Glossopteris (genre fossile)
Glossopteris est un genre de plantes de l'ordre éteint des Glossopteridales. C'est le genre le mieux connu de cet ordre. Ce dernier (parfois appelé Arberiales ou Dictyopteridales) appartient au groupe (sans doute une division) éteint des Pteridospermatophytes.

| Règne | Plantae |
|---|---|
| Division | † Pteridospermatophyta |
| Ordre | † Glossopteridales |
| Famille | † Glossopteridaceae |
Les Glossopteridales étaient abondantes au Permien. Elles sont considérées comme un marqueur stratigraphique du Permien bien que des spécimens d'âge Trias aient été rapportés et qu'il existe une occurrence anormale au Jurassique[1].
Distribution : Inde, Australie, Amérique du Sud, Antarctique, Afrique.
Étymologie
- « Glossopteris » vient du grec glossa, qui signifie « langue », en raison de la forme des feuilles.
Historique
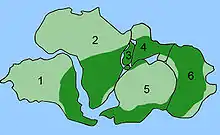
1 Amérique du Sud, 2 Afrique, 3 Madagascar, 4 Sous-continent indien, 5 Antarctique, 6 Australie.
L'ordre Glossopteris a été décrit en détail pour la première fois par le botaniste Adolphe Brongniart en 1828 (Prodrome d'une histoire des végétaux fossiles). Les glossoptérides et leurs différentes espèces avaient d'abord été classés tantôt dans différents groupes de gymnospermes : cycadophytes, fougères à graines, gnétophytes, Cordaitales, tantôt comme angiospermes, lorsque la découverte de leurs organes mégasporophylles[2], en 1977, confirma qu'il s'agissait bien d'un ordre disparu apparenté aux fougères à graines.
C'est la répartition des fossiles de Glossopteris qui a conduit le géologue Eduard Suess à envisager qu'il y avait eu par le passé une connexion entre Inde, Australie, Afrique, Amérique du Sud et Antarctique[3]. Il appela ce « super-continent » Gondwana d'après une région d'Inde (Gondwâna) où Glossopteris est abondant. Cette répartition de Glossopteris viendra soutenir la théorie d'Alfred Wegener sur la dérive des continents.
Description
Organes végétatifs

On suppose que glossopteris était une plante ligneuse qui avait un port de buisson ou d'arbre. Les plus grands spécimens ont pu atteindre 30 m de haut. Le tissu xylémique des glossoptérides est du pycnoxyle (xylème épais au parenchyme mince) présentant des cernes de croissance marqués. Les rayons sont unisériés, les parois radiales des trachéides portent des ponctuations araucarioïdes (on les range parfois dans le genre Araucarioxylon). Les parois primaires des trachéides s'organisent selon des lignes radiales, plusieurs vacuoles sont hexagonales. Les canaux de xylème sont monoxyles. Le tronc de Glossopteris skaarensis présente une paroi primaire dont les trachéides possèdent des stomates ronds.
Les organes souterrains ne plongent pas très profondément et sont classés comme Vertebraria. Au centre il y a un xylème primaire directeur, entouré de quatre à sept rayons de bois secondaire, séparés les uns des autres par des vacuoles. Le xylème secondaire présente des cernes de croissance. Le bois secondaire est entouré d'un périderme mince et spongieux. Le bois primaire est formé de trachéides annulaires, le bois secondaire de trachéides à stomates.
Les feuilles sont simples, alternes ou distribuées en hélices compactes autour des branches. Plusieurs indices donnent à penser que les feuilles se rattachaient sur les rameaux par petits groupes, comme c'est le cas pour le gingko. Les glossoptérides perdaient leurs feuilles à l'automne. La plupart des feuilles ont été retrouvées isolées.
Glossopteris est à ce jour le genre de végétaux le plus représenté dans les fossiles de cette période. Ses feuilles lancéolées présentent une nervure centrale bien nette, avec un réseau de nervures secondaires. On en a décrit plus de 200 variétés, vraisemblablement en raison de l'absence d'une classification standardisée. Par exemple 70 espèces ont été décrites rien qu'en Inde[4]. La plus commune est Glossopteris browniana, dont les feuilles mesurent plus de 30 cm et présentent une extrémité arrondie. L'épiderme de Glossopteris fibrosa a pu être étudiée : les fentes se limitent à la face postérieure des feuilles et sont entourées de quatre à huit cellules sœurs. Plusieurs variétés présentent des barbes sur la face postérieure des feuilles.
L'anatomie des feuilles n'a pu être étudiée que sur quelques espèces. Glossopteris schopfii possède de quatre à cinq faisceaux conducteurs le long de la nervure centrale. Les fascicules sont divisés par d'épaisses cellules fibreuses. La densité des tubes criblés, qu'on ne trouve également que sur la face postérieure, atteint 40 par mm². La nervure centrale de Glossopteris skaarensis est formée d'un large faisceau conducteur unique, à trachéides directeurs, entouré d'une mince membrane séparatrice. On trouve aussi des pétioles sur les formes apparues au Trias.
Gangamopteris est une espèce surtout représentée au Permien inférieur, dont les feuilles n'ont pas de nervure centrale perceptible. Les autres types de feuilles bien identifiées sont celles de Belemnopteris et de Rhabdotaenia ; on a trouvé plus rarement Rubidgea, Palaeovittaria et Euryphyllum.
Organes reproducteurs
Les organes reproducteurs (ovules et étamines) sont disposés sur les feuilles. Les organes mâles et femelles se trouvent sur des feuilles différentes.
Les organes porteurs de graines
Les organes femelles consistent en une structure dorso-ventrale, sur laquelle les graines se développent. On a donné divers noms à cette structure : capitulum, mégasporophylle, cupule, fertiliseur. Plusieurs de ces organes sont susceptibles d'avoir porté des cônes.
La morphologie des organes porteurs de graines est beaucoup plus variée que celle des organes végétatifs. On trouve ainsi :
- le scutum, support de graines en forme d'écu (comme son nom le suggère). La partie centrale, où se trouvent les rangées de graines, est bordées d'ailettes. Ottokaria présente une tige fine dont la tête comporte une sorte d'aigrette ou plumet, au niveau des graines de la partie centrale. On ignore si la tige était attachée à la nervure centrale ou à l'attache de la feuille. À chaque extrémité des pétioles de Glossopteris conspicua, la grappe Austroglossa walkomii comportait jusqu'à 20 graines.
- Lidgeottia est une grappe de quatre à huit structures (mégasporophylles) attachée à une feuille. Chaque mégasporophylle est aplati, d'un diamètre de 7 mm. Le bord est découpé à dentelé. Les graines de l'espèce Samaropsis ont un diamètre de 2 mm et comportent des ailettes pour le transport éolien.
- Chez Rusangea d’Afrique du Sud, chaque feuille possède deux pétioles, qui ne portent chacun qu'une seule graine. Rusangea elegans possède des feuilles longues de 2,8 cm et des graines ovales, sans ailettes, de 3,5 mm de diamètre.
- Denkania comporte six cupules à graine, que l'on trouve à l'extrémité d'un long pétiole sur chaque feuille de Glossopteris. D'un diamètre de 10 mm, elles ne contiennent qu'une graine chacune.
- On a trouvé des mégasporophylles enroulés de Glossopteris dans les couches de la fin du Permien au Queensland. Les rangées de graines se logent dans l'interstice de 1,5 mm ainsi formé. Cet interstice est rempli de filaments qui jouent sans doute le rôle d'aileron de dérive dans le transport des graines.
Les types de graine les plus courants sont le pterygosperme et les stéphanostomes.
Les organes producteurs de pollen
Les organes portant le pollen sont toujours plus difficiles à trouver dans les fossiles que les organes portant les graines. L'organe le plus souvent retrouvé s'appelle Glossotheca : c'est le prolongement du pétiole d'une feuille. Son extrémité est couverte de groupes d'étamines allongéeset peut comporter jusqu'à 100 étamines.
Les étamines isolées des glossopterides sont appelées Arberiella, Lithangium et Polytheca ; leurs sporanges sont uniloculaires. Les grains de pollen d’Arberiella possèdent deux poches d'air. Chez Arberiella vulgaris, ils sont longs de 85 µm. Chez Lithangium ils n'ont qu'un seul stigmate, et chez Polytheca, ils sont monocolpés (c'est-à-dire qu'ils ne comportent qu'un seul sillon).
Écologie
Les glossoptérides colonisaient, selon toute vraisemblance, les zones marécageuses : on le déduit de la présence de sacs remplis d'air le long de leur rhizome. Ils étaient adaptés à un cycle de saisons très différenciées, mais ont su s'étendre sur toutes les terres comprises entre 40 et 90° de latitude sud. Les feuilles fossilisées en épaisses couches donnent à penser que ces plantes étaient à feuilles caduques. Les cernes du bois sont des indices de croissance saisonnière[3].
Répartition
Les glossoptérides se sont répandus sur toute l'étendue du Gondwana et on trouve aujourd'hui leurs fossiles essentiellement en Australie, en Afrique, en Amérique du Sud et dans l’Antarctique. Ils ont constitué l'essentiel de la flore de ces régions au cours du Paléozoïque tardif.
Systématique
La classification génétique des glossoptérides demeure un mystère. Ces fougères partagent les caractéristiques morphologiques de différents groupes, mais celui dont elles sont, somme toute, les plus proches, est celle des fougères à graines, parmi lesquelles on les range pour l'instant.
Schopf considérait les glossoptérides comme l’ancêtre des gnetales[5]. Les recherches de cladistique de Doyle et Donoghue les rangent comme parentes des angiospermes[6].
Notes et références
- (en)Delevoryas, T., Person, C.P., 1975. Mexiglossa varia gen.et sp.nov., a new genus of glossopteroid leaves from the Jurassic of Oaxaca, Mexico.Palaeontographica 154B, 114-120
- D'après R.E. Gould, T. Delevoryas, « The biology of Glossopteris: Evidence from petrified seed-bearing and pollen-bearing organs », Alcheringia Archives, vol. 1, , p. 387-399.
- Cf. (en) R.S. Hill, E.M. Truswell, S. McLaughlin, M.E. Dettmann et A. E. Orchard (dir.), Flora of Australia. Volume 1., Introduction, Melbourne, ABRS/CSIRO Australia, (réimpr. 2), 670 p. (ISBN 0-643-05965-2), « Evolution of the Australian Flora: Fossil evidence », p. 260
- (en) Chandra et Suranges, 1975
- Cf. J. M. Schopf, « Morphologic interpretation of fertile structures in glossopterid gymnosperms », Review of paleobotany and palynology, no 21, , p. 25-64
- Cf. J.A. Doyle, M.J. Donoghue et B.A. Thomas et R.A. Spicer (dir.), Systematic and Taxonomic Approaches in Paleobotany, Londres, Oxford Univ. Press, , « Relationships of angiosperms and Gnetales: a numerical cladistic analysis »
Voir également
Liens externes
- Glossipteridales
- Ressources relatives au vivant :
- Tela Botanica
- (en) Paleobiology Database
- (en) Germplasm Resources Information Network
- (mul + en) iNaturalist
- (en) International Plant Names Index
- (en) Plants of the World Online
Bibliographie
- (en) Thomas Taylor et Edith Taylor, The Biology and Evolution of Fossil Plants, Englewood Cliffs, Prentice Hall, , 982 p. (ISBN 0-13-651589-4), p. 558–574