Permien
Le Permien est un système géologique qui a duré de 298,9 ± 0,2 à 252,2 ± 0,5 millions d'années. C'est la dernière période du Paléozoïque, précédée par le Carbonifère et suivie par le Trias qui est le premier système du Mésozoïque. Le Permien a été nommé d'après la province russe de Perm où se situent des gisements fossilifères de cette période. La fin du Permien est marquée par la troisième des cinq principales extinctions de masse survenues sur Terre : c'est la plus sévère, qui, selon les estimations des scientifiques, a vu disparaître 70 % des espèces de la terre ferme[5] et 96 % des espèces marines[6].
Dyas
| Notation chronostratigraphique | P |
|---|---|
| Notation française | r |
| Notation RGF | r |
| Stratotype initial |
|
| Niveau | Période / Système |
| Érathème / Ère - Éonothème / Éon |
Paléozoïque Phanérozoïque |
Stratigraphie
| Début | Fin |
|---|---|
| Fossiles stratigraphiques |
fusulinidés ammonites |
|---|
- P. supérieur (Lopingien, ≃Thuringien)
- P. moyen (Guadalupien, ≃Saxonien)
- P. inférieur (Cisuralien, ≃Autunien)
Dyas : Zechstein et Rotliegend
Paléogéographie et climat
| Taux de O2 atmosphérique |
env. 23 %vol[1] (115 % de l'actuel) |
|---|---|
| Taux de CO2 atmosphérique |
env. 900 ppm[2] (3 fois le niveau d'avant la révolution industrielle) |
| Température moyenne |
16 °C[3] (+2 °C par rapport à l'actuel) |
| Niveau moyen des mers | +60 (début) à −20 (fin) m (par rapport à l'actuel)[4] |
- Trapps de Sibérie
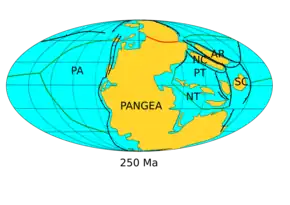
- 280 Ma : formation de la Pangée
Faune et flore
- bipédie
- vol
- disparition des trilobites
- 290 Ma : cycadales (plantes en forme de palmiers)
- 252 Ma : archosauriens (≃reptiles)
| Affleurements notables |
|
|---|
Subdivisions
Comme pour toutes les périodes géologiques anciennes, les couches stratigraphiques de référence sont bien connues, mais leur datation exacte est sujette à des variations de quelques millions d'années suivant les mesures isotopiques et les auteurs. Les datations des subdivisions correspondent à celles de l'échelle des temps géologiques publiée en 2012 (Geologic Time Scale 2012, GTS2012)[7] - [8].
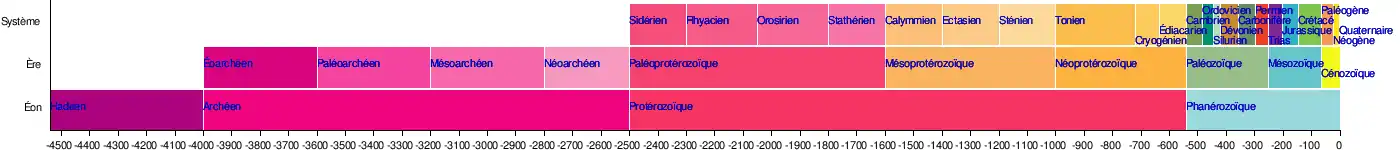
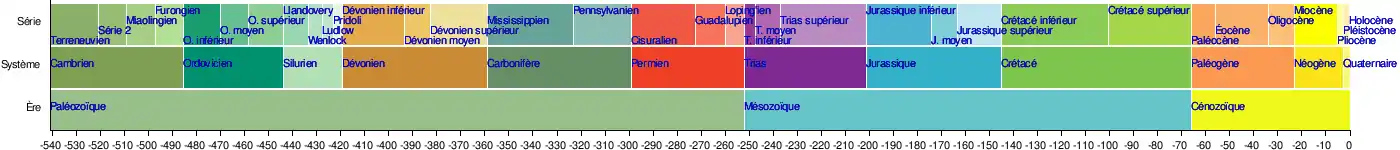
La Commission internationale de stratigraphie divise le Permien en trois séries (ou époques) : le Cisuralien (298,9 ± 0,2 à 272,3 ± 0,5 Ma), le Guadalupien (272,3 ± 0,5 à 259,9 ± 0,4 Ma) et le Lopingien (259,9 ± 0,4 à 252,2 ± 0,5 Ma)[7].
- Cisouralien (Cisuralian en anglais) :
- Assélien (298,9 ± 0,2 - 295,5 ± 0,4 Ma)
- Sakmarien (295,5 ± 0,4 - 290,1 ± 0,1 Ma)
- Artinskien (290,1 ± 0,1 - 279,3 ± 0,6 Ma)
- Koungourien (279,3 ± 0,6 - 272,3 ± 0,5 Ma)
- Guadalupien :
- Roadien (272,3 ± 0,5 - 268,8 ± 0,5 Ma)
- Wordien (268,8 ± 0,5 - 265,1 ± 0,4 Ma)
- Capitanien (265,1 ± 0,4 - 259,9 ± 0,4 Ma)
- Lopingien :
- Wuchiapingien (259,9 ± 0,4 - 254,2 ± 0,1 Ma)
- Changhsingien (254,2 ± 0,1 - 252,2 ± 0,5 Ma)
Particularités locales
En Europe du Nord et centrale, et particulièrement dans la littérature germanique et ancienne, on trouve une division du Permien différente, en deux parties, avec des séries du Rotliegend (-302 Ma à -258 Ma) suivies de séries du Zechstein (-258 Ma à -251 Ma). Lorsque l'on fait référence à ces deux divisions du Permien, on parle alors non plus de Permien, mais de Dyas (par analogie avec le Trias qui lui, comporte trois divisions) et ce Dyas s'étend alors de -302 Ma à -251 Ma.
Paléogéographie et climat
Le niveau moyen de la mer est resté assez bas durant le Permien. Toutes les masses continentales, à l'exception d'une portion de l'Asie du Sud-Est, étaient agglomérées en un seul supercontinent appelé Pangée, qui s'étendait de l'équateur aux pôles, entouré par un océan nommé Panthalassa (la « mer universelle »). Cette période géologique voit le début du processus de rifting qui entraîne le morcellement de la Pangée. Se développe ainsi la Téthys, un paléo-océan qui s'ouvre progressivement à partir du Permien supérieur d'est en ouest à travers la Pangée[9].
Ce grand continent crée des conditions climatiques impliquant de grandes variations de température et de précipitations (donc d'érosion) selon les saisons et les moments de la journée. En son centre, la température peut passer de 0 °C à 40 °C dans la journée, ce qui fait apparaître, chez diverses espèces originaires de lignées différentes, des « voiles » thermorégulatrices, permettant de capter le soleil au matin, et d'évacuer la chaleur dans la journée[10]. Dans les régions polaires australes, correspondant à l'actuel bassin du Congo, perdure un inlandsis présent depuis le Carbonifère, mais le reste de la Pangée connait des conditions climatiques arides avec des températures élevées et de faibles précipitations[10].
Faune et flore
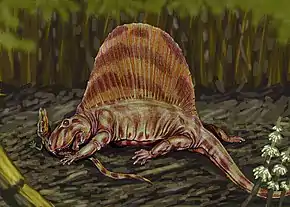
La faune a connu quelques évolutions intéressantes à cette période : on y note notamment l'apparition de la bipédie temporaire avec Aphelosaurus dès le Permien inférieur et la bipédie très probablement permanente avec Eudibamus à la même époque[11]. Des animaux vertébrés volants font leur apparition avec Coelurosauravus au Permien supérieur (vol planant et non battu)[12] - [13]. Les synapsides sont les premiers amniotes ayant prospéré et qui se sont considérablement diversifiés lors de cette époque. Les synapsides du Permien comprenaient quelques membres importants tels que Dimetrodon et Edaphosaurus. Leurs adaptations spéciales les ont permis de s'épanouir dans le climat plus sec du Permien et ayant atteint des tailles considérables pour dominer les autres tétrapodes[10]. Durant le début du Guadalupien, un nouveau groupe de synapsides, les thérapsides, vont dominer l'immense majorité des faunes du Permien et du Trias.
Les formes de vie dominantes sont diverses : plantes comme les fougères qui dispersent des spores, grands amphibiens principalement temnospondyles, premiers reptiles, thérocéphales (« reptiles mammaliens ») incluant les ancêtres des futurs mammifères et archosaures incluant les ancêtres des futurs dinosaures. La vie marine est riche en mollusques, échinodermes et brachiopodes. Les derniers trilobites ont disparu avant la fin du Permien. Les conditions plus sèches ont favorisé les gymnospermes comme les « fougères à graines » telles les glossoptérides, et les premiers conifères (arbres modernes), apparus durant le Permien. C'est aussi du Permien que datent les fossiles de la plus grande « libellule » jamais identifiée, Meganeuropsis, qui dépasse de peu la fameuse Meganeura du Carbonifère de la taille d'un chat.
Les coquilles fossilisées d'espèces non-vertébrées sont souvent utilisées pour identifier les strates géologiques du Permien : les fusulinidés, foraminifères benthiques qui disparaissent quasi totalement à la limite entre le Permien moyen et supérieur, et les ammonites (principalement goniatites) dont l'équivalent moderne est le Nautile. D'autres marqueurs stratigraphiques pour le Permien sont les dents de conodontes, organismes marins d'aspect vermiforme, disparus à la fin du Trias (peut-être apparentés aux vertébrés, bien que cette hypothèse soit discutée[14] - [15]).
 Reconstitution par Dimitri Bogdanov d’Edaphosaurus, un pélycosaure herbivore et, à ses pieds, Platyhystrix, un amphibien, sont pourvus de voiles thermorégulatrices, adaptations aux contrastes thermiques journaliers.
Reconstitution par Dimitri Bogdanov d’Edaphosaurus, un pélycosaure herbivore et, à ses pieds, Platyhystrix, un amphibien, sont pourvus de voiles thermorégulatrices, adaptations aux contrastes thermiques journaliers. Reconstitution par D. Bogdanov de Dimetrodon et d’Eryops.
Reconstitution par D. Bogdanov de Dimetrodon et d’Eryops. Reconstitution par D. Bogdanov d'un groupe d'Estemmenosuchus uralensis en compagnie d'un Eotitanosuchus.
Reconstitution par D. Bogdanov d'un groupe d'Estemmenosuchus uralensis en compagnie d'un Eotitanosuchus. Reconstitution par D. Bogdanov d'un Titanophoneus dévorant un Ulemosaurus.
Reconstitution par D. Bogdanov d'un Titanophoneus dévorant un Ulemosaurus. Reconstitution par D. Bogdanov d’un Inostrancevia alexandri attaquant un Scutosaurus juvénile.
Reconstitution par D. Bogdanov d’un Inostrancevia alexandri attaquant un Scutosaurus juvénile.
Les extinctions

Selon les auteurs, on reconnaît au moins trois[16], peut-être quatre[17] crises pendant le Permien, dont la dernière, tout à la fin du Permien, est de loin la mieux connue et probablement la plus sévère. La plus ancienne peut avoir eu lieu autour de la limite Sakmarien/Artinskien ou autour de la limite Artinskien/Kungurien. Elle peut même être avoir consisté en un lent déclin de la biodiversité pendant le Sakmarien et l'Artinskien, mais une étude récente sur des synapsides de ces étages n'a pas pu la mettre en évidence[16]. La seconde crise, qui semble avoir eu un impact plus important sur les synapsides, a eu lieu autour de la limite Kungurian/Roadien, qui est aussi la limite Cisuralien/Guadaloupien, mais cette crise semble avoir consisté en un déclin progressif de la biodiversité s'étendant sur quelques millions d'années. La troisième a eu lieu vers la fin du Guadaloupien[18] et semble avoir été assez sévère chez les vertébrés continentaux[17]. Elle a entre autres éliminé les dinocéphales, un groupe de thérapsides qui était assez abondant pendant le Wordien et le Capitanien[19]. Le dernier varanopidé connu date du Capitanien[20], mais ce groupe (ainsi que les autres synapsides basaux) était peut-être éteint avant la crise de la fin de cet étage.
Le système Permien se termine vers -251,4 millions d'années par la plus grande extinction d'espèces connue, couramment nommée « crise du Permo-Trias ». Selon les estimations scientifiques, 75 % des espèces terrestres et 96 % des espèces marines disparaissent. Parmi les espèces animales et végétales qui disparaissent, citons les trilobites, les graptolites, certaines fougères, certains coraux ou encore de nombreux groupes de tétrapodes, dont de nombreux dicynodontes, les gorgonopsiens, pareiasaures et de nombreux temnospondyles. De toutes les lignées d'Ammonites fort diversifiées depuis le Dévonien, seuls deux genres survivent : les Prolecanitida et les Ceratitida, à partir desquels le groupe se diversifiera à nouveau au Mésozoïque.
Les causes de cette extinction massive sont toujours discutées entre scientifiques. Les plus souvent citées sont une asphyxie des océans (ou anoxie), un volcanisme majeur en Sibérie, une baisse importante du niveau de la mer (ou régression) ou une combinaison de plusieurs causes.
Les indices d'un impact météoritique datant de cette époque ont récemment été découverts en Antarctique (2006), dans la Terre de Wilkes : on évalue le diamètre de cette météorite à 48 km. D'autres sites probables d'impacts météoriques ont été relevés, comme le cratère de Bedout situé au large des côtes australiennes. Cependant certains géologues avancent que Bedout est l'empreinte d'un phénomène volcanique[21].
Quelles qu'en soient les causes initiales, la plupart des indicateurs paléontologiques et géochimiques convergent à attribuer l'extinction massive à une dégradation très marquée de l'environnement[22].
Affleurements du Permien
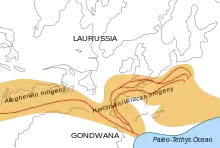
Au Permien, la tectogenèse hercynienne principale est achevée et la chaîne hercynienne est progressivement réduite par l'érosion, formant une plate-forme continentale, tandis que les sédiments détritiques issus du démantèlement de ces montagnes s'accumulent dans des bassins sédimentaires[23].
Plusieurs zones sont particulièrement riches en affleurements de dépôts datant du Permien.
Pour les sédiments marins :
- les versants occidentaux de l'Oural, près de Perm en Russie ;
- la Chine ;
- l'État du Texas aux États-Unis où le bassin permien présente l'un des dépôts les plus épais connus, particulièrement dans les Guadalupe Mountains (qui ont donné leur nom au Guadalupien) ;
- les chaînes de montagnes situées entre les Dolomites italiennes et l'Himalaya : Dolomites, Dinarides, Hellénides, Monts Taurus, Zagros, Oman, Himalaya senso lato.
Pour les sédiments continentaux :
- les montagnes de l'Oural, près de Perm ;
- les régions d'Arlit et d'Agadez au Niger, où deux nouveaux fossiles d'amphibiens (Nigerpeton ricqlesi et Saharastega moradiensis) ont été trouvés en 2003 dans la formation de Moradi, laissant penser qu'il y avait dans cette zone des formes de vies différentes de celles d'Afrique australe et de Russie[24] - [25] ;
- les bassins du Karoo (Afrique du Sud) et de Bowen (Australie).
Notes et références
- (de) teneur en oxygène dans l'atmosphère au Phanérozoïque
- (en) dioxyde de carbone au Phanérozoïque
- (en) température de la Terre
- (fr) variation du niveau des mers au Phanérozoïque
- David Burnie, L'encyclopédie des dinosaures, Rouge & Or, Chine, 2005, p. 56
- Sébastien Steyer, La Terre avant les dinosaures, Belin, Paris, 2009, p. 159
- Gradstein et al. 2012.
- « Charte stratigraphique internationale (2012) » [PDF], sur http://www.stratigraphy.org/ (consulté le ).
- (en) I. Metcalfe, « Gondwana dispersion and Asian accretion: tectonic and palaeogeographic evolution of eastern Tethys », Journal of Asian Earth Sciences, vol. 66, , p. 2 (DOI 10.1016/j.jseaes.2012.12.020).
- David Burnie, L'encyclopédie des dinosaures, op. cit., p. 52
- Sébastien Steyer, La Terre avant les dinosaures, op. cit., p. 147 et 150
- Sébastien Steyer, La Terre avant les dinosaures, op. cit., p. 151
- (en) Valentin Buffa, Eberhard Frey, J.-Sébastien Steyer et Michel Laurin, « A new cranial reconstruction of Coelurosauravus elivensis Piveteau, 1926 (Diapsida, Weigeltisauridae) and its implications on the paleoecology of the first gliding vertebrates », Journal of Vertebrate Paleontology, vol. 41, no 2, , e1930020 (ISSN 0272-4634 et 1937-2809, DOI 10.1080/02724634.2021.1930020, lire en ligne, consulté le )
- Alain Blieck, Susan Turner, Carole J. Burrow et Hans-Peter Schultze, « Fossils, histology, and phylogeny: Why conodonts are not vertebrates », Episodes, vol. 33, no 4, , p. 234–241 (ISSN 0705-3797 et 2586-1298, DOI 10.18814/epiiugs/2010/v33i4/002, lire en ligne, consulté le )
- Susan Turner, Carole J. Burrow, Hans-Peter Schultze et Alain Blieck, « False teeth: conodont-vertebrate phylogenetic relationships revisited », Geodiversitas, vol. 32, no 4, , p. 545–594 (ISSN 1280-9659 et 1638-9395, DOI 10.5252/g2010n4a1, lire en ligne, consulté le )
- (en) Gilles Didier et Michel Laurin, « Distributions of extinction times from fossil ages and tree topologies: the example of mid-Permian synapsid extinctions », PeerJ, vol. 9, , e12577 (ISSN 2167-8359, PMID 34966586, PMCID PMC8667717, DOI 10.7717/peerj.12577, lire en ligne, consulté le )
- (en) S.G. Lucas, « Permian tetrapod extinction events », Earth-Science Reviews, vol. 170, , p. 31–60 (DOI 10.1016/j.earscirev.2017.04.008, lire en ligne, consulté le )
- (en) Michael O. Day, Jahandar Ramezani, Samuel A. Bowring et Peter M. Sadler, « When and how did the terrestrial mid-Permian mass extinction occur? Evidence from the tetrapod record of the Karoo Basin, South Africa », Proceedings of the Royal Society B: Biological Sciences, vol. 282, no 1811, , p. 20150834 (ISSN 0962-8452 et 1471-2954, PMID 26156768, PMCID PMC4528552, DOI 10.1098/rspb.2015.0834, lire en ligne, consulté le )
- (en) Michael, Saniye Güven, Fernando Abdala, S. Jirah, B. Rubidge et J. Almond, « Youngest dinocephalian fossils extend the Tapinocephalus Zone, Karoo Basin, South Africa », South African Journal of Science, vol. 111, nos 3/4, (DOI 10.17159/sajs.2015/20140309, lire en ligne, consulté le )
- (en) Sean P. Modesto, Roger M. H. Smith, Nicolás E. Campione et Robert R. Reisz, « The last “pelycosaur”: a varanopid synapsid from the Pristerognathus Assemblage Zone, Middle Permian of South Africa », Naturwissenschaften, vol. 98, no 12, , p. 1027–1034 (ISSN 0028-1042 et 1432-1904, DOI 10.1007/s00114-011-0856-2, lire en ligne, consulté le )
- Sébastien Steyer, La Terre avant les dinosaures, op. cit., p. 162
- Global and Platenary Change, Volume 55, Issues 1-3, p. 1-236 (January 2007) Environmental and Biotic Changes during the Paleozoic-Mesozoic Transition, Elsevier Publication, Amsterdam
- Claude Lorenz, Géologie des pays européens, Dunod, , p. 24.
- (en) C. A. Sidor, F. R. O'Keefe, R. Damiani, J. S. Steyer, R. M. H. Smith, H. C. E. Larsson, P. C. Sereno, O. Ide et A. Maga, « Permian tetrapods from the Sahara show climate-controlled endemism in Pangaea », Nature, vol. 434, no 7035, , p. 886-889 (DOI 10.1038/nature03393).
- Jean-Sébastien Steyer, « Le Permien au Sahara », sur http://www.larecherche.fr, (consulté le ).
Voir aussi
Bibliographie
- (en) F.M. Gradstein, J.G Ogg, M. Schmitz et G. Ogg, The Geologic Time Scale 2012, Elsevier, , 1176 p. (ISBN 978-0-444-59448-8, lire en ligne).
- (en) Felix M. Gradstein, James G. Ogg et Alan G. Smith, A Geologic Time Scale 2004, Cambridge, Cambridge University Press, , 610 p. (ISBN 0-521-78142-6)
Articles connexes
Liens externes
- (en) « International Commission on Stratigraphy. Subcommission on Permian Stratigraphy », sur http://www.stratigraphy.org/.
- (en) University of California avec une stratigraphie plus moderne du permien
- (en) Université de Berkeley avec une stratigraphie plus classique