les œufs aquatiques éclosent généralement au bout d'une semaine, quand la capsule se fend sous l'action d'une enzyme relâchée par la larve en développement[102].
Têtards
.jpg.webp)
Les larves qui émergent des œufs, que l'on connaît sous le nom de têtards, ont généralement un corps ovale allongé, avec une queue aplatie verticalement. En règle générale, les têtards sont totalement aquatiques, mais au moins une espèce, Nannophrys ceylonensis, a des têtards semi-aquatiques qui peuvent vivre sur des rochers humides[103] - [104]. Les têtards n'ont pas de paupière et ont des squelettes cartilagineux, une ligne latérale, des branchies pour assurer leur respiration (branchies externes dans un premier temps puis branchies internes) et une queue aplatie verticalement qu'ils utilisent pour nager[78].
Au début de son développement, le têtard possède une poche branchiale qui recouvre ses branchies et ses pattes avant. Les poumons commencent rapidement à se développer et sont utilisés de manière complémentaire pour la respiration. Certaines espèces réalisent leur métamorphose dans l'œuf et éclosent directement sous la forme de grenouilles miniatures. Les têtards sont dépourvus de vraies dents, mais les mâchoires supérieures de certaines espèces présentent deux rangées parallèles de structures kératinisées. Les mâchoires inférieures portent généralement trois rangées de ces dents entourées d'un bec corné, mais le nombre de rangées peut varier et l'agencement exact de la bouche constitue un moyen d'identification des espèces[102]. Chez les Pipidae, à l'exception de Hymenochirus, les têtards ont une pare de barbillons antérieurs, qui les font ressembler à des poissons-chats[77]. Leurs queues sont raidies par une chorde, mais ne contiennent aucun élément osseux ou cartilagineux, excepté quelques vertèbres à la base qui forme l'urostyle durant la métamorphose. On pense qu'il s'agit d'une adaptation à leur mode de vie. Comme leur transformation en grenouilles arrive très rapidement, la queue est uniquement constituée de tissu mou, car l'os et le cartilage prennent plus de temps à être absorbés. La nageoire caudale et l'extrémité de la queue sont fragiles et se déchirent rapidement, ce qui est vu comme une adaptation pour échapper à des prédateurs qui tentent de les attraper par la queue[105].
Les têtards sont principalement herbivores, se nourrissant majoritairement d'algues comme les diatomées qu'il filtre dans l'eau à travers ses branchies. Certaines espèces sont carnivores au stade têtard, mangeant des insectes, de plus petits têtards et des poissons. La Rainette de Cuba (Osteopilus septentrionalis) fait partie des espèces chez lesquelles les têtards peuvent être cannibales. Les têtards qui développent leurs membres le plus rapidement peuvent être mangés par les autres, et ce sont donc les animaux qui ont le développement le plus long qui sont avantagés[106].
Les têtards sont très vulnérables aux poissons, aux tritons, aux dytiques et aux oiseaux tels que les martins-pêcheurs qui s'en nourrissent. Certains têtards, comme ceux du Crapaud buffle (Rhinella marina), sont poisons. Le stade têtard peut durer seulement une semaine chez des espèces à la reproduction rapide ou peut laisser l'animal passer un ou deux hivers avant qu'il ne se métamorphose au printemps[107].
Métamorphose
À la fin du stade têtard, la grenouille se métamorphose et son corps se transforme rapidement en celui d'un adulte. Cette métamorphose dure seulement 24 heures, et est initiée par la production d'une hormone, la thyroxine. Cela conduit différents tissus à se développer de différentes façons. Les principaux changements sont le développement de poumons et la disparition des branchies et des poches branchiales, laissant les pattes avant visibles. La mâchoire inférieure devient la grande mandibule de l'adulte carnivore, et le long intestin spiralé du têtard herbivore est remplacé par un intestin court typique d'un prédateur[102]. Le système nerveux devient bien adapté à l'audition et la vision stéréoscopique, et pour de nouvelles méthodes de locomotion et d'alimentation[102]. Les yeux sont repositionnés plus haut sur la tête et les paupières et glandes associées apparaissent. Le tympan, l'oreille moyenne et l'oreille interne se développent. La peau devient plus épaisse et plus dure, la ligne latérale disparaît et les glandes de la peau apparaissent[102]. La dernière étape est la disparition de la queue, mais elle a lieu plus tard, le tissu étant utilisé pour accélérer la croissance des membres[108]. C'est lors de leur métamorphose que les anoures sont les plus sensibles aux prédateurs. En effet, les grenouilles ont tout juste perdu leur queue et leurs membres commencent tout juste à être utilisés pour la locomotion[72].
 Larve de Grenouille rousse Rana temporaria quelques jours avant la métamorphose.
Larve de Grenouille rousse Rana temporaria quelques jours avant la métamorphose. Étape de la métamorphose avec la mâchoire déformée, de grands yeux et une relique de la poche branchiale.
Étape de la métamorphose avec la mâchoire déformée, de grands yeux et une relique de la poche branchiale..JPG.webp) Jeune grenouille avec courte queue, dont la métamorphose est presque complète.
Jeune grenouille avec courte queue, dont la métamorphose est presque complète.
Adultes
Après la métamorphose, les jeunes adultes se dispersent dans leur nouvel habitat terrestre ou continuent à vivre dans l'eau. Presque tous les anoures sont carnivores au stade adulte, se nourrissant d'invertébrés, comme des arthropodes, des annélides, des gastéropodes, et des limaces[109]. Quelques-unes des plus grosses espèces peuvent manger d'autres grenouilles, des petits mammifères et des poissons. Certaines grenouilles utilisent leur langue gluante pour attraper des proies rapides[110], tandis que d'autres poussent la nourriture dans leur bouche avec leurs pattes. La grenouille arboricole Xenohyla truncata est une exception car en partie herbivore, son alimentation comprenant une grande proportion de fruit[111]. Les grenouilles adultes sont elles-mêmes les proies de différents prédateurs. La Grenouille léopard (Rana pipiens) est par exemple consommée par les hérons, les faucons, les poissons, les grandes salamandres, les serpents, les ratons laveurs, les moufettes, les visons, les grenouilles-taureaux et d'autres animaux[112].
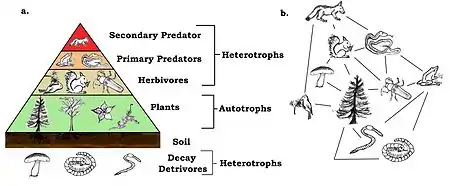
Les anoures sont des prédateurs primaires et un maillon important de la chaîne alimentaire. Étant hétérotherme, ils font une utilisation efficace de leur alimentation en dépensant peu d'énergie pour les processus métaboliques, tandis que le reste est transformé en biomasse. Ils sont eux-mêmes consommés par les prédateurs secondaires et sont les principaux consommateurs terrestres d'invertébrés, la plupart attrapés sur des plantes. En réduisant la population d'herbivore, ils jouent un rôle dans la croissance des plantes et participent à l'équilibre des écosystèmes[113].
On sait peu de choses sur la longévité des anoures à l'état sauvage. La squelettochronologie est une méthode permettant d'évaluer l'âge des animaux à partir du squelette. Cette méthode a été utilisée pour étudier la Grenouille des montagnes à pattes jaunes (Rana muscosa), dont les phalanges des orteils présentaient des lignes de croissance saisonnière avec ralentissement de la croissance en hiver. La plus vieille grenouille avait dix lignes, et on estimait donc son âge à 14 ans, en comptant les quatre ans de stade têtard[114]. Certains animaux détenus en captivité ont vécu jusqu'à 40 ans, record obtenu pour un Crapaud commun (Bufo bufo). Le Crapaud buffle (Bufo marinus) peut vivre 24 ans en captivité, et la Grenouille-taureau (Rana catesbeiana) 14 ans[115]. Les grenouilles des climats tempérés hibernent durant l'hiver, et quatre espèces sont connues pour pouvoir supporter de geler durant cette période, dont la Grenouille des bois (Rana sylvatica)[116].
Soins parentaux


Bien que les soins apportés à la progéniture soient mal étudiés chez les grenouilles, près de 20 % pourrait s'occuper d'une manière ou d'une autre de l'avenir de leur progéniture[117]. L'évolution des soins parentaux chez les grenouilles est dirigée principalement par la taille de l'étendue d'eau dans lequel elles se reproduisent. Les espèces qui se reproduisent dans de petits points d'eau ont généralement des comportements parentaux plus complexes[118]. Du fait de la prédation importante des œufs et des larves dans les points d'eau de plus grande taille, un grand nombre d'espèces pondent leurs œufs sur la terre. Un des deux parents doit alors s'assurer que les œufs ne se dessèchent pas en maintenant une certaine humidité pour assurer leur survie[119]. Par ailleurs les parents doivent ensuite assurer le transport des têtards juste éclos vers un point d'eau[118].
Dans les petits points d'eau, les prédateurs sont la plupart du temps absents et la compétition entre les têtards est la principale limite à leur survie. Certaines espèces de grenouilles évitent cette concurrence en utilisant de petits phytotelmes (axiles de feuilles remplis d'eau ou cavité dans du bois) comme sites pour déposer quelques têtards[120]. Si ces sites permettent aux têtards de ne pas être en compétition, ils manquent également de nutriments pour survivre sans aide de leurs parents. Les espèces ayant adopté cette stratégie laissent à leurs têtards des œufs remplis de substances nutritives mais non fécondés[118]. La femelle Grenouille des fraises (Oophaga pumilio) pond ses œufs sur le sol de la forêt tropical. Le mâle les protège des prédateurs et leur apporte de l'eau via son cloaque pour les garder humides. Quand ils éclosent, la femelle emmène les têtards sur son dos pour rejoindre une broméliacée remplie d'eau ou autre point d'eau similaire, en déposant seulement un œuf à chaque endroit. Elle les visite régulièrement et les nourrit en pondant un ou deux œufs non fertilisés dans le phytotelma, et ce jusqu'à ce que le têtard atteigne le stade de la métamorphose[121]. Oophaga granulifera s'occupe de ses têtards d'une manière similaire[122].
Diverses autres formes de soins parentaux peuvent être observées chez les grenouilles. Le petit mâle Colostethus subpunctatus garde les œufs pondus par sa femelle, sous une pierre ou un morceau de bois. Quand les œufs éclosent, il transporte les têtards sur son dos jusqu'à une mare temporaire, où il s'immerge en partie pour laisser tomber un ou quelques têtards, avant de se diriger vers un autre point d'eau et recommencer l'opération[123]. Le mâle Alyte accoucheur (Alytes obstetricans) porte les œufs avec lui, attachés sur ses pattes arrière. Il les conserve humide en s'immergeant régulièrement dans l'eau et leur évite de devenir trop humide en élevant ses quartiers arrière. Après trois à six semaines, il se dirige vers un point d'eau et les œufs éclosent[124]. Engystomops pustulosus construit un nid flottant fait de mousse pour protéger ses œufs de la prédation. Cette mousse est faite de protéines et de lectines et semble avoir des propriétés antimicrobiennes[125]. Plusieurs couples de grenouilles peuvent construire un nid de ce type en commun, à partir d'un radeau préexistant. Les œufs sont pondus au centre, suivis de couches alternatives de mousse et d'œufs, le tout recouvert de mousse[126].
Certaines grenouilles protègent leur descendance dans leur propre corps. Les mâles et les femelles Assa darlingtoni gardent leurs œufs, qui sont pondus sur le sol. Une fois éclos, le mâle lubrifie son corps avec le gel les entourant, et s'immerge dans la masse d'œufs éclos. Les têtards se dirigent alors vers des poches de peau qu'il possède sur ses flancs, où ils vont se développer jusqu'à la métamorphose[127]. Les femelles du genre Rheobatrachus, présent en Australie et probablement éteinte aujourd'hui, avalent les œufs fertilisés, qui se développent dans leur estomac. Elles cessent alors de se nourrir et de sécréter des sucs gastriques. Les têtards utilisent le vitellus des œufs pour se nourrir. Après 6 ou 7 semaines, ils sont prêts à se métamorphoser. La mère régurgite les petites grenouilles, qui s'échappent de sa bouche[128]. La femelle Rhinoderma darwinii du Chili pond jusqu'à 40 œufs sur le sol, où ils sont gardés par le mâle. Quand les têtards sont prêts à éclore, ils sont gobés par le mâle, qui les garde dans son sac vocal particulièrement grand. Ils sont immergés dans un liquide visqueux et mousseux qui contient des matières nutritives supplémentaires à celle du vitellus. Ils y restent 7 à 10 semaines avant de se métamorphoser, après quoi ils sortent de la bouche du mâle[129].
Défense


Les anoures ont de nombreux prédateurs potentiels (dont d'autres amphibiens tels que les salamandres) [130]. À première vue, les anoures semblent sans défense avec leur petite taille, leur peau fine et l'absence de défenses telles que des épines, des griffes ou des dents. Plusieurs utilisent le camouflage pour éviter d'être vue par certains prédateurs (Une peau tachetée ou rayée de couleurs identiques à celles de son environnement permettent à l'animal d'être difficile à percevoir, il existe des anoures qui ont la couleur, mais aussi la forme d'une feuille morte). Certaines espèces peuvent faire de grands bonds, généralement vers l'eau, pour échapper à leurs prédateurs, tandis que d'autres ont développé d'autres méthodes de défense[91].
La peau de plusieurs anoures contient des substances toxiques appelées bufotoxines qui les rendent immangeables pour les prédateurs. La plupart des crapauds et certaines grenouilles ont de grandes glandes productrices de poison, les glandes parotoïdes, situées sur les côtés de leurs têtes derrière les yeux, ainsi que d'autres glandes un peu partout sur le corps. Des glandes sécrètent aussi un mucus et des substances rendant les animaux glissants et donc difficiles à saisir. Si les effets nocifs du poison ou de molécules répulsives sont immédiats, le prédateur stoppe son agression et l'anoure peut s'échapper. Si les effets sont plus tardifs, le prédateur va apprendre à éviter cette espèce à l'avenir[131]. Les grenouilles toxiques indiquent leur toxicité en se parant de couleurs vives, une stratégie adaptative connue comme l'aposématisme. C'est le cas notamment des grenouilles de la famille des Dendrobatidae. Elles sont généralement rouges, orange ou jaunes, souvent avec des marques noires qui font contraste. Allobates zaparo n'est pas toxique, mais mime l'apparence de deux espèces toxiques qui partagent son aire de répartition pour se prémunir des prédateurs[132]. D'autres espèces, comme le Sonneur à ventre de feu (Bombina bombina), ont leurs couleurs dissuasives sous le corps. Elles les exhibent alors lorsqu'elles sont attaquées, adoptant une pose pour montrer leur ventre[5].

Certaines grenouilles, comme les Dendrobatidae, sont particulièrement toxiques. Les indigènes d'Amérique du Sud extraient le poison de ces grenouilles pour en recouvrir leurs armes pour la chasse[133], bien que peu d'espèces soient suffisamment toxiques pour être utilisées de la sorte. Au moins deux espèces non toxiques d'Amérique tropicale (Pristimantis gaigei et Lithodytes lineatus) miment l'apparence des Dendrobatidae pour se protéger elles-mêmes[134] - [135]. Certaines grenouilles obtiennent leur poison grâce aux fourmis et autres arthropodes qu'elles consomment[136]. D'autres, comme les Pseudophryne d'Australie (Pseudophryne corroboree et Pseudophryne pengilleyi), synthétisent leurs alcaloïdes elles-mêmes[137]. Les substances produites par les anoures peuvent être irritantes, hallucinogènes, neurotoxiques, vasoconstrictrices ou provoquer des convulsions. De nombreux prédateurs se sont adaptés et peuvent tolérer un degré d'empoisonnement fort avec ces substances, mais d'autres créatures, y compris les hommes qui attrapent les anoures, peuvent être fortement touchées[138].
Certains anoures se défendent en intimidant leurs prédateurs. Le Crapaud commun (Bufo bufo) adopte une posture particulière lorsqu'on l'attaque, gonflant son corps et se tenant dressé sur ses pattes de devant avec la tête baissée[139]. La Grenouille-taureau (Rana catesbeiana) se tapit au sol avec ses yeux fermés et la tête penchée vers l'avant quand elle est menacée. De cette façon les glandes parotoïdes sont dans leur position la plus efficace, les autres glandes sur son dos commencent à suinter des sécrétions toxiques et les parties de son corps les plus vulnérables sont protégées[91]. Certaines grenouilles « crient » pour surprendre le prédateur. La Rainette versicolore (Hyla versicolor) produit un bruit lourd qui rappelle parfois la Grande musaraigne Blarina brevicauda[91]. Bien que les crapauds soient évités par de nombreux prédateurs, la Couleuvre rayée (Thamnophis sirtalis) en consomme régulièrement. Les jeunes Crapauds d'Amérique (Bufo americanus) lorsqu'ils sont approchés par ce serpent font le mort : ils se couchent et restent immobiles, tactique souvent fructueuse, le serpent passant à côté du crapaud sans le détecter[140].
Répartition géographique et habitat

Les anoures sont présents sur tous les continents à l'exception de l'Antarctique, mais ne sont pas présents sur diverses îles, notamment sur celles qui sont éloignées des continents[141] - [142]. De nombreuses espèces sont restreintes à des zones géographiques limitées du fait de changements de climat ou de territoires inhospitaliers comme les étendues de mer, les chaînes montagneuses, les déserts, les forêts coupées à blanc, les routes ou diverses autres constructions humaines qui constituent des barrières infranchissables pour ces animaux[143]. Généralement, on rencontre une plus grande diversité d'anoures dans les zones tropicales que dans les zones tempérées comme l'Europe[144]. Certaines grenouilles vivent dans des habitats arides, comme les déserts, grâce à des adaptations adéquates. Ainsi les membres du genre australien Cyclorana creusent eux-mêmes des galeries souterraines un cocon imperméable à l'eau dans lequel ils peuvent estiver durant les périodes sèches. Une fois qu'il pleut, les animaux émergent et se reproduisent dès qu'elles ont trouvé une mare temporaire. Le développement des œufs et des têtards est alors très rapide en comparaison des autres espèces, ceci afin que la reproduction puisse se terminer avant que la mare temporaire ne tarisse[145]. Certaines grenouilles sont adaptées aux environnements froids. La Grenouille des bois (Rana sylvatica), dont l'habitat s'étend dans le cercle Arctique, creuse dans le sol en hiver. Bien que la plupart de son corps gèle à ce moment, elle protège ses organes vitaux des dommages grâce à la très forte concentration de glucose en leur sein[21].
Taxinomie
Environ 88 % des espèces d'amphibiens sont classées dans l'ordre des anoures[18]. Cet ordre comprend environ 4 810 espèces appartenant à 33 familles[146], parmi lesquelles les Leptodactylidae (1100 espèces), les Hylidae (800 espèces) et les Ranidae (750 espèces) sont les plus importantes[18]. Les anoures comprennent toutes les grenouilles modernes et diverses espèces fossiles. Les caractéristiques des anoures adultes comprennent neuf vertèbres présacrales ou moins, un long ilium incliné vers l'avant, la présence d'un urostyle, l'absence de queue, des membres avant plus courts que les membres arrière, le radius et l'ulna fusionnés, tout comme le tibia et le fibula, des os de la cheville allongés, l'absence d'un os préfrontal, la présence d'un os hyoïde, une mâchoire inférieure dépourvue de dents, une langue sans support, des espaces lymphatiques en dessous de la peau et un muscle, le protractor lentis, attaché au cristallin[147]. La larve des anoures, le têtard, a un unique stigmate pour respirer et sa bouche est pourvue d'un bec corné et de denticules[147]
Les grenouilles et les crapauds sont classés en trois sous-ordres : celui des Archaeobatrachia, qui inclut quatre familles de grenouilles "primitives" ; celui des Mesobatrachia, qui comprend cinq familles de grenouilles un peu plus ""évoluées"" ; et celui des Neobatrachia, de loin le plus répandu, qui contient les 24 autres familles d'anoures "modernes", dont les espèces les plus communes présentes à travers le monde. Le sous-ordre des Neobatrachia est lui-même divisé en deux superfamilles, les Hyloidea et les Ranoidea[148]. Cette classification s'appuie sur des caractéristiques morphologiques comme le nombre de vertèbres, la structure de la ceinture scapulaire et la morphologie des têtards. Alors que cette classification est largement acceptée, les liens entre les différentes familles d'anoures restent débattus[149].
Certaines espèces d'anoures s'hybrident communément. Par exemple, la Grenouille verte d'Europe (Pelophylax kl. esculentus) est un hybride entre la Petite grenouille verte (Pelophylax lessonae) et la Grenouille rieuse (Pelophylax ridibundus)[150]. Le Sonneur à ventre de feu (Bombina bombina) et le Sonneur à ventre jaune Bombina variegata sont également très proches et peuvent s'hybrider. Les hybrides sont moins fertiles que leurs parents, et on trouve une zone d'hybridation où ils sont très répandus[151].
Liste des familles
Selon Amphibian Species of the World (17 septembre 2014)[152] :
- Allophrynidae Savage, 1973
- Alsodidae Mivart, 1869
- Alytidae Fitzinger, 1843
- Aromobatidae Grant, Frost, Caldwell, Gagliardo, Haddad, Kok, Means, Noonan, Schargel & Wheeler, 2006
- Arthroleptidae Mivart, 1869
- Ascaphidae Fejérváry, 1923
- Batrachylidae Gallardo, 1965
- Bombinatoridae Gray, 1825
- Brachycephalidae Günther, 1858
- Brevicipitidae Bonaparte, 1850
- Bufonidae Gray, 1825
- Calyptocephalellidae Reig, 1960
- Centrolenidae Taylor, 1951
- Ceratobatrachidae Boulenger, 1884
- Ceratophryidae Tschudi, 1838
- Conrauidae Dubois, 1992
- Craugastoridae Hedges, Duellman & Heinicke, 2008
- Cycloramphidae Bonaparte, 1850
- Dendrobatidae Cope, 1865
- Dicroglossidae Anderson, 1871
- Eleutherodactylidae Lutz, 1954
- Heleophrynidae Noble, 1931
- Hemiphractidae Peters, 1862
- Hemisotidae Cope, 1867
- Hylidae Rafinesque, 1815
- Hylodidae Günther, 1858
- Hyperoliidae Laurent, 1943
- Leiopelmatidae Mivart, 1869
- Leptodactylidae Werner, 1896
- Limnodynastidae Lynch, 1969
- Mantellidae Laurent, 1946
- Megophryidae Bonaparte, 1850
- Micrixalidae Dubois, Ohler & Biju, 2001
- Microhylidae Günther, 1858
- Myobatrachidae Schlegel, 1850
- Nasikabatrachidae Biju & Bossuyt, 2003
- Nyctibatrachidae Blommers-Schlösser, 1993
- Odontobatrachidae Barej, Schmitz, Günther, Loader, Mahlow & Rödel, 2014
- Odontophrynidae Lynch, 1969
- Pelobatidae Bonaparte, 1850
- Pelodytidae Bonaparte, 1850
- Petropedetidae Noble, 1931
- Phrynobatrachidae Laurent, 1941
- Pipidae Gray, 1825
- Ptychadenidae Dubois, 1987
- Pyxicephalidae Bonaparte, 1850
- Ranidae Rafinesque, 1814
- Ranixalidae Dubois, 1987
- Rhacophoridae Hoffman, 1932
- Rhinodermatidae Bonaparte, 1850
- Rhinophrynidae Günther, 1859
- Scaphiopodidae Cope, 1865
- Sooglossidae Noble, 1931
- Telmatobiidae Fitzinger, 1843
Histoire évolutive
Les origines et les relations évolutives entre les trois principaux groupes d'amphibiens sont sujets à controverses. La phylogénie moléculaire s'appuyant sur l'analyse de l'ADN ribosomique réalisée en 2005 suggère que les salamandres et les cécilies sont plus proches entre eux qu'ils le sont des anoures, et que la divergence entre les trois groupes date du Paléozoïque ou du début du Mésozoïque, avant la séparation du supercontinent Pangée et peu après leur divergence d'avec les sarcoptérygiens[153]. Une autre analyse moléculaire phylogénétique a conclu que les lissamphibiens sont apparus pour la première fois il y a environ 330 millions d'années et que l'hypothèse selon laquelle les anoures dérivaient des Temnospondyli est la théorie la plus crédible. Les Neobatrachia semblent être originaires d'Afrique et d'Inde, les salamandres d'Asie de l'Est et les cécilies de la Pangée tropicale[154]. D'autres chercheurs, bien qu'ils acceptent les principaux points de cette théorie, pensent que la divergence des lissamphibiens a eu lieu au Permien, il y a moins de 300 millions d'années, une date qui est plus en rapport avec les découvertes paléontologiques[155]. Une étude complémentaire menée en 2011 sur des taxons éteints et existants étudiés du point de vue de leur morphologie et également de leur analyse moléculaire a conclu que Lissamphibia est un groupe monophylétique et qu'il devait être placé dans les Lepospondyli plutôt que parmi les Temnospondyli. L'étude postule que Lissamphibia n'est pas apparu avant la fin du Carbonifère, il y a entre 290 et 305 millions d'années. La séparation entre les anoures et les Caudata est estimée avoir eu lieu il y a 292 millions d'années, bien plus tard que la plupart des études moléculaires ne le suggèrent, et les cécilies auraient quant à eux divergé il y a 239 millions d'années[156].

En 2008, Gerobatrachus hottoni, un temnospondyle partageant à la fois des caractéristiques des grenouilles et des salamandres, est découvert au Texas. Il est daté de 290 millions d'années avant notre ère, et considéré comme un chaînon manquant, proche de l'ancêtre commun des grenouilles et des salamandres, ce qui est cohérent avec l'idée largement acceptée suivant laquelle les grenouilles et les salamandres sont plus fortement apparentées (formant un clade appelé Batrachia) qu'elles le sont avec les cécilies[157] - [158]. Le premier Salientia connu (voir ci-dessous), plus proche des grenouilles existantes que des salamandres existantes, est Triadobatrachus massinoti, vivant au début du Trias à Madagascar (il y a environ 250 millions d'années), et des fragments de Czatkobatrachus polonicus vivant à la même période en Pologne[159]. Le crâne de Triadobatrachus ressemble à celui d'une grenouille, large avec de grandes orbites oculaires, mais les fossiles ont des caractéristiques qui divergent clairement des grenouilles actuelles, notamment un corps plus long avec plus de vertèbres. La queue a des vertèbres séparées qui diffèrent du coccyx fusionné des grenouilles actuelles. Le tibia et le fibula sont également séparés, et Triadobatrachus n'était donc vraisemblablement pas un bon sauteur[159].
Salientia (du latin (la) (salio), « sauter ») est un groupe comprenant les grenouilles modernes dans l'ordre des anoures et les fossiles qui leur sont fortement apparentés (comme Triadobatrachus et Czatkobatrachus). Les principales caractéristiques de ces « proto-grenouilles » de l'ordre des Salientia incluent les 14 vertèbres présacrées (les grenouilles modernes en ont 8 ou 9), un long ilium tourné vers l'avant au niveau du pelvis, la présence d'un os pariétal et d'une mâchoire inférieure dépourvue de dents. La première grenouille fossile que l'on place dans la lignée des anoures proprement dits est Prosalirus bitis, qui vivait au début du Jurassique[3] - [160]. Elle a été découverte en 1995 dans la formation de Kayenta en Arizona, et date du début du Jurassique (entre 199,6 et 175 millions d'années avant notre ère), faisant de Prosalirus un groupe plus récent que Triadobatrachus[161]. Comme ces descendants, Prosalirus ne présente pas les pattes fortement agrandies mais possède la structure pelvique typique des grenouilles modernes. À la différence de Triadobatrachus, Prosalirus a déjà perdu presque toute sa queue[162] et était bien adapté au saut[163].
La première véritable grenouille connue est Vieraella herbstii, qui vivait au début du Jurassique. On connaît cet animal uniquement à partir des impressions fossilisées du dos et du ventre d'un animal unique, dont la taille était d'environ 33 mm. Notobatrachus degiustoi datant du milieu du Jurassique est plus jeune et date d'il y a entre 155 et 170 millions d'années. Les principales évolutions visibles sur cette espèce est le raccourcissement du corps et la perte de la queue. On pense que l'évolution des anoures modernes s'est achevée à la fin du Jurassique. Depuis, les évolutions au niveau du nombre de chromosomes ont été environ 20 fois plus rapides chez les mammifères que chez les grenouilles, indiquant que le phénomène de spéciation était plus rapide chez les mammifères[164].
Des fossiles de grenouilles ont été retrouvés sur tous les continents à l'exception de l'Antarctique mais des preuves biogéographiques suggèrent qu'ils ont également vécu en Antarctique dans une ère précédente quand le climat y était plus chaud[165].
Phylogénie
Place au sein des lissamphibiens
Phylogénie des ordres d'amphibiens modernes d'après Marjanovic & Laurin (2007)[166] :
| Lissamphibia |
| ||||||
Phylogénie interne
Phylogénie des familles basales de l'ordre Anura, d'après Pyron et Wiens (2011)[167] ainsi que Frost et al. (2006)[168] et Heinicke et al. (2009)[169] pour les noms de clades :
|
|
Les anoures et l'Homme
Statut de conservation et sauvegarde

En 2006, parmi les 4 035 espèces d'amphibiens dépendant de l'eau au cours de leur cycle de vie, 1 356 (33,6 %) étaient considérés comme menacés. Il se pourrait même que ce chiffre soit sous-estimé car pour 1 427 on ne dispose pas de suffisamment d'éléments pour établir leur statut de sauvegarde[170]. Les populations d'anoures ont décliné fortement depuis les années 1950. Plus d'un tiers des espèces sont considérées comme menacées d'extinction, et plus de 120 espèces semblent s'être éteintes depuis les années 1980[171]. Parmi ces espèces disparues on compte notamment les Rheobatrachus d'Australie et le Crapaud doré du Costa Rica. La disparition de ce dernier a une importance particulière pour les scientifiques car il vivait dans la réserve de la forêt de nuages de Monteverde et a souffert d'un crash de sa population en 1987, comme 20 autres espèces d'anoures que l'on trouve dans cette région. Cet effondrement de la population, au-delà de la normale, ne peut pas être directement relié aux activités humaines telles que la déforestation[172]. Ailleurs, la disparition de leur habitat est la principale cause de déclin des populations d'anoures, avec la pollution, le changement climatique, l'accroissement des radiations UV et l'introduction d'espèces prédatrices ou entrant en compétition avec les espèces locales[173]. Une étude canadienne conduite en 2006 a estimé que l'augmentation du trafic routier était une cause de déclin plus importante que la disparition d'habitats[174]. Des maladies infectieuses émergentes, comme la chytridiomycose et le ranavirus, dévastent également certaines populations[175] - [176].
Plusieurs spécialistes pensent que les amphibiens, dont les anoures, sont de très bons indicateurs de la bonne santé d'un écosystème du fait de leur position intermédiaire dans la chaîne alimentaire, leur peau perméable et leur vie à la fois aquatique et terrestre[177]. Il apparaît d'ailleurs que les espèces avec des œufs et des larves aquatiques sont plus sujettes au déclin que celles qui ont un développement direct sans stade larvaire[178].
Les mutations chez les grenouilles ont augmenté depuis les années 1990. Ces déformations comprennent notamment des pattes manquantes ou supplémentaires. Diverses causes sont identifiées ou considérées comme possibles, comme l'augmentation de la radiation ultraviolette qui pourrait affecter la ponte à la surface des mares, la contamination de l'eau par des pesticides et des engrais et la présence de parasites comme le trématode Ribeiroia ondatrae. Probablement que tous ces éléments contribuent d'une manière complexe à stresser les animaux, les rendant plus sensibles aux maladies et plus vulnérables aux attaques de parasites. Les malformations limitent la mobilité des animaux qui aurait donc du mal à atteindre l'âge adulte. L'augmentation du nombre de grenouilles capturées par des oiseaux risque au bout du compte d'augmenter la probabilité que les autres grenouilles soient parasitées, car le cycle des trématodes qui parasitent ces animaux est complexe et comprend des escargots de la famille des Planorbidae et les oiseaux comme hôtes intermédiaires[179] - [180].
Dans quelques rares cas, des programmes d'élevage en captivité ont été menés avec succès[181] - [182]. En 2007, l'application d'une bactérie probiotique s'est révélée pour enrayer la chytridiomycose[183]. Un projet mené actuellement, le Panama Amphibian Rescue and Conservation Project, a été développé à la suite de cette découverte pour secourir les espèces menacées par cette maladie à l'Est du Panama, et pour faire avancer la recherche sur la thérapie par les probiotiques[184] - [185]. L'Association mondiale des zoos et des aquariums a déclaré 2008 l'« Année de la grenouille » pour attirer l'attention sur les problèmes de conservation qui menacent ces animaux[186].
Le Crapaud buffle (Bufo marinus) est une espèce originaire d'Amérique du Sud et d'Amérique centrale, et capable de s'adapter facilement à d'autres environnements. Dans les années 1930 il a été introduit à Porto Rico et dans diverses autres îles des Caraïbes pour réguler les insectes nuisibles dans les champs cultivés[187]. En 1935, 3000 Crapauds buffles sont relâchés dans les plantations de canne à sucre du Queensland, en Australie, pour lutter contre des insectes ravageurs tels que Dermolepida albohirtum, dont les larves font énormément de dégâts dans les cannes. Les premiers résultats sont positifs, mais il devient par la suite clair que le crapaud se développe et perturbe l'équilibre de l'écosystème local. Ils se reproduisent librement, se multiplient et entrent en compétition avec les grenouilles locales, mangent les abeilles et d'autres invertébrés non nuisibles. Ils ont peu de prédateurs dans ce nouvel habitat, et empoisonnent les animaux domestiques, les oiseaux carnivores et les mammifères qui cherchent à les consommer. Dans plusieurs de ces pays il est maintenant considéré comme une espèce invasive nuisible, et les scientifiques cherchent un moyen de contrôler sa population[188].
Les anoures dans l'alimentation


Les cuisses de grenouilles sont consommées par l'Homme dans diverses parties du monde. Elles provenaient de petites populations sauvages, mais la surexploitation a conduit à la raréfaction de cette ressource. C'est pourquoi s'est développé l'élevage des grenouilles et un marché mondial en la matière[189]. Les principaux pays importateurs sont la France, la Belgique, le Luxembourg, et les États-Unis, tandis que les pays exportateurs sont l'Indonésie et la Chine[189]. Chaque année, entre 1 200 et 2 400 tonnes de Grenouille-taureau (Rana catesbeiana), principalement produites en Chine, sont vendues sur le marché mondial[190].
La consommation de grenouilles peut toutefois présenter des risques. Elles peuvent avoir bioaccumulé des toxines environnementales ou radionucléides (à la suite de la catastrophe de Tchernobyl par exemple). Certaines espèces ont une peau qui produit des toxines. La grenouille léopard du lac Saint-Pierre (Canada) s'est avérée contenir des PCB et du Mirex (mais en quantité à ce jour considérée comme non dangereuse pour les consommateurs). Les grenouilles peuvent transmettre divers pathogènes. Elles sont considérées avec d'autres animaux comme un des réservoirs de salmonelles (sur la peau et les intestins), y compris des souches endémiques rares chez l'homme mais pouvant poser problème. Cette souche également trouvée dans les poulaillers et effluents d'élevages de bovins et de volailles et en aval de stations d'épuration pourraient contaminer les zones humides et faire des amphibiens des réservoirs de telles souches[191].
Les grenouilles véhiculent aussi des bactéries comme Aeromonas hydrophila, responsable de la maladie des pattes rouges dont les symptômes sont des ulcères cutanés, un gonflement du ventre, une rougeur des pattes arrière et du bas-ventre. Cette bactérie est responsable de gastro-entérites et d'infections de plaies cutanées chez l'Homme[192].
Utilisation par la science
Les grenouilles sont parfois utilisées pour réaliser des dissections en cours d'anatomie dans les lycées et les universités, souvent après qu'on leur eut injecté des substances colorées pour mieux voir certaines organes. Cette pratique tend à se réduire avec les préoccupations croissantes concernant le bien-être, et des « grenouilles virtuelles » sont aujourd'hui disponibles pour des dissections[193].
Les grenouilles ont souvent servi pour des expérimentations animales au fil de l'histoire de la science. Le biologiste du XVIIIe siècle Luigi Galvani a découvert le lien entre l'électricité et le système nerveux via ses études sur des grenouilles[194]. En 1852, H. F. Stannius utilise un cœur de grenouille dans une procédure appelée ligature de Stannius pour démontrer que le ventricule et l'oreillette battent indépendamment l'un de l'autre et à des vitesses différentes[195]. La Xénope lisse (Xenopus laevis) était très utilisée dans les laboratoires dans la première moitié du XXe siècle pour réaliser des tests de grossesse. Un échantillon d'urine venant d'une femme enceinte injecté dans une grenouille femelle provoque en effet sa ponte, comme l'a observé le zoologiste anglais Lancelot Hogben. Ceci est du a une hormone, l'hormone gonadotrophine chorionique, présente en grande quantité dans l'urine d'une femme durant sa grossesse[196]. En 1952, Robert Briggs et Thomas J. King clonent une grenouille à partir de cellules somatiques. La même technique est plus tard utilisée pour créer la brebis Dolly, et leur expérience constitue la première transplantation nucléaire réussie pour un animal supérieur[197].
Les grenouilles sont utilisées pour les recherches sur le clonage et d'autres branches de l'embryologie. Bien que d'autres méthodes soient aujourd'hui disponibles pour réaliser des tests de grossesse, les biologistes continuent à utiliser Xenopus comme un organisme modèle dans la biologie du développement car ses embryons sont gros et faciles à manipuler, elles peuvent être obtenues facilement et elles sont faciles à élever en laboratoire[198]. Xenopus laevis est tout de même de plus en plus souvent remplacée par une grenouille qui lui est apparentée, Xenopus tropicalis, plus petite et qui a l'avantage d'atteindre sa maturité sexuelle à cinq mois plutôt qu'un ou deux ans pour X. laevis[199], ce qui permet d'étudiant plus facilement les évolutions entre les générations. Le génome de X. tropicalis est séquencé[200].
Comme les toxines produites par les anoures sont extraordinairement diverses, elles intéressent fortement les biochimistes qui y voient une « pharmacie naturelle ». L'alcaloïde épibatidine, un analgésique 200 fois plus puissant que la morphine, est par exemple rencontré chez une espèce de Dendrobatidae. D'autres substances isolées sur la peau des grenouilles pourraient apporter une résistance aux infections par le VIH[201]. Les Dendrobatidae sont l'objet de recherches pour évaluer leur potentiel thérapeutique[202].
Utilisation des sécrétions toxiques

On a longtemps pensé que les Mésoaméricains pré-colombiens utilisaient les sécrétions toxiques du crapaud-buffle comme hallucinogène, mais ils utilisaient plus probablement les substances produites par Bufo alvarius. Celles-ci comprennent la bufoténine (5-MeO-DMT), un psychotrope utilisé aujourd'hui comme stupéfiant. Généralement, les sécrétions de la peau étaient séchées et fumées[203]. La prise illicite de drogue en léchant un crapaud a été rapportée par les médias mais il pourrait s'agir d'une légende urbaine[204].
Les exsudats provenant de la peau du Phyllobate terrible (Phyllobates terribilis) étaient traditionnellement utilisés par les indigènes pour empoisonner les flèches qu'ils utilisaient pour la chasse. L'extrémité du projectile était frottée sur le dos d'une grenouille et la flèche était ensuite lancée via une sarbacane. La combinaison des deux alcaloïdes toxiques que sont la batrachotoxine et la homobatrachotoxine est extrêmement puissante, et une grenouille peut fournir suffisamment de poison pour tuer 22 000 souris[205]. Deux autres espèces, le Phyllobate à bande dorée (Phyllobates aurotaenia) et le phyllobate bicolore (Phyllobates bicolor), étaient utilisées de la même manière. Elles sont toutefois moins toxiques et moins abondantes que le Phyllobate terrible. Ces grenouilles étaient empalées à la pointe d'un bâton et chauffée au-dessus d'un effet pour en extraire le maximum de poison pour recouvrir les flèches[205].
Les anoures dans la culture populaire

Les grenouilles et crapauds tiennent une place importante dans le folklore, les contes de fées et la culture populaire. Ils tendent à être dépeints comme laids et maladroits, mais avec des talents cachés. Les exemples sont nombreux, parmi lesquels le personnage du Roi Grenouille ou Henri de Fer, Michigan J. Frog ou Kermit la grenouille. Le dessin animé One Froggy Evening de la Warner Brothers met en scène Michigan J. Frog, qui ne fera que danser et chanter pour le travailleur de démolition qui ouvre la capsule de temps, mais ne fait rien en public[206]. Le Roi grenouille est un conte de fées des frères Grimm sur une grenouille qui se transforme en un beau prince après avoir sauvé la balle d'or de la princesse, et l'avoir suivi dans son palais[207]. Kermit la grenouille est un personnage consciencieux et discipliné du Muppet Show et de 1, rue Sésame (Sesame Street), dans lequel il est ouvertement amical et très talentueux[208].
Les crapauds ont une réputation plus sinistre. Ils sont associés dans le folklore européen aux sorcières, et on pensait qu'ils avaient des pouvoirs magiques. Les sécrétions toxiques de leur peau étaient utilisées dans des potions maléfiques, mais ont également été mises à profit pour créer des remèdes magiques pour les maladies humaines et animales. Ils ont été associés au diable ; dans Le Paradis perdu de John Milton, Satan est ainsi représenté sous la forme d'un crapaud qui verse du poison dans l'oreille d'Eve[209]. Dans la Bible, la deuxième des dix plaies d'Égypte est l'invasion des terres par des milliers de ces batraciens. D'après les scientifiques qui se sont penchés sur cet évènement, le phénomène pourrait s'expliquer par une sécheresse ou par l'empoisonnement des eaux du Nil. En effet, dans des situations de stress, ces animaux sont capables d'accélérer leur développement pour fuir plus vite leur milieu, d'où une explosion de leur nombre.
Les Moche, ancien peuple du Pérou, adoraient les animaux, et les grenouilles étaient souvent représentées dans leur art[210]. Au Panama, une légende locale raconte que la bonne fortune viendrait à la personne qui trouverait une grenouille dorée. Certains croyaient que lorsque l'une de ces grenouilles mourrait, elle se transformait en un talisman d'or connue sous le nom de Huaca. Aujourd'hui, en dépit de l'extinction de l'espèce à l'état sauvage, les grenouilles dorées du Panama demeure un symbole culturel important et sont représentées sur des molas en tissu décoratif faits par le peuple Kuna[211].
Publication originale
- Fischer von Waldheim, 1813 : Zoognosia tabulis synopticis illustrata : in usum praelectionum Academiae imperialis medico-chirugicae mosquensis edita, vol. 1 (texte intégral).
Annexes
Bibliographie
- (en) Ellin Beltz, Frogs: Inside their Remarkable World, Firefly Books, (ISBN 1-55297-869-9)
- (en) H.G. Cogger, R.G. Zweifel et D. Kirschner, Encyclopedia of Reptiles & Amphibians Second Edition, Fog City Press, (ISBN 1-877019-69-0)
- (en) R. Estes et O. A. Reig., « The early fossil record of frogs: a review of the evidence. », Evolutionary Biology of the Anurans: Contemporary Research on Major Problems, Columbia, University of Missouri Press, , p. 11–63.
- (en) Carmela Gissi, Diego San Mauro, Graziano Pesole et Rafael Zardoya, « Mitochondrial phylogeny of Anura (Amphibia): A case study of congruent phylogenetic reconstruction using amino acid and nucleotide characters », Gene, vol. 366, no 2, , p. 228–237 (PMID 16307849, DOI 10.1016/j.gene.2005.07.034)
- (en) J. A Holman, Fossil Frogs and Toads of North America, Indiana University Press, (ISBN 0-253-34280-5)
- (en) Diego San Mauro, Miguel Vences, Marina Alcobendas, Rafael Zardoya et Axel Meyer, « Initial diversification of living amphibians predated the breakup of Pangaea », American Naturalist, vol. 165, no 5, , p. 590–599 (PMID 15795855, DOI 10.1086/429523, lire en ligne)
- Françoise Serre-Collet, Sur la piste des reptiles et des amphibiens: Serpents, grenouilles, lézards..., sachez les reconnaître, Dunod, coll. « L'amateur de nature », , 192 p. (ISBN 9782100593859)
- (en) Robert C. Stebbins et Nathan W. Cohen, A Natural History of Amphibians, Princeton University Press, (ISBN 0-691-03281-5)
- (en) M. J. Tyler, Australian Frogs A Natural History, Reed Books, (ISBN 0-7301-0468-0)
Liens externes
- (en) Référence Amphibian Species of the World : Anura Fischer von Waldheim, 1813 (consulté le )
- (en) Référence Animal Diversity Web : Anura (consulté le )
- (en) Référence Catalogue of Life : Anura (consulté le )
- (en) Référence Fauna Europaea : Anura (consulté le )
- (fr+en) Référence ITIS : Anura (consulté le )
- (en) Référence NCBI : Anura (taxons inclus) (consulté le )
- (en) Référence Tree of Life Web Project : Anura (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023
- (en) Référence Wild Herps : photographies de Anura (consulté le )
- L'encyclopédie des Anoures
Notes et références
- Daniel Richard, André Beaumont et Pierre Cassier, Biologie animale - Les Cordés - 9e édition : Anatomie comparée des vertébrés : Anatomie comparée des vertébrés, Dunod, coll. « Sciences de la vie », , 688 pages (ISBN 978-2-10-054131-7).
- « Anoure », Centre National de Ressources Textuelles et Lexicales (consulté le ).
- (en) David C. Cannatella, « Salientia », sur Tree of Life Web Project, (consulté le ).
- [Badger & Netherton 1995] (en) David Badger et John Netherton, Frogs, Airlife Publishing Ltd, (ISBN 978-1-85310-740-5), p. 19.
- (en) Sergius L. Kuzmin, « Bombina bombina », sur AmphibiaWeb, Berkeley, Université de Californie, (consulté le ).
- (en) K Lips, F. Solís, R. Ibáñez, C. Jaramillo et Q. Fuenmayor, « Atelopus zeteki », sur IUCN Red List of Threatened Species. Version 2012.1, (consulté le ).
- Marie Élodie Clément, Les maladies infectieuses et parasitaires des amphibiens, École vétérinaire de Maison Alfort, Thèse vétérinaire, (lire en ligne), p. 4-13.
- « Anoures », Encyclopédie Larousse, Larousse, 1971-1976, p. 805 (lire en ligne).
- (en) F. Flam, « Finding earliest true frog will help paleontologists understand how frog evolved its jumping ability », Knight Ridder/Tribune News Service via HighBeam Research, (consulté le ).
- (en) Maurice Burton, The Observer's Book of British Wild Animals, Frederick Warne & Co, (ISBN 978-0-7232-1503-5), p. 204–209.
- (en) R. A. Relyea, « The impact of insecticides and herbicides on the biodiversity and productivity of aquatic communities », Ecological Applications, vol. 15, no 2, , p. 618–627 (DOI 10.1890/03-5342).
- (en) R.A. Relyea, « The lethal impact of Roundup on aquatic and terrestrial amphibians », Ecological Applications, vol. 15, no 4, , p. 1118–1124 (DOI 10.1890/04-1291).
- (en) T. B. Hayes, A. Collins, M. Lee, M. Mendoza, N. Noriega, A. A. Stuart et A. Vonk, « Hermaphroditic, demasculinized frogs after exposure to the herbicide atrazine at low ecologically relevant doses », Proceedings of the National Academy of Sciences of the United States of America, vol. 99, no 8, , p. 5476–5480 (PMID 11960004, PMCID 122794, DOI 10.1073/pnas.082121499).
- (en) P. T. J. Johnson, J. M. Chase, K. L. Dosch, R. B. Hartson, J. A. Gross, D. J. Larson, D. R. Sutherland et S. R. Carpenter, « Aquatic eutrophication promotes pathogenic infection in amphibians », Proceedings of the National Academy of Sciences of the United States of America, vol. 104, no 40, , p. 15781–15786 (PMID 17893332, PMCID 2000446, DOI 10.1073/pnas.0707763104).
- (en) « Tiny frog claimed as world's smallest vertebrate », The Guardian, (lire en ligne, consulté le ).
- AFP, « Le plus petit vertébré du monde est... une grenouille », Le Point, (consulté le ).
- (en) R. P. Levine, J. A. Monroy et E. L. Brainerd, « Contribution of eye retraction to swallowing performance in the northern leopard frog, Rana pipiens », Journal of Experimental Biology, vol. 207, no Pt 8, , p. 1361–1368 (PMID 15010487, DOI 10.1242/jeb.00885).
- (en) Frederick Harvey Pough, Robin M. Andrews, John E. Cadle, Martha L. Crump, Alan H. Savitsky et Kentwood D. Wells, Herpetology: Third Edition, Benjamin Cummings, (ISBN 0-13-100849-8).
- « Les Amphibiens », Université de Jussieu (consulté le ).
- (en) Kevin Minott, « How frogs jump », National Geographic, (consulté le ).
- (en) P. Tesler, « The amazing adaptable frog », Exploratorium : The museum of science, art and human perception, (consulté le ).
- (en) « Litoria caerulea », James Cook University (consulté le ).
- (en) S. B. Emerson et D. Diehl, « Toe pad morphology and mechanisms of sticking in frogs », Biological Journal of the Linnean Society, vol. 13, no 3, , p. 199–216 (DOI 10.1111/j.1095-8312.1980.tb00082.x).
- (en) M. B. Harvey, A. J. Pemberton et E. N. Smith, « New and poorly known parachuting frogs (Rhacophoridae : Rhacophorus) from Sumatra and Java », Herpetological Monographs, vol. 16, , p. 46–92 (DOI 10.1655/0733-1347(2002)016[0046:NAPKPF]2.0.CO;2).
- (en) « Couch's spadefoot (Scaphiopus couchi) », Arizona-Sonora Desert Museum (consulté le ).
- (en) M. Walker, « Legless frogs mystery solved », BBC News, (lire en ligne).
- Daniel Richard, André Beaumont et Pierre Cassier, Biologie animale - Les Cordés - 9e édition : Anatomie comparée des vertébrés : Anatomie comparée des vertébrés, Dunod, coll. « Sciences de la vie », , 688 p. (ISBN 978-2-10-054131-7), p. 527.
- Stebbins et Cohen, p. 10-14.
- (en) S. W. Frost, « Notes on feeding and molting in frogs », The American Naturalist, vol. 66, no 707, , p. 530–540 (DOI 10.1086/280458, JSTOR 2456779).
- Badger & Netherton 1995, p. 27.
- (en) H. R. Smyth, Amphibians and Their Ways, Macmillan, (ISBN 978-0-02-612190-3).
- (en) M. C. Dickerson, The Frog Book: North American Frogs and Toads, Dover Publications, (ISBN 978-0-486-21973-8).
- (en) D. C. Blackburn, « Trichobatrachus robustus », AmphibiaWeb, (consulté le ).
- (en) W. H. Wente et J. B. Phillips, « Fixed green and brown color morphs and a novel color‐changing morph of the Pacific tree frog Hyla regilla », The American Naturalist, vol. 162, no 4, , p. 461–473 (PMID 14582008, DOI 10.1086/378253, JSTOR 10.1086/378253).
- (en) John Kimball, « Vertebrate Lungs: Frog Lungs », Kimball's Biology Pages, (consulté le ).
- (en) Adam Boisvert, « Barbourula kalimantanensis », AmphibiaWeb, Berkeley, Université de Californie, (consulté le ).
- « Une grenouille sans poumons intrigue les scientifiques », sur ladepeche.fr (consulté le ).
- (en) John Kimball, « Animal Circulatory Systems: Three Chambers: The Frog and Lizard », Kimball's Biology Pages, (consulté le ).
- Peter H. Raven, Kenneth A. Mason, Jonathan B. Losos et Susan S. Singer, Biologie générale, De Boeck Supérieur, , 1406 p. (ISBN 9782804163051), p. 1024.
- (en) Deborah Lee, « Telmatobius culeus », AmphibiaWeb, Berkeley, Université de Californie, (consulté le ).
- (en) « Frog Digestive System », TutorVista.com, (consulté le )
- (en) R. L. Dorit, W. F. Walker et R. D. Barnes, Zoology, Saunders College Publishing, (ISBN 0-03-030504-7), p. 849
- (en) « Frog's internal systems », TutorVista.com, (consulté le )
- (en) Dorota, « Life cycle of a frog », Frogland, (consulté le )
- (en) David M. Sever et Nancy L. Staub, « Hormones, sex accessory structures, and secondary sexual characteristics in amphibians », Hormones and Reproduction of Vertebrates – Vol 2: Amphibians (consulté le ), p. 83–98
- (en) Michel Laurin et Jacques A. Gauthier, « Amniota », Tree of Life Web Project, (consulté le )
- [Howard & Rogers 1995] (en) Ian, P. Howard et Brian J. Rogers, Binocular Vision and Stereopsis, Oxford University Press, , sur books.google.co.uk (ISBN 0195084764, lire en ligne), p. 651.
- Badger & Netherton 1995, p. 31–35.
- [Muntz 1964] (en) W. R. A. Muntz, Vision in frogs, San Francisco, W. H. Freeman, coll. « Scientific American Offprints », , 10 p. (OCLC 15304238).
- Badger & Netherton 1995, p. 38.
- Stebbins et Cohen, p. 67-69.
- [Armstrong & Roberts 1998] (en) Cecilia E. Armstrong et William M. Roberts, « Electrical properties of frog saccular hair cells: distortion by enzymatic dissociation », Journal of Neuroscience, vol. 18, no 8, , p. 2962–2973 (PMID 9526013, lire en ligne [sur _ _ _], consulté en ).
- [Narins, Wilson & Mann 2013] Peter M. Narins, Maria Wilson et David Mann, « Ultrasound Detection in Fishes and Frogs: Discovery and Mechanisms », dans C. Köppl et al (éds.), Insights from Comparative Hearing Research,Chapter: Ultrasound Detection in Fishes and Frogs: Discovery and Mechanisms, , sur researchgate.net (lire en ligne).
- [Foratto et al. 2021] Roseli Foratto, Diego Llusia, Luis Felipe Toledo et Lucas Rodriguez Forti, « Treefrogs adjust their acoustic signals in response to harmonics structure of intruder calls », Behavioral Ecology, vol. 32, no 3, , p. 416–427 (résumé).
- (en) « Bullfrog », Ohio Department of Natural Resources (consulté le )
- (en) Gary Nafis, « Ascaphus truei: Coastal Tailed Frog », California Herps, (consulté le )
- (en) Debjani Roy, « Communication signals and sexual selection in amphibians », Current Science, vol. 72, , p. 923–927 (lire en ligne [PDF])
- (en) H. C. Gerhardt, « The evolution of vocalization in frogs and toads », Annual Review of Ecology and Systematics, vol. 25, , p. 293–324 (DOI 10.1146/annurev.es.25.110194.001453)
- Badger & Netherton 1995, p. 39–44.
- (en) Bill Jr. Hilton, Jug-o-Rum: Call of the Amorous Bullfrog, vol. 1, Hilton Pond Center for Piedmont Natural History, coll. « The Piedmont Naturalist »,
- (en) Pat Nash, « The RRRRRRRRiveting Life of Tree Frogs », sur beachwatchers.wsu.edu, (consulté le )
- (en) Aristophanes, « The Frogs », sur records.viu.ca (consulté le )
- (en) Rick Emmer, « How do frogs survive winter? Why don't they freeze to death? », Scientific American, (lire en ligne, consulté le )
- (en) Sara M. Kayes, Rebecca L. Cramp et Craig E. Franklin, « Metabolic depression during aestivation in Cyclorana alboguttata », Comparative Biochemistry and Physiology – Part A: Molecular & Integrative Physiology, vol. 154, no 4, , p. 557–563 (DOI 10.1016/j.cbpa.2009.09.001)
- (en) N. J. Hudson, S. A. Lehnert, A. B. Ingham, B. Symonds, C. E. Franklin et G. S. Harper, « Lessons from an estivating frog: sparing muscle protein despite starvation and disuse », American Journal of Physiology, vol. 290, no 3, , R836–R843 (PMID 16239372, DOI 10.1152/ajpregu.00380.2005)
- (en) « Top 10 best jumper animals », Scienceray (consulté le )
- (en) R. S. James et R. S. Wilson, « Explosive jumping: extreme morphological and physiological specializations of Australian rocket frogs (Litoria nasuta) », Physiological and Biochemical Zoology, vol. 81, no 2, , p. 176–185 (PMID 18190283, DOI 10.1086/525290)
- (en) S. Nauwelaerts, J. Schollier et P. Aerts, « A functional analysis of how frogs jump out of water », Biological Journal of the Linnean Society, vol. 83, no 3, , p. 413–420 (DOI 10.1111/j.1095-8312.2004.00403.x)
- Badger & Netherton 1995, p. 51.
- (en) H. C. Astley et T. J. Roberts, « Evidence for a vertebrate catapult: elastic energy storage in the plantaris tendon during frog jumping », Biology Letters, vol. 8, no 3, , p. 386–389 (PMID 22090204, PMCID 3367733, DOI 10.1098/rsbl.2011.0982)
- (en) J. Scott, « The locust jump: an integrated laboratory investigation », Advances in Physiology Education, vol. 29, no 1, , p. 21–26 (PMID 15718379, DOI 10.1152/advan.00037.2004)
- (en) « Anura », Encyclopædia Britannica Online (consulté le )
- (en) H. S. Fitch, « An ecological study of the collared lizard (Crotaphytus collaris) », University of Kansas Publications, Museum of Natural History, vol. 8, , p. 213–274
- (en) M. Walton et B. D. Anderson, « The aerobic cost of saltatory locomotion in the fowler's toad (Bufo woodhousei fowleri) », Journal of Experimental Biology, vol. 136, , p. 273–288 (PMID 3404074, lire en ligne)
- (en) A. N. Ahn, E. Furrow et A. A. Biewener, « Walking and running in the red-legged running frog, Kassina maculata », Journal of Experimental Biology, vol. 207, no Pt 3, , p. 399–410 (PMID 14691087, DOI 10.1242/jeb.00761)
- (en) M. Pickersgill, A. Schiøtz, K. Howell et L. Minter, « Kassina maculata », IUCN Red List of Threatened Species, (consulté le )
- (en) « Pipidae », AmphibiaWeb, University of California, Berkeley (consulté le )
- (en) W. E. Duellman et G. R. Zug, « Anura: From tadpole to adult », Encyclopædia Britannica (consulté le )
- (en) C. Radhakrishnan et K. C. Gopi, « Extension of range of distribution of Nasikabatrachus sahyadrensis Biju & Bossuyt (Amphibia : Anura : Nasikabatrachidae) along Western Ghats, with some insights into its bionomics », Current Science, vol. 92, no 2, , p. 213–216 (ISSN 0011-3891, lire en ligne)
- (en) Eugenia Farrar et Jane Hey, « Spea bombifrons », AmphibiaWeb, University of California, Berkeley (consulté le )
- (en) « Scaphiopodidae », AmphibiaWeb, University of California, Berkeley (consulté le )
- (en) Dale Roberts et Jean-Marc Hero, « Heleioporus albopunctatus », IUCN Red List of Threatened Species, (consulté le )
- (en) Marc Staniszewski, « Madagascan Burrowing Frogs: Genus: Scaphiophryne (Boulenger, 1882) », sur amphibian.co.uk, (consulté le )
- (en) M. Venesci, C. J. Raxworthy, R. A. Nussbaum et F. Glaw, « A revision of the Scaphiophryne marmorata complex of marbled toads from Madagascar, including the description of a new species », Herpetological Journal, vol. 13, , p. 69–79 (lire en ligne)
- (en) W. Federle, W. J. P. Barnes, W. Baumgartner, P. Drechsler et J. M. Smith, « Wet but not slippery: boundary friction in tree frog adhesive toe pads », Journal of the Royal Society Interface, vol. 3, no 10, , p. 689–697 (DOI 10.1098/rsif.2006.0135).
- (en) Doris Mabel Cochran, Living Amphibians of the World, Doubleday, (ISBN 978-0-241-90338-4, présentation en ligne), p. 112.
- (en) « Phyllomedusa ayeaye », AmphibiaWeb (consulté le ).
- (en) Sharon B. Emerson et M. A. R. Koehl, « The interaction of behavioral and morphological change in the evolution of a novel locomotor type: "flying frogs », Evolution, vol. 44, no 8, , p. 1931–1946 (DOI 10.2307/2409604, JSTOR 2409604)
- (en) Sunny Shah et Rachna Tiwari, « Rhacophorus nigropalmatus », AmphibiaWeb, Berkeley, Université de Californie, (consulté le )
- (en) « Wallace's Flying Frog Rhacophorus nigropalmatus », National Geographic: Animals (consulté le )
- Stebbins et Cohen, p. 154-162
- (en) N. B. Davies et T. R. Halliday, « Deep croaks and fighting assessment in toads Bufo bufo », Nature, vol. 274, no 5672, , p. 683–685 (DOI 10.1038/274683a0)
- (en) David R. Long, « Energetics and reproduction in female Scaphiopus multiplicatus from Western Texas », Journal of Herpetology, vol. 23, no 2, , p. 176–179 (DOI 10.2307/1564026, JSTOR 1564026)
- (en) Alan Channing et Kim M. Howell, Amphibians of East Africa, Comstock Publishing, (ISBN 0-8014-4374-1), p. 104–117
- (en) L. Sandberger, A. Hillers, J. Doumbia, N-S. Loua, C. Brede et M-O. Rödel, « Rediscovery of the Liberian Nimba toad, Nimbaphrynoides liberiensis (Xavier, 1978) (Amphibia: Anura: Bufonidae), and reassessment of its taxonomic status », Zootaxa, Magnolia Press, vol. 4355, , p. 56–68 (ISSN 1175-5334, lire en ligne)
- (en) Perry W. Gilbert, « Observations on the eggs of Ambystoma maculatum with especial reference to the green algae found within the egg envelopes », Ecology, vol. 23, no 2, , p. 215–227 (DOI 10.2307/1931088, JSTOR 1931088)
- (en) Bruce Waldman et Michael J. Ryan, « Thermal advantages of communal egg mass deposition in wood frogs (Rana sylvatica) », Journal of Herpetology, vol. 17, no 1, , p. 70–72 (DOI 10.2307/1563783, JSTOR 1563783)
- (en) Alberto R. Estrada et S. Blair Hedges, « At the lower size limit in tetrapods: a new diminutive frog from Cuba (Leptodactylidae: Eleutherodactylus) », Copeia, vol. 1996, no 4, , p. 852–859 (DOI 10.2307/1447647, JSTOR 1447647)
- (en) Kellie Whittaker et Peera Chantasirivisal, « Leptodactylus pentadactylus », AmphibiaWeb, Berkeley, Université de Californie, (consulté le )
- (en) Kellie Whittaker, « Agalychnis callidryas », AmphibiaWeb, Berkeley, Université de Californie, (consulté le )
- (en) K.M. Warkentin, « Adaptive plasticity in hatching age: a response to predation risk trade-offs », Proceedings of the National Academy of Sciences, vol. 92, no 8, , p. 3507–3510 (DOI 10.1073/pnas.92.8.3507)
- Stebbins et Cohen, p. 179-194
- (en) Deepthi D. Wickramasinghe, Kerri L. Oseen et Richard J. Wassersug, « Ontogenetic Changes in Diet and Intestinal Morphology in Semi-Terrestrial Tadpoles of Nannophrys ceylonensis (Dicroglossidae) », Copeia, vol. 2007, no 4, , p. 1012–1018 (JSTOR 25140719)
- (en) Peter Janzen, « Nannophrys ceylonensis », AmphibiaWeb, Université de Californie, Berkeley, (consulté le )
- (en) « Tadpole Locomotion: Axial Movement and Tail Functions in a Largely Vertebraeless Vertebrate »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), sur intl-icb.oxfordjournals.org (consulté le )
- (en) Martha L. Crump, « Cannibalism by younger tadpoles: another hazard of metamorphosis », Copeia, American Society of Ichthyologists and Herpetologists, vol. 4, no 4, , p. 1007–1009 (DOI 10.2307/1445301, JSTOR 1445301)
- Stebbins et Cohen, p. 173-175
- (en) Boris Ivan Balinsky, « Animal development: Metamorphosis », Encyclopedia Britannica (consulté le )
- Serre-Collet 2013, p. 49
- (en) Alexis C. Noel, Hao-Yuan Guo, Mark Mandica et David L. Hu, « Frogs use a viscoelastic tongue and non-Newtonian saliva to catch prey », Journal of The Royal Society Interface, vol. 14, no 127, , p. 20160764 (ISSN 1742-5689 et 1742-5662, PMID 28148766, DOI 10.1098/rsif.2016.0764, lire en ligne, consulté le )
- (en) H. R. Da Silva et X. C. De Britto-Pereira, « How much fruit do fruit-eating frogs eat? An investigation on the diet of Xenohyla truncata (Lissamphibia: Anura: Hylidae) », Journal of Zoology, vol. 270, no 4, , p. 692–698 (DOI 10.1111/j.1469-7998.2006.00192.x)
- (en) Donna Graham, « Northern leopard frog (Rana pipiens) », An Educator's Guide to South Dakota's Natural Resources (consulté le )
- (en) Stuart L. Pimm, « The structure of food webs », Theoretical Population Biology, vol. 16, no 2, , p. 144–158 (PMID 538731, DOI 10.1016/0040-5809(79)90010-8, lire en ligne)
- (en) K. R. Matthews et C. Miaud, « A skeletochronological study of the age structure, growth, and longevity of the mountain yellow-legged frog, Rana muscosa, in the Sierra Nevada, California », Copeia, vol. 2007, no 4, , p. 986–993 (ISSN 0045-8511, DOI 10.1643/0045-8511(2007)7[986:ASSOTA]2.0.CO;2, lire en ligne)
- (en) Frank Slavens et Kate Slavens, « Frog and toad index », Longevity (consulté le )
- (en) KB Storey, « Life in a frozen state: adaptive strategies for natural freeze tolerance in amphibians and reptiles », American Journal of Physiology, vol. 258, no 3 Pt 2, , p. 559–568 (PMID 2180324)
- (en) M. L. Crump, « Parental care among the Amphibia », Advances in the Study of Behavior, advances in the Study of Behavior, vol. 25, , p. 109–144 (ISBN 978-0-12-004525-9, DOI 10.1016/S0065-3454(08)60331-9)
- (en) J. L. Brown, V. Morales et K. Summers, « A key ecological trait drove the evolution of biparental care and monogamy in an amphibian », American Naturalist, vol. 175, no 4, , p. 436–446 (PMID 20180700, DOI 10.1086/650727)
- (en) Jennifer A. Sheridan et Joanne F. Ocock, « Parental care in Chiromantis hansenae (Anura: Rhacophoridae) », Copeia, vol. 2008, no 4, , p. 733–736 (DOI 10.1643/CH-07-225)
- (en) J. L. Brown, V. Morales et K. Summers, « Divergence in parental care, habitat selection and larval life history between two species of Peruvian poison frogs: An experimental analysis », Journal of Evolutionary Biology, vol. 21, no 6, 2008a, p. 1534–1543 (PMID 18811668, DOI 10.1111/j.1420-9101.2008.01609.x)
- (en) T. Grant, D. R. Frost, J. P. Caldwell, R. Gagliardo, C. F. B. Haddad, P. J. R. Kok, D. B. Means, B. P. Noonan, W. E. Schargel et W. Wheeler, « Phylogenetic systematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae) », Bulletin of the American Museum of Natural History, vol. 299, , p. 1–262 (ISSN 0003-0090, DOI 10.1206/0003-0090(2006)299[1:PSODFA]2.0.CO;2)
- (en) René van Wijngaarden et Federico Bolaños, « Parental care in Dendrobates granuliferus (Anura: Dendrobatidae), with a description of the tadpole », Journal of Herpetology, vol. 26, no 1, , p. 102–105 (DOI 10.2307/1565037, JSTOR 1565037)
- (en) María Claudia Fandiño, Horst Lüddecke et Adolfo Amézquita, « Vocalisation and larval transportation of male Colostethus subpunctatus (Anura: Dendrobatidae) », Amphibia-Reptilia, vol. 18, no 1, , p. 39–48 (DOI 10.1163/156853897X00297)
- (en) « Common midwife toad, Alytes obstetricans », Repties et Amphibiens de France (consulté le )
- (en) « Proteins of frog foam nests », Université de Glasgow (consulté le )
- (en) Laura Dalgetty et Malcolm W. Kennedy, « Building a home from foam—túngara frog foam nest architecture and three-phase construction process », Biology Letters, vol. 6, no 3, , p. 293–296 (PMID 20106853, PMCID 2880057, DOI 10.1098/rsbl.2009.0934)
- (en) « Assa darlingtoni », Frogs Australia Network, (consulté le )
- (en) E. Semeyn, « Rheobatrachus silus », Animal Diversity Web, University of Michigan Museum of Zoology, (consulté le )
- (en) Fran Sandmeier, « Rhinoderma darwinii », AmphibiaWeb, (consulté le )
- (en) Henry M. Wilbur, Peter J. Morin et Reid N. Harris, « Anuran Responses to Salamander Predation », BioScience, Oxford University Press on behalf of the American Institute of Biological Sciences, vol. 34, no 4, , p. 257 (DOI 10.2307/1309466, lire en ligne)
- (en) George T. Barthalmus et William J. Zielinski, « Xenopus skin mucus induces oral dyskinesias that promote escape from snakes », Pharmacology Biochemistry and Behavior, vol. 30, no 4, , p. 957–959 (PMID 3227042, DOI 10.1016/0091-3057(88)90126-8)
- (en) Catherine R. Darst et Molly E. Cummings, « Predator learning favours mimicry of a less-toxic model in poison frogs », Nature, vol. 440, no 7081, , p. 208–211 (PMID 16525472, DOI 10.1038/nature04297)
- (en) C. W. Myers et J. W. Daly, « Dart-poison frogs », Scientific American, vol. 248, no 2, , p. 120–133 (PMID 6836257, DOI 10.1038/scientificamerican0283-120)
- (en) J. M. Savage, The Amphibians and Reptiles of Costa Rica, University of Chicago Press, (ISBN 0-916984-16-8)
- (en) W. E. Duellman, « The Biology of an Equatorial Herpetofauna in Amazonian Ecuador », University of Kansas Museum of Natural History Miscellaneous Publication, vol. 65,
- (en) R.A. Saporito, H. M. Garraffo, M. A. Donnelly, A. L. Edwards, J. T. Longino et J. W. Daly, « Formicine ants: an arthropod source for the pumiliotoxin alkaloids of dendrobatid poison frogs », Proceedings of the National Academy of Sciences, vol. 101, no 21, , p. 8045–8050 (PMID 15128938, PMCID 419554, DOI 10.1073/pnas.0402365101)
- (en) B. P. Smith, M. J. Tyler, T. Kaneko, H. M. Garraffo, T. F. Spande et J. W. Daly, « Evidence for biosynthesis of pseudophrynamine alkaloids by an Australian myobatrachid frog (Pseudophryne) and for sequestration of dietary pumiliotoxins », Journal of Natural Products, vol. 65, no 4, , p. 439–47 (PMID 11975476, DOI 10.1021/np010506a)
- (en) T. Grant, « Poison Dart Frog Vivarium », American Museum of Natural History (consulté le )
- (en) Nicholas Arnold et Denys Ovenden, Reptiles and Amphibians of Britain and Europe, Harper Collins Publishers, (ISBN 0-00-219964-5), p. 73–74
- (en) Floyd E. Hayes, « Antipredator behavior of recently metamorphosed toads (Bufo a. americanus) during encounters with garter snakes (Thamnophis s. sirtalis) », Copeia, American Society of Ichthyologists and Herpetologists, vol. 1989, no 4, , p. 1011–1015 (DOI 10.2307/1445987, JSTOR 1445987)
- (en) « Freaky Frogs », National Geographic Explorer (consulté le )
- (en) Vance Ferrell, « Geographical Distribution », Evolution Encyclopedia, Volume 3, Evolution Facts, (consulté le )
- (en) Paddy Ryan, « Story: Frogs », The Encyclopedia of New Zealand, (consulté le )
- (en) Chris Dahl, Vojtech Novotny, Jiri Moravec et Stephen J. Richards, « Beta diversity of frogs in the forests of New Guinea, Amazonia and Europe: contrasting tropical and temperate communities », Journal of Biogeography, vol. 36, no 5, , p. 896–904 (DOI 10.1111/j.1365-2699.2008.02042.x)
- (en) « Cyclorana platycephala », Frogs Australia Network, (consulté le )
- (en) Guillaume Lecointre et Hervé Le Guyader, Classification phylogénétique du vivant, Belin, , 512 p.
- (en) David Cannatella, « Anura », Tree of Life web project, (consulté le )
- (en) L.S. Ford et D. C. Cannatella, « The major clades of frogs », Herpetological Monographs, vol. 7, , p. 94–117 (DOI 10.2307/1466954, JSTOR 1466954)
- (en) J. Faivovich, C. F. B. Haddad, P. C. A. Garcia, D. R. Frost, J. A. Campbell et W. C. Wheeler, « Systematic review of the frog family Hylidae, with special reference to Hylinae: Phylogenetic analysis and revision », Bulletin of the American Museum of Natural History, vol. 294, , p. 1–240 (DOI 10.1206/0003-0090(2005)294[0001:SROTFF]2.0.CO;2)
- (en) S. L. Kuzmin, « Pelophylax esculentus », sur amphibiaweb.org, (consulté le )
- (en) S. Köhler, « Mechanisms for partial reproductive isolation in a Bombina hybrid zone in Romania », sur Dissertation for thesis, (consulté le )
- Amphibian Species of the World, consulté le 17 septembre 2014
- (en) Diego San Mauro, Miguel Vences, Marina Alcobendas, Rafael Zardoya et Axel Meyer, « Initial Diversification of living amphibians predated the breakup of Pangaea », The American Naturalist, vol. 165, no 5, , p. 590–599 (PMID 15795855, DOI 10.1086/429523, JSTOR 429523)
- (en) Zhang Peng, Zhou Hui, Chen Yue-Qin, Liu Yi-Fei et Liang-Hu Qu, « Mitogenomic perspectives on the origin and phylogeny of living amphibians », Systematic Biology, vol. 54, no 3, , p. 391–400 (DOI 10.1080/10635150590945278)
- (en) David Marjanović et Michel Laurin, « Fossils, molecules, divergence times, and the origin of lissamphibians », Systematic Biology, vol. 56, no 3, , p. 369–388 (DOI 10.1080/10635150701397635)
- (en) R. Alexander Pyron, « Divergence time estimation using fossils as terminal taxa and the origins of Lissamphibia », Systematic Biology, vol. 60, no 4, , p. 466–481 (DOI 10.1093/sysbio/syr047)
- (en) Anne Casselman, « "Frog-amander" fossil may be amphibian missing link », National Geographic News, (consulté le )
- (en) Jason S. Anderson, Robert R. Reisz, Diane Scott, Nadia B. Fröbisch et Stuart S. Sumida, « A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders », Nature, vol. 453, no 7194, , p. 515–518 (PMID 18497824, DOI 10.1038/nature06865)
- (en) David Cannatella, « Triadobatrachus massinoti », sur Tree of Life, (consulté le )
- (en) Z. Roček, Amphibian Biology: Paleontology: The Evolutionary History of Amphibians, vol. 4, Surrey Beatty & Sons, (ISBN 0-949324-87-6, lire en ligne), chap. 14 (« Mesozoic Amphibians »), p. 1295–1331
- (en) D. B. Weishampel, P. Dodson (éd.) et H. Osmólska (éd.), Dinosaur distribution (Early Jurassic, North America): The Dinosauria (2nd edition), University of California Press, (ISBN 978-0-520-24209-8), p. 530–532
- (en) Neil H. Shubin et Farish A. Jenkins, « An Early Jurassic jumping frog », Nature, vol. 377, no 6544, , p. 49–52 (DOI 10.1038/377049a0)
- (en) J. Foster, Jurassic west : the dinosaurs of the Morrison Formation and their world, Indiana University Press, (ISBN 0-253-34870-6), « Anura (Frogs) », p. 135–136
- (en) A. C. Wilson, V. M. Sarich et L. R. Maxson, « The importance of gene rearrangement in evolution: evidence from studies on rates of chromosomal, protein, and anatomical evolution », Proceedings of the National Academy of Sciences, vol. 71, no 8, , p. 3028–3030 (DOI 10.1073/pnas.71.8.3028, lire en ligne)
- (en) S. E. Evans, M. E. H. Jones et D. W. Krause, « A giant frog with South American affinities from the Late Cretaceous of Madagascar », Proceedings of the National Academy of Sciences, vol. 105, no 8, , p. 2951–2956 (PMID 18287076, PMCID 2268566, DOI 10.1073/pnas.0707599105)
- Marjanović, D. & Laurin, M. (2007) Fossils, Molecules, Divergence Times, and the Origin of Lissamphibians . Systematic Biology 56, 369-388.
- (en) R. Alexander Pyron et John J. Wiens, « A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians », Molecular Phylogenetics and Evolution, vol. 61, no 2, , p. 543–583 (PMID 21723399, DOI 10.1016/j.ympev.2011.06.012, lire en ligne)
- (en) D. R. Frost, T. Grant, J. N. Faivovich, R. H. Bain, A. Haas, C. L. F. B. Haddad, R. O. De Sá, A. Channing, M. Wilkinson, S. C. Donnellan, C. J. Raxworthy, J. A. Campbell, B. L. Blotto, P. Moler, R. C. Drewes, R. A. Nussbaum, J. D. Lynch, D. M. Green et W. C. Wheeler, « The Amphibian Tree of Life », Bulletin of the American Museum of Natural History, vol. 297, , p. 1–291 (DOI 10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2)
- (en) Heinicke M. P., Duellman, W. E., Trueb, L., Means, D. B., MacCulloch, R. D. et Hedges, S. B., « A new frog family (Anura: Terrarana) from South America and an expanded direct-developing clade revealed by molecular phylogeny », Zootaxa, vol. 2211, , p. 1–35 (lire en ligne)
- (en) J. M. Hoekstra, J. L. Molnar, M. Jennings, C. Revenga, M. D. Spalding, T. M. Boucher, J. C. Robertson, T. J. Heibel et K. Ellison, « Number of Globally Threatened Amphibian Species by Freshwater Ecoregion », The Atlas of Global Conservation: Changes, Challenges, and Opportunities to Make a Difference., The Nature Conservancy, (consulté le )
- (en) S. N. Stuart, J. S. Chanson, N. A. Cox, B. E. Young, A. S. L. Rodrigues, D. L. Fischman et R. W. Waller, « Status and trends of amphibian declines and extinctions worldwide », Science, vol. 306, no 5702, , p. 1783–1786 (PMID 15486254, DOI 10.1126/science.1103538)
- (en) J. Alan Pounds, Michael P. L. Fogden, Jay M. Savage et George C. Gorman, « Tests of null models for amphibian declines on a tropical mountain », Conservation Biology, vol. 11, no 6, , p. 1307–1322 (DOI 10.1046/j.1523-1739.1997.95485.x)
- (en) « Worldwide Amphibian Declines: How big is the problem, what are the causes and what can be done? », AmphibiaWeb, (consulté le )
- (en) « Frog population decrease mostly due to traffic », New Scientist, (lire en ligne, consulté le )
- (en) M. J. Voordouw, D. Adama, B. Houston et P. Govindarajulu, « Prevalence of the pathogenic chytrid fungus, Batrachochytrium dendrobatidis, in an endangered population of northern leopard frogs, Rana Pipiens », BMC Ecology, vol. 10, no 6, (DOI 10.1186/1472-6785-10-6)
- (en) Elizabeth M. Harp et James W. Petranka, « Ranavirus in wood frogs (Rana sylvatica): Potential sources of transmission within and between ponds », Journal of Wildlife Diseases, vol. 42, no 2, , p. 307–318 (PMID 16870853)
- (en) Kathryn Phillips, Tracking the Vanishing Frogs, Penguin Books, (ISBN 0-14-024646-0)
- (en) Karen R. Lips, « Decline of a tropical montane amphibian fauna », Conservation Biology, vol. 12, , p. 106–117 (DOI 10.1111/j.1523-1739.1998.96359.x, JSTOR 2387466)
- (en) Andrew R. Blaustein et Pieter T. J. Johnson, « The complexity of deformed amphibians », Frontiers in Ecology and the Environment, vol. 1, no 2, , p. 87–94 (DOI 10.1890/1540-9295(2003)001[0087:TCODA]2.0.CO;2, lire en ligne)
- (en) J. G. Burkhart, G. Ankley, H. Bell, H. Carpenter, D. Fort, D. Gardiner, H. Gardner, R. Hale, J. C. Helgen, P. Jepson, D. Johnson, M. Lannoo, D. Lee, J. Lary, R. Levey, J. Magner, C. Meteyer, M. D. Shelby et G. Lucier, « Strategies for assessing the implications of malformed frogs for environmental health », Environmental Health Perspectives, vol. 108, no 1, , p. 83–90 (PMID 10620528, PMCID 1637865, DOI 10.2307/3454299, JSTOR 3454299)
- (en) Richard Black, « New frog centre for London Zoo », BBC News, (lire en ligne, consulté le )
- (en) « National recovery plan for the Southern Corroboree Frog (Pseudophryne corroboree): 5. Previous recovery actions », Environment.gov.au (consulté le )
- (en) « Bacteria show promise in fending off global amphibian killer », Physorg.com, (consulté le )
- (en) « Project launched to fight frog-killing fungus », The Guardian, 12 mai 12 mai 2009 (consulté le )
- (en) « Panama Amphibian Rescue and Conservation Project », sur amphibianrescue.org (consulté le )
- (en) « 2008: Year of the Frog », sur bgci.org, (consulté le )
- (en) Michael J. Tyler, Australian Frogs, Penguin Books, (ISBN 0-670-90123-7), p. 111
- (en) Elizabeth Cameron, « Cane toad », Australian Museum, (consulté le )
- (en) I. G. Warkentin, D. Bickford, N. S. Sodhi et J. A. Corey, « Eating frogs to extinction », Conservation Biology, vol. 23, no 4, , p. 1056–1059 (PMID 19210303, DOI 10.1111/j.1523-1739.2008.01165.x)
- (en) « Cultured Aquatic Species Information Programme: Rana catesbeiana », FAO: Fisheries and Aquaculture Department (consulté le )
- C. Mammina, L. Cannova, S. Carfì Pavia et A Nastasi, « Présence endémique de Salmonella enterica de sérotype Cerro au sud de l'Italie », Euro Surveill, vol. 5, no 7, , p. 84-86
- Cl. Richard, « A propos d'Aeromonas hydrophila », Medecine et Maladies Infectieuses, vol. 18, no 2, , p. 92
- (en) « California Schools Leading Race to Stop Dissections », Animal Welfare Institute, (lire en ligne, consulté le )
- (en) David Ames Wells, The science of common things: a familiar explanation of the first principles of physical science. For schools, families, and young students, (lire en ligne), p. 290
- (en) « Stannius ligature », Biology online, (consulté le )
- (en) S. Sarkar, « Lancelot Hogben, 1895–1975 », Genetics, vol. 142, no 3, , p. 655–660 (PMID 8849876, PMCID 1207007)
- (en) Christen Brownlee, « Nuclear Transfer: Bringing in the Clones », Proceedings of the National Academy of Sciences of the United States of America (consulté le )
- (en) L. Browder et L. Iten, « Xenopus as a Model System in Developmental Biology », Dynamic Development, Université de Calgary, (consulté le )
- (en) S. Klein, « Developing the potential of Xenopus tropicalis as a genetic model », Trans-NIH Xenopus Working Group (consulté le )
- (en) « Xenopus tropicalis », Joint Genome Institute (consulté le )
- (en) S. E. VanCompernolle, R. J. Taylor, K. Oswald-Richter, J. Jiang, B. E. Youree, J. H. Bowie, M. J. Tyler, M. Conlon, D. Wade, C. Aiken, T. S. Dermody, V. N. KewalRamani, L. A. Rollins-Smith et D. Unutmaz, « Antimicrobial peptides from amphibian skin potently inhibit human immunodeficiency virus infection and transfer of virus from dendritic cells to T cells », Journal of Virology, vol. 79, no 18, , p. 11598–11606 (PMID 16140737, PMCID 1212620, DOI 10.1128/JVI.79.18.11598-11606.2005)
- (en) G. Phillipe et L. Angenot, « Recent developments in the field of arrow and dart poisons », Journal of Ethnopharmacology, vol. 100, nos 1–2, , p. 85–91 (PMID 15993556, DOI 10.1016/j.jep.2005.05.022)
- (en) T. Lyttle, D. Goldstein et J. Gartz, « Bufo toads and bufotenine: fact and fiction surrounding an alleged psychedelic », Journal of Psychoactive Drugs, vol. 28, no 3, , p. 267–290 (PMID 8895112, DOI 10.1080/02791072.1996.10472488)
- (en) T. Lyttle, « Misuse and legend in the "toad licking" phenomenon », International Journal of Addiction, vol. 28, no 6, , p. 521–538 (PMID 8486435)
- (en) Charles W. Myers, John W. Daly et Borys Malkin, « A dangerously toxic new frog (Phyllobates) used by Emberá Indians of western Colombia, with discussion of blowgun fabrication and dart poisoning », Bulletin of the American Museum of Natural History, vol. 161, , p. 307–366 (lire en ligne)
- (en) Roger Ebert, « Chuck Jones: Three Cartoons (1953–1957) », Chicago Sun-Times, (lire en ligne, consulté le )
- (en) Brothers Grimm, « The Frog Prince », East of the Web (consulté le )
- (en) « Walk of Fame, Kermit the Frog awarded with a star on the Hollywood Walk of Fame, December 1, 2002 », Walkoffame.com (consulté le )
- (en) William E. Burns, Witch Hunts in Europe and America: An Encyclopedia, Greenwood Publishing Group, (ISBN 0-313-32142-6, lire en ligne), p. 7
- (en) Katherine Berrin et Larco Museum, The Spirit of Ancient Peru: Treasures from the Museo Arqueológico Rafael Larco Herrera, New York, Thames and Hudson, (ISBN 0-500-01802-2)
- (en) B. Gratwicke, « The Panamanian Golden Frog », sur Panama Amphibian Rescue and Conservation Project,