Changement de couleur des feuilles
Durant les mois d'automne, des arbres feuillus et des fougères connaissent un changement de couleur des feuilles et des frondes avant qu'elles ne tombent. Ce changement de couleurs, qui existe chez beaucoup d'arbres des régions tempérées[2], est souvent appelé couleurs automnales. Il est associé au dernier stade du développement de la feuille, la sénescence qui se caractérise par de véritables bouleversements biochimiques.


Chlorophylle et couleur verte
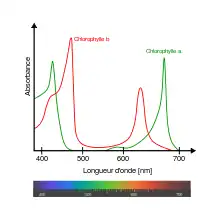


Une feuille est verte à cause de la présence d'un groupe de pigments appelé chlorophylle. Quand la chlorophylle est abondante dans les cellules, comme c'est le cas pendant la période de croissance des végétaux, le vert des molécules de chlorophylle domine et masque la couleur de tout autre pigment qui pourrait être présent dans la feuille. Ainsi, en été les feuilles sont généralement vertes.
La chlorophylle est un élément essentiel de la biochimie végétale par son rôle dans la photosynthèse qui consiste à absorber les rayons solaires et utiliser l'énergie résultant de cette absorption dans l'élaboration de la “nourriture” de la plante - les sucres simples, produits à partir d'eau et de dioxyde de carbone. Ces sucres sont la base du métabolisme de la plante et sont donc indispensables pour sa croissance et son développement. Comme toute molécule, la chlorophylle a une durée de vie, et elle est donc continuellement détruite. Cependant, durant la période de croissance, la perte des molécules de chlorophylle est compensée par une synthèse équivalente, ainsi le niveau de chlorophylle reste-t-il élevé et les feuilles restent vertes. Les végétaux investissent jusqu'à un tiers de leur photosynthèse nette dans la synthèse du système de captage du rayonnement[4].
À la fin de l'été, en réponse à des signaux environnementaux tels que le froid et la diminution de la durée du jour, les nervures qui assurent le transport de fluides vers l'intérieur et l'extérieur de la feuille sont progressivement obstruées par un bouchon de liège à la base des feuilles, à l'endroit où précisément le pétiole se brisera (phénomène d'abscission). Avec le développement de cette couche de liège, l'afflux d'eau et de minéraux se réduit de plus en plus rapidement. C'est durant cette période que le niveau de chlorophylle commence à décroître : le processus très actif de la sénescence implique le recyclage des nutriments essentiels pour la plante conduisant notamment à un turn-over protéique supérieur à 60 % (40 % restant dans la feuille)[5]. Or une proportion importante de chlorophylle est associée aux protéines membranaires des thylakoïdes situés dans les chloroplastes. Avant que le bouchon de liège n'ait complètement obstrué le pétiole, la plante récupère les protéines chloroplastiques en séparant le complexe protéine-pigment[5]. Cette réallocation des ressources au reste de la plante, dans la perspective de leur réutilisation au printemps, s'accompagne du changement de couleur des feuilles, de leur chute[6], et de leur recyclage au sol où les constituants foliaires sont décomposés par les microorganismes du sol[7].
Pigments qui contribuent aux autres couleurs
Caroténoïdes
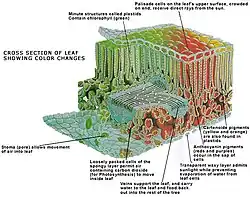
Quand l'automne approche, certains facteurs à la fois internes et externes à la plante provoquent un ralentissement du renouvellement chlorophyllien, la vitesse de renouvellement devient alors inférieure à celle de la dissociation de la chlorophylle. Pendant cette période, la synthèse en chlorophylle diminuant graduellement, l'effet de masque s'atténue lentement. C'est alors que les autres pigments, qui étaient présents dans les cellules durant toute la vie de la feuille, commencent à être visibles. Ce sont surtout les caroténoïdes, qui émettent dans des couleurs jaunes, orange, et brunes, et dans une moindre mesure les tanins colorés (bruns, orangés diffus).
Les caroténoïdes se trouvent - au côté des pigments chlorophylliens - dans des organites appelées plastes, des structures minuscules à l'intérieur des cellules des feuilles. Ceux-ci sont parfois si abondants dans la feuille qu'ils donnent à la plante une couleur vert-jaune, même en été. Mais dans la plupart des cas, les caroténoïdes deviennent apparents pour la première fois en automne, quand les feuilles commencent à perdre leur chlorophylle.
Leur jaune et leur orange teintent les feuilles des espèces d'arbres à bois dur tel que le frêne, l'érable, le tulipier, le peuplier, le tremble, le bouleau, le merisier, le platane, le sassafras, et l'aulne. Plus stables que la chlorophylle, leur teneur diminue moins vite au cours de la sénescence[8].
Les caroténoïdes se retrouvent couramment dans le monde du vivant et sont notamment responsables de la couleur caractéristique de la carotte, du maïs, du canari, des jonquilles, ainsi que celle du jaune d'œuf et de la banane.
Anthocyanes
Les couleurs rouges et pourpres, et leurs combinaisons qui parent les feuillages d'automne, proviennent d'une autre famille de pigments appelées anthocyanes. À la différence des caroténoïdes, ces pigments ne sont généralement pas présents dans la feuille au cours de la période de croissance. Ces molécules issues de la dégradation de sucres, sont synthétisées à la fin de l'été. Avant de se débarrasser de ses feuilles, l'arbre les vide des nutriments importants, comme le phosphate. Ceci modifie leur métabolisme. La dégradation des sucres mène alors à la synthèse de novo des anthocyanes dont les couleurs rouges attirent moins les pucerons que les couleurs vertes des feuilles estivales, ces insectes pondant moins sur les feuilles automnales[9] - [10].
Pigments bruns
Le stade final de la sénescence de la feuille s'accompagne de la désintégration des chromoplastes et d'une oxydation enzymatique des composés phénoliques (de type tanins. Bien qu'elles soient coriaces et peu goûteuses, il est possible de mâcher les feuilles pour évaluer l'évolution de ces tanins colorés par l'oxydation. Ils sont à l'origine du goût amer et astringent des feuilles vertes, et celui encore plus prononcé des feuilles jaune-orangé. La complexation des tanins avec les protéines lors du stade final de la sénescence, explique le goût terne de carton des feuilles brunes qui ont pris une couleur tabac (nommée ainsi à cause des feuilles mortes du tabac)[11]. Le paradoxe des complexes tanins-protéines brunâtres est qu'ils limitent la digestibilité des protéines par les décomposeurs classiques de la litière qui n'ont pas l'équipement enzymatique capable de les digérer mais les champignons mycorhiziens des arbres ont cet équipement (phosphatases, protéases) et récupèrent l'azote et le phosphore au profit de la symbiose mycorhizienne[12].
La faculté de percevoir la coloration, l'amertume et l'astringence de ces polyphénols a probablement été sélectionnée au cours de l'évolution pour avertir les herbivores et les omnivores de l’absorption de ces composés potentiellement toxiques[13] conduisant à la formation de « pigments bruns », molécules organiques de fort poids moléculaire dont la structure chimique est encore très mal connue[14].
Le changement de couleur progressif des feuilles au cours de l'automne
Le changement de couleur, lié au processus de sénescence, est variable au sein du feuillage, et même au sein d'une feuille. Cette variation est due à plusieurs facteurs : variation du pH vacuolaire, variation des concentrations d'ions métalliques qui chélatent certains hydroxyles et modifient la couleur du pigment, degré de dégradation de copigments comme les flavonols et les caroténoïdes, polymérisation et oxydation des tanins[5].
Les changements de couleur des feuilles dans la litière
La mort des cellules végétales des feuilles entraîne la dégradation de tous les pigments et la diffusion des tanins vacuolaires qui précipitent avec les protéines cytosoliques, ce qui explique que la couleur des feuilles vire vers des couleurs brunes de plus en plus foncées puis noires. Puis apparaissent sur le limbe foliaire des taches plus claires voire blanches, de quelques millimètres à plusieurs dizaines de centimètres englobant tout un paquet de feuilles et de débris. Elles correspondent au développement de mycéliums saprophytes qui décomposent les composés colorés (les tanins combinés aux protéines), la lignine puis la cellulose. Cette « flore » mycologique des pourritures blanches de la litière[15] appartient souvent à des basidiomycètes (genres Marasmius, Clitocybe, Collybia, Lepista, Agaricus, Lepiota, etc.)[16].
Rôles de ces pigments
Plusieurs hypothèses physiologiques (fonction de déchets, invalidée car la production de pigments est coûteuse en énergie[17] ; fonction de photoprotection, de lutte contre le froid[18] et effet antioxydant)[19] aux feuilles pendant qu'elles libèrent certains de leurs éléments nutritifs vers le tronc et les racines) ou des hypothèses écologiques (coévolution arbre-insecte ; pigments bruns fixant les protéines, l'azote et le phosphate[20], composés minéraux formant des réserves progressivement libérées et rendues biodisponibles pour la consommation des plantes grâce à l'activité fongique rhizosphérique)[21] - [22] sont proposées pour expliquer le rôle des pigments rouges[23] et des pigments bruns. Les tanins qui sont déjà eux-mêmes des pigments anthocyanes (rouges ou jaunes)[24] sont en effet impliqués dans la répulsion des insectes tels que les pucerons qui pondent leurs œufs sur des arbres en fin de saison. Au printemps, ces œufs éclosent et les larves phytophages se nourrissent sur leur hôte avec des effets parfois dévastateurs[25]. Cette couleur aposématique de défense des plantes contre les herbivores intervenant au cours de l'évolution de la communication visuelle indiquerait aux insectes non seulement la toxicité des feuilles automnales (hypothèse du signal honnête affichant cette toxicité) mais aussi leurs ressources alimentaires limitées, et enrayerait l'homochromie de certains insectes verts ravageurs plus soumis à la prédation[26]. Inversement cette couleur est un signal positif, attirant les pollinisateurs des plantes, les frugivores, ce qui favorise l'endozoochorie[27].
Il y a plus d'espèces d'arbres produisant en automne des feuilles rouges (typiquement les Érables) en Asie et en Amérique du Nord qu'en Europe. Cette différence s'expliquerait par la nature tropicale de ces arbres apparus au Paléogène, il y a 35 millions d’années. Au gré de la tectonique des plaques, le climat devient peu à peu plus modéré (série de réchauffements et de glaciations) et voit l'apparition de saisons, ces arbres devant notamment résister à leurs défoliateurs automnaux grâce à leurs feuilles rouges. Il y a trois millions d'années, une nouvelle glaciation aurait contribué à la migration de ces arbres vers les régions situées au sud, plus chaudes. En Europe, les chaînes de montagne orientées est-ouest (essentiellement les Alpes) auraient bloqué ces migrations, les arbres ainsi que leurs ravageurs survivant mal au climat périglaciaire[28].
Galerie
_crop.jpg.webp) Feuilles d'automne
Feuilles d'automne

 Érable plane (Acer platanoides) en couleurs automnales
Érable plane (Acer platanoides) en couleurs automnales



 Feuilles de hêtre européen (Fagus sylvatica)
Feuilles de hêtre européen (Fagus sylvatica) Un érable palmé en automne
Un érable palmé en automne Feuilles d'un érable palmé
Feuilles d'un érable palmé Forêt du parc Rivière-du-Moulin durant l'automne à Chicoutimi
Forêt du parc Rivière-du-Moulin durant l'automne à Chicoutimi Feuille restant retenue dans l'herbe
Feuille restant retenue dans l'herbe
Articles connexes
Références
- (en) D.M. Glass, Bruce A. Bohm, « The accumulation of cinnamic and benzoic acid derivatives in Pteridium aquilinum and Athyrium felix-femina », Phytochemistry, vol. 8, no 2, , p. 371-377 (DOI 10.1016/S0031-9422(00)85433-3)
- Au total, 10 % des espèces d'arbres des régions tempérées rougissent et 15 % jaunissent mais quelles que soient leurs nuances de jaune ou de rouge à l’origine, toutes les feuilles finissent brunes. « Les changements de couleurs affectent aussi des arbres et des arbustes des régions tropicales : mais comme, chez eux, poussée et chute des feuilles sont continuelles, car les feuilles ne sont pas synchrones entre elles faute de saisons, le phénomène est moins spectaculaire ». Cf Marc-André Selosse, Les Goûts et les couleurs du monde. Une histoire naturelle des tannins, de l'écologie à la santé, Actes Sud, , p. 83
- (en) P. Matile, « Fluorescent idioblasts in autumn leaves of Ginkgo biloba », Bot. Helv., vol. 104, , p. 87-92.
- (en) John Albert Raven, « A cost-benefit analysis of photon absorption by photosynthetic unicells », New Phytol, vol. 98, no 4, , p. 593–625 (DOI 10.1111/j.1469-8137.1984.tb04152.x).
- (en) P. Matile, « Biochemistry of Indian summer: physiology of autumnal leaf coloration », Exp Gerontol., vol. 35, no 2, , p. 145-158.
- Cette chute est généralement rendue nécessaire par le fait que les feuilles sont vulnérables au gel (à l'exception des feuilles persistantes), en raison du phénomène appelé embolie hivernale, faisant apparaître lors du gel des microbulles de gaz dans la sève glacée (les gaz dissous dans cette sève sont peu solubles). Au moment du dégel, ces bulles d’air grossissent et provoquent l’interruption de la circulation de sève si les tensions sont suffisantes. Si l'arbre gardait ses feuilles en hiver, elles dépériraient par déshydratation à cause de cette embolie. Le même phénomène a lieu en cas de sécheresse prolongée. Lors de ces événements critiques, les végétaux émettent des ultrasons décelés avec des appareils acoustiques adaptés, qu'on eut interprété, selon une vision anthropomorphique, comme un « cri d’alarme » anticipant l’embolie. Cf. (en) A. Ponomarenko, O. Vincent, A. Pietriga, H. Cochard, É. Badel, P. Marmottant, « Ultrasonic emissions reveal individual cavitation bubbles in water-stressed wood », Journal of the Royal Society Interface, vol. 11, no 99, (DOI 10.1098/rsif.2014.0480)
- (en) Stephen G. Pallardy, Physiology of Woody Plants, Academic Press, , p. 212-213.
- (en) P. Matile, « Biochemistry of Indian summer: physiology of autumnal leaf coloration », Experimental Gerontology, vol. 35, no 2, , p. 145-158.
- (en) Feild TS, Lee DW, Holbrook NM, « Why leaves turn red in autumn. The role of anthocyanins in senescing leaves of red-osier dogwood », Plant Physiology, vol. 127, no 2, , p. 566–574.
- (en) Jon E. Sanger, « Quantitative Investigations of Leaf Pigments From Their Inception in Buds Through Autumn. Coloration to Decomposition in Falling Leaves », Ecology, vol. 52, no 6, , p. 1075-1089.
- Marc-André Selosse, Les Goûts et les couleurs du monde. Une histoire naturelle des tannins, de l'écologie à la santé, Actes Sud, , p. 84
- (en) Kraus T. E. C., Yu Z., Preston C. M., Dahlgren R. A. & Zasoski R. J. 2003b. Linking chemical reactivity and protein precipitation to structural characteristics of foliar tannins. Journal of Chemical Ecology, 29: 703–730.
- (en) Georg G. Gross, Richard W. Hemingway, Takashi Yoshida, Plant Polyphenols : Chemistry, Biology, Pharmacology, Ecology, Springer, , p. 877.
- [PDF] Stéphane Jacquemoud, Jean-Baptiste Féret & Susan L. Ustin, Compréhension et modélisation de la couleur des feuilles, École thématique interdisciplinaire du CNRS – Couleur, question d’échelle : l’espace, 2009, p. 185
- Appelées aussi pourritures blanches de l'humus, elles se distinguent de la pourriture blanche du bois et de la pourriture brune sous l'action desquelles le bois acquiert une texture cubique puis se transforme progressivement en une masse pulvérulente brune.
- Guy Durrieu, Écologie des champignons, Masson, , p. 45.
- (en) Mark N. Merzlyak, Anatoly Gitelson, « Why and What for the Leaves Are Yellow in Autumn? On the Interpretation of Optical Spectra of Senescing Leaves (Acerplatanoides L.) », Journal of Plant Physiology, vol. 145, no 3, , p. 315-320 (DOI 10.1016/S0176-1617(11)81896-1).
- Les tannins captent les radiations non-rouges dans des feuilles rouges et les relâchent sous forme de chaleur. L’été indien au Canada, est plus coloré qu'en Europe de l'Ouest : « il y a plus de tannins, or justement, les automnes sont plus lumineux et plus frais là-bas ! » Cf. Marc-André Selosse, « Les couleurs et les goûts de l'automne », sur franceinter.fr,
- (en) Mark N.Merzlyak, Anatoly Gitelson, « Why and What for the Leaves Are Yellow in Autumn? On the Interpretation of Optical Spectra of Senescing Leaves (Acerplatanoides L.) », Journal of Plant Physiology, vol. 145, no 3, , p. 315-320 (DOI 10.1016/S0176-1617(11)81896-1).
- « Ces pigments bruns emprisonnent tous les restes cellulaires : il y a là de l’azote et du phosphate. Les pigments bruns sont trop gros pour être entraînés par l’eau : au sol, les feuilles restent brunes. Écologiquement, c’est vital car sans cela, l’azote et le phosphate seraient emportés par les pluies hivernales ! Avec le redoux printanier, les microbes du sol attaqueront ces pigments bruns et libéreront lentement l’azote et le phosphate. Et à ce moment-là, les racines pourront les re-capter ! Les feuilles brunes sont un engrais à libération retardée ! Grâce aux pigments bruns, ce qui n’a pas été récupéré avant la chute des feuilles est recyclé sous l’arbre. Les tannins colorés d’automne préparent les pigments bruns qui enrichissent le sol. Voilà pourquoi les plantes annuelles ne changent pas de couleur : améliorer le sol l’année suivante leur est inutile ! » Cf. Marc-André Selosse, « La chronique du vivant », sur franceinter.fr,
- (en) W. D. Hamilton, Sam Brown, « Autumn tree colors as a handicap signal », Proceedings of the Royal Society B: Biological Sciences, vol. 268, no 1475, , p. 1489-1493 (DOI 10.1098/rspb.2001.1672, lire en ligne)
- (en) Marco Archetti, « The Origin of Autumn Colours by Coevolution », Journal of Theoretical Biology, vol. 205, no 4, , p. 625-630 (DOI 10.1006/jtbi.2000.2089).
- (en) W.A. Hoch, E.L. Zeldin, B.H. McCown, « Physiological significance of anthocyanins during autumnal leaf senescence », Tree Physiol., vol. 21, no 1, , p. 1-8.
- Les anthocyanes sont en effet des tanins, comme le confirme le mâchage d'une feuille jaune ou rouge, donnant une sensation amère mêlée d'astringence.
- (en) David W. Lee, « Anthocyanins in autumn leaf senescence », Advances in Botanical Research, vol. 37, , p. 147-165 (DOI 10.1016/S0065-2296(02)37048-4).
- (en) Simcha Lev-Yadun, Amots Dafni, Moshe A. Flaishman, Moshe Inbar, Ido Izhaki, Gadi Katzir, Gidi Ne'eman, « Plant coloration undermines herbivorous insect camouflage », BioEssays, vol. 26, no 10, , p. 1126–1130 (DOI 10.1002/bies.20112).
- (en) S. Lev-Yadun & K. S. Gould, « Role of Anthocyanins in Plant Defence ». Springer (Ed). In Anthocyanins: Biosynthesis, Functions, and Applications, 2009, p.1-28
- (en) Simcha Lev-Yadun, Jarmo K. Holopainen, « Why red-dominated autumn leaves in America and yellow-dominated autumn leaves in Northern Europe? », New Phytologist, vol. 183, no 3, , p. 506–512 (DOI 10.1111/j.1469-8137.2009.02904.x).