Inostrancevia
Inostrancevia est un genre éteint de grands thérapsides carnivores ayant vécu durant le Permien supérieur dans ce qui sont aujourd'hui la Russie européenne et l'Afrique du Sud. Les premiers fossiles connus de ce gorgonopsien ont été découverts dans la Dvina septentrionale, en Russie, durant la fin du XIXe siècle, où deux squelettes quasi-complets ont été exhumés. Par la suite, plusieurs autres matériel fossiles ont été découverts dans d'autres oblasts, et les divers trouvailles conduiront à une confusion quand aux nombre exact d'espèces valides au sein de ce pays, avant que seulement trois d'entre eux ne soit officiellement reconnues : I. alexandri, I. latifrons et I. uralensis. Des recherches plus récentes menée en Afrique en Sud ont découverts des restes assez biens préservés du genre, étants attribués à l'espèce nommé I. africana. Le genre est nommé en l'honneur d'Alexandre Inostrantsev, professeur de Vladimir P. Amalitsky, le paléontologue ayant décrit le taxon.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Sous-ordre | † Gorgonopsia |
| Famille | † Gorgonopsidae |
| Sous-famille | † Inostranceviinae |
Espèces de rang inférieur
- † Amalitzkia Pravoslavlev, 1927
- I. proclivis Pravoslavlev, 1927
- Amalitzkia vladimiri Pravoslavlev, 1927
- Amalitzkia annae Pravoslavlev, 1927
- I. vladimiri Vyushkov, 1953
Inostrancevia est le plus grand représentant connu des gorgonopsiens, les plus imposants spécimens fossiles indiquant une taille estimé entre 3 et 3,5 m de long. Il est caractérisée par son squelette très robuste, son crâne large et d'une dentition très avancée, possédant de grandes canines dont les plus longues peuvent atteindre 15 cm et qui peuvent avoir servi à cisailler la peau des proies. Comme la plupart des autres gorgonopsiens, Inostrancevia avait un angle d'ouverture de la mâchoire particulièrement grande, ce qui l'aurait permis à infliger des morsures fatales. Ces caractéristiques font de lui l'un des tétrapodes superprédateurs les plus spécialisés du Paléozoïque.
D'abord régulièrement classé comme proche des taxons africains tels que Gorgonops ou les rubidgeinés, les analyses phylogénétiques publiées depuis 2018 le considèrent comme appartenant à un regroupement de gorgonopsiens russes dérivées, étant désormais classés aux côtés des genres Suchogorgon, Sauroctonus et Pravoslavlevia. D'après les archives fossiles russes, la faune où a vécu Inostrancevia était un désert froid possédant un écosystème fluviaux contenant de nombreux tétrapodes, où il s'avère avoir été le principal prédateur.
Historique des recherches
Espèces reconnues
Durant les années 1890, le paléontologue russe Vladimir Amalitsky découvre des sédiments d'eaux douces datant du Permien supérieur au sein de la Dvina septentrionale, dans l'oblast d'Arkhangelsk, au nord de la Russie européenne. La localité consiste en un ruisseau avec des grès et des expositions en forme de lentille (en) dans un escarpement de berge, contenants de nombreux squelettes fossiles particulièrement bien conservés[1]. La trouvaille de ce type de faune datant de cette période, auparavant connu uniquement d'Afrique du Sud et d'Inde, est considérée comme l'une des plus grandes découvertes paléontologiques de la fin du XIXe siècle et du début du XXe siècle[2]. Après la reconnaissance préliminaire du lieu, Amalitsky effectue des recherches systématiques avec sa compagne Anna Petrovna (ru)[1]. Les exhumations des fossiles durent alors jusqu'en 1914, année où les recherches s'arrête en raison du début la Première Guerre Mondiale[2]. Les fossiles découverts au sein du lieu seront par la suite déplacé au musée de géologie et de minéralogie de l'académie des sciences de Russie. Tous les fossiles répertoriés ne furent pas préparés, et plus de 100 tonnes de concrétions furent promis à de nouvelles découvertes par le musée en question[1].

Les multiples activités administratives et les conditions difficiles durant les dernières années d'Amalitsky ont sévèrement entravé ses recherches sur les fossiles, conduisant à sa mort en 1917. Cependant, parmi tous les fossiles identifiés avant son décès, figure deux squelettes remarquablement complets de grand gorgonopsiens, catalogués PIN 1758 et PIN 2005/1578 (auquel ce dernier sera plus tard reconnu comme le lectotype du genre[2] - [3]). Après identification, il attribue les deux spécimens à un nouveau genre et espèce, qu'il nomme Inostranzevia alexandri[1]. Bien qu'il ne fournit pas l'étymologie du terme dans ses descriptions, le nom complet du taxon est nommé en l'honneur du géologue réputé Alexandre Inostrantsev (ru), qui n'est autre que le professeur d'Amalitsky lui-même[3]. Ses descriptions seront officiellement publiées à titre posthume en 1922, et les deux squelettes seront mis sous la propriété de l'institut de paléontologie de l'Académie des sciences de Russie. Cependant, l'article d'Amalitsky décrit généralement toute les découvertes fossiles effectués dans la Dvina septentrionale, et non Inostrancevia en lui-même, l'article mentionnant que d'autres recherches sur ces gorgonopsiens étant sujets à recherches[1].
C'est en 1927 qu'un des collègues d'Amalitsky, Pavel Pravoslavlev (ru), publie la première description formelle du genre. Dans sa monographie, il nomme plusieurs espèces supplémentaires[N 1] et révise en détail la morphologie des deux squelettes connus d'I. alexandri[4]. Parmi les espèces nommées, seul I. latifrons est reconnue comme étant une espèce clairement distincte au sein du genre, étant basée sur des crânes découverts au sein de l'oblast d'Arkhangelsk ainsi que d'un squelette très incomplet provenant du village de Zavrazhye (en), situé dans l'oblast de Vladimir[2]. L'épithète spécifique latifrons vient du latin latus « large » et frōns « front », en référence à la taille et la constitution crânienne plus robuste qu'I. alexandri[4]. Dans son ouvrage, Pravoslavlev change d'ailleurs la typographie du nom « Inostranzevia » en « Inostrancevia »[4]. Ce dernier terme est depuis entrée dans l'usage universel et doit être maintenue selon la règle de l'article 33.3.1 de la CINZ[5]. Bien que le travail de Pravoslavlev ait été d'une importance majeure, une réexamination de son anatomie squelettique est nécessaire afin d'élargir la compréhension du mode de vie de l'animal[6].
| Image externe | |
| Photographie et croquis du spécimen holotype d'I. uralensis. | |
En 1974, Leonid Tatarinov décrit la troisième espèce, I. uralensis, sur la base de rares restes d'une partie de la boîte crânienne provenant d'un individu plus petit que les deux autres espèces reconnues. L'épithète spécifique uralensis fait référence à la rivière Oural, situé dans l'oblast d'Orenbourg, lieu d'où fut trouvé le spécimen holotype du taxon[7] - [3].
Entre 2010 et 2011, Nthaopa Ntheri et John Nyaphuli découvrent deux spécimens de grand gorgonopsiens lors d'un travail de terrain dirigé par Jennifer Botha, co-auteur de l'université du Witwatersrand, dans la ferme de Nooitgedacht, au sein du bassin du Karoo, en Afrique du Sud. C'est en 2023, après la préparation des fossiles, que Christian F. Kammerer et ses collègues publient une révision qui confirme à leur grande surprise que ces spécimens appartiennent à Inostrancevia, ce qui est une grande première, car ce genre fut auparavant signalée qu'en Russie. Cependant, ces spécimens possèdent quelques différences permettant de le distinguer des espèces russes, étant alors classé dans l'espèce nouvellement érigé I. africana, l'épithète spécifique faisant référence à l'Afrique, continent d'où le taxon a vécu. L'article décrivant officiellement cet animal se porte principalement sur l'importance stratigraphique des découvertes et n'est qu'une brève introduction à l'anatomie du nouveau matériel fossile, ces derniers étant sujets pour une étude qui sera publiée ultérieurement[8].
Autres espèces attribuées
En raison de la mauvaise qualité de préservation des certains fossiles d'Inostrancevia, plusieurs spécimens furent donc considérés à tort comme appartenant à des taxons distincts. Actuellement, seules quatre espèces sont reconnues, I. alexandri, I. latifrons, I. uralensis et I. africana[7] - [2] - [5] - [8].
Dans sa monographie de 1927, Pravoslavlev nomme deux espèces supplémentaires du genre Inostrancevia : I. parva et I. proclivis[4]. En 1940, le paléontologue Ivan Efremov émet des doutes sur cette classification, et considère que le spécimen holotype de I. parva doit être vue comme un juvénile du genre et non comme une espèce distincte[9] - [5]. C'est en 1953 que Boris Pavlovitch Vjuschkov révise complétement les espèces nommées pour Inostrancevia. Pour I. parva, il le déplace dans un nouveau genre, qu'il nomme Pravoslavlevia, en l'honneur à l'auteur original ayant nommé l'espèce[10]. Bien qu'étant un genre distinct et valide, il s'avère que Pravoslavlevia est un taxon étroitement apparenté[2] - [5] - [11] - [12]. Toujours dans son article, il considère qu'I. proclivis n'est en fait qu'un synonyme junior d'I. alexandri, mais reste ouvert à la question de l'existence de cette espèce, arguant de son opinion avec la conservation insuffisante des spécimens types[10]. Ce taxon se fera définitivement jugée comme étant conspécifique à I. alexandri dans la révision du genre menée par Tatarinov en 1974[7].
Toujours dans l'ouvrage, Pravoslavlev nomme un autre genre de gorgonopsiens, Amalitzkia, avec les deux espèces qu'il inclut : A. vladimiri et A. annae, toutes deux nommées en honneur au couple de paléontologues ayant effectué les travaux sur les premiers spécimens connus d'I. alexandri[4]. En 1953, Vjuschkov découvre que le genre Amalitzkia n'est qu'un synonyme junior d'Inostrancevia, renommant A. vladimiri en I. vladimiri[10], avant que ce dernier ne soit lui-même reconnu en tant que synonyme junior d'I. latifrons par des publications ultérieures[7] - [2]. Pour une raison peu claire, Vjuschkov désigne A. annae comme un nomen nudum, alors que sa description est tout à fait viable[10]. Tout comme A. vladimiri, A. annae sera synonymisé avec I. latifrons par Tatarinov en 1974[7].
D'autre espèces appartenant à des genres, voire à des lignées distinctes, furent parfois classés par inadvertance dans le genre Inostrancevia. Par exemple, en 1940, Efremov classe un gorgonopsien au statut à l'époque problématique sous le nom d'I. progressus[2]. Cependant, en 1955, Alexeï Bystrov déplace cette espèce le genre distinct Sauroctonus[13] - [2] - [5]. Un grand os maxillaire découvert dans l'oblast de Vladimir durant les années 1950 fut également attribué à Inostrancevia, mais le fossile sera réattribué à un grand thérocéphale en 1997, et plus tard désigné comme l'holotype du genre Megawhaitsia[14].
Description
Taille
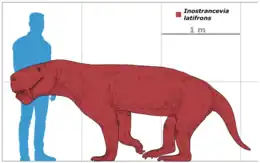
Les spécimens PIN 2005/1578 et PIN 1758, appartenant à I. alexandri, figurent parmi les plus grands et les plus complets des fossiles de gorgonopsiens ayant été identifiés à ce jour. Les deux spécimens mesurent environ 3 m de long, les crânes mesurant à eux seuls plus de 50 cm[1]. Cependant, I. latifrons, bien que connu à partir de fossiles moins complets, est estimé d'avoir eu une taille plus imposante, le crâne mesurant 60 cm de long, indiquant qu'il aurait mesuré 3,5 m pour un poids de 300 kg[15]. La taille d'I. uralensis est inconnue en raison des fossiles très incomplets, mais il semble être plus petit qu'I. latifrons[2].
Anatomie globale
.jpg.webp)
Presque toutes les descriptions effectuées sur le genre dans l'ensemble concernent I. alexandri, les autres espèces n'étant documentées que par des restes partiels ou très fragmentaires[2]. L'anatomie squelettique d'I. africana est sujet pour une future étude plus approfondi, car l'article le décrivant se porte sur l'importance stratigraphique des découvertes et non sur sa morphologie[8].
La forme globale du crâne d'Inostrancevia est typiquement semblable à ceux des autres gorgonopsiens[1], bien qu'il possède certaines différences permettant de le distinguer des représentants africains[5]. Il possède un crâne élargi en arrière, un museau relevé et allongé, des orbites relativement petites et des arcs crâniens minces[2] - [6]. Le foramen pinéal est situé près du bord postérieur des pariétaux et repose sur une forte projection au milieu d'un creux allongé comme impression[1]. La suture sagittale est renforcée par des courbures complexes. La surface ventrale des os palatins est complètement lisse, manquant de trace de dents palatines ou de tubercules. Tout comme Viatkogorgon, la marge supérieure du carré est épaissie[6]. Les trois espèces russes reconnues se distinguent par des caractéristiques notables entre eux : I. alexandri se démarque par son occiput relativement étroit, une fosse temporale ovale large et arrondie ainsi que les brides transversales du ptérygoïde avec des dents ; I. latifrons se distingue par un museau comparativement plus bas et plus large, une région pariétale plus grande, moins de dents ainsi que des tubérosités palatines moins développées ; et I. uralensis est caractérisée par une fosse temporale ovale en forme de fente allongée transversalement[2].

Les mâchoires d'I. alexandri sont puissamment développées, équipées de dents capables de retenir et de déchirer la peau des proies. Les dents sont d'ailleurs dépourvues de cuspides et se distinguent en trois types : les incisives, les canines et les postcanines[N 2]. Toutes les dents sont plus ou moins compressées latéralement et ont des bords avant et arrière finement dentelés[6]. Lorsque la bouche est fermée, les canines supérieures se mettent en position sur les côtés de la mandibule, atteignant son bord inférieur[1]. Les canines d'I. alexandri mesurant entre 12 et 15 cm, ils figurent parmi les plus grandes identifiées au sein des thérapsides pré-mammaliens[6], seul l'anomodonte Tiarajudens ayant des canines de taille similaire[16]. Dans les mâchoires supérieures et inférieures, ces canines sont de tailles à peu près égales et sont légèrement incurvées[6]. Les incisives s'avèrent être très robustes. Les dents postcanines sont présentes sur la mâchoire supérieure, sur ses bords alvéolaires légèrement tournés vers le haut. En revanche, ils sont totalement absents de la mâchoire inférieure. Il y a des indications que le remplacement dentaire aurait eu lieu par les dents jeunes, se développant à la racine des anciennes et les supplantant progressivement[1]. La capsule des canines est très large, contenant jusqu'à trois capsules de canines de remplacement à différents stades de développement[6].
Le squelette est de constitution très robuste. Les phalanges unguéales ont une forme triangulaire aiguë[1] - [4] - [6].
Classification
Dans la description originale publiée en 1922, Inostrancevia fut initialement classé comme un gorgonopsien proche du genre africain Gorgonops[1]. Par la suite, peu de gorgonopsiens seront répertoriés en Russie, mais l'identification de Pravoslavlevia marquera un nouveau tournant dans sa classification. En 1974, Tatarinov classe les deux genres dans la famille des Inostranceviidae[7]. En 1989, Denise Sigogneau-Russell propose une classification similaire, mais déplace le taxon réunifiant les deux genres en tant que sous-famille, étant renommé Inostranceviinae, et est classé dans la famille plus générale des Gorgonopsidae[17]. En 2002, dans sa révision des gorgonopsiens russes, Mikhaïl F. Ivakhnenko érige à nouveau la famille des Inostranceviidae et classe le taxon en tant qu'une des lignée de la super-famille des « Rubidgeoidea », placé au côtés des Rubidgeidae et des Phtinosuchidae (en)[18]. Un an plus tard, en 2003, il reclasse Inostrancevia dans la famille des Inostranceviidae, à la manière de la proposition de Tatarinov, mais ce dernier le classe seul, faisant de ce dernier un taxon monotypique[19]. En 2007, sur la base des observations effectués sur les os occipitaux et les canines, Eva V. I. Gebauer déplace Inostrancevia en tant que taxon frère des Rubidgeinae, un lignée constituée de robustes gorgonopsiens africains[20]. En 2016, Christian Kammerer, dans sa révision des Rubidgeinae, considère la classification de Gebauer comme « insatisfaisante », citant que bon nombre des caractères utilisés par son analyse sont basés sur des proportions de crâne qui sont variables au sein des taxons, à la fois individuellement et ontogénétiquement (c'est-à-dire des traits qui changent au cours de la croissance)[21].
En 2018, dans leur description officielle de Nochnitsa, Kammerer et Vladimir Masyutin proposent que tous les taxons russes et africains doivent être réunis séparément en deux clades distincts. Pour les genres russes (à l'exception des taxons basaux), cette relation s'appuie sur des traits crâniens notables, tels que le contact étroit entre ptérygoïde et le vomer. La découverte d'autres gorgonopsiens russes et la relation entre eux et Inostrancevia ne fut jamais reconnue auparavant[5], pour la simple raison que certains auteurs les rapprochaient indubitablement des genres africains[20]. La classification proposée par Kammerer et Masyutin servira de base pour toutes les autres étude phylogénétiques ultérieures des gorgonopsiens[11] - [12]. Comme pour les classifications précédentes, Pravoslavlevia est toujours considéré comme le taxon frère d'Inostrancevia[5] - [11] - [12].
Ci-dessous, le cladogramme du taxon Gorgonopsia selon Kammerer & Rubidge (2022)[12], qui suit en grande partie les cladogrammes précédemment établis depuis 2018[5] - [11] :
Paléobiologie
L'une des caractéristiques les plus reconnaissables d'Inostrancevia, et des autres gorgonopsiens en général, est la présence de longues canines ressemblant à des sabres sur la mâchoire supérieure et inférieure. La manière dont ces animaux auraient utilisés cette dentition est débattue, la force de morsure des prédateurs à dents de sabre comme Inostrancevia, à l'aide d'analyses tridimensionnelles, fut déterminée par Stephan Lautenschlager et ses collègues en 2020 pour découvrir des réponses[22]. Leurs découvertes détaillent que malgré la convergence morphologique parmi les prédateurs à dents de sabre, il existe une diversité dans les techniques de mise à mort possibles. Le gorgonopsien de taille similaire Rubidgea est capable de produire une force de morsure de 715 newtons. Bien qu'il n'ait pas la force de la mâchoire nécessaire pour être capable d'écraser l'os, l'analyse détaille que les gorgonopsiens les plus massifs possèdent une morsure plus puissante que les autres prédateur à dents de sabre[23]. L'étude indique également que la mâchoire d'Inostrancevia est capable d'une ouverture massive, permettant à ce dernier de délivrer une morsure fatale similaire à la technique de mise à mort hypothétique du célèbre Smilodon, un prédateur à dents de sabre ultérieur[22].
Paléoécologie
Russie européenne


{kind=link}
Durant la fin du Permien, l'Oural méridional était situé autour de la latitude 28-34°N et fut défini comme un « désert froid » dominé par des dépôts fluviaux[24]. La formation de Salarevo en particulier (un horizon d'où est originaire Inostrancevia) se déposait dans une zone saisonnière semi-aride à aride avec de multiples lacs d'eau peu profonde qui sont périodiquement inondés[25]. La paléoflore d'une grande partie de la Russie européenne de l'époque était dominée par un genre de peltaspermacéen (en), Tatarina, et d'autres genres apparentés, suivit par des ginkophytes (en) et des conifères. En revanche, les fougères sont relativement rares et les sphénophytes ne sont présents que localement[24]. On retrouve également des plantes hygrophytes et halophytes dans des zones côtières ainsi que des conifères plus résistants à la sécheresse et aux altitudes plus élevées[26].
Les sites fossilifères d'où Inostrancevia fut répertorié contient d'abondants fossiles d'organismes terrestres et d'eau douce peu profonde, incluant des ostracodes[27], des poissons, des reptiliomorphes comme Chroniosuchus et Kotlassia (en), le temnospondyle Dvinosaurus, le paréiasaure Scutosaurus et le dicynodonte Vivaxosaurus (en)[28] - [26]. Inostrancevia était le principal prédateur de son environnement et aurait pu s'attaquer à la majorité des tétrapodes précédemment mentionnés[28]. D'autres prédateurs plus petits ont existés aux côtés d'Inostrancevia, tels que le plus petit gorgonopsien apparenté Pravoslavlevia et le thérocéphale Annatherapsidus[28] - [2].
Extinction
Les gorgonopsiens, incluant Inostrancevia, ont disparu durant la fin du Lopingien lors de l'extinction Permien-Trias, principalement à cause des activités volcaniques qui sont originaires des trapps de Sibérie. L'éruption qui en a résulté a provoqué un dérèglement climatique important défavorable à leur survie, conduisant à leur extinction. Leur niches écologiques ont cédé la place aux écosystèmes terrestres modernes comprenant les sauropsides, majoritairement des archosaures, ainsi que parmi les rares thérapsides survivant à l'événement, les mammifères[29]. Cependant, certains gorgonopsiens russes ont déjà disparues un peu avant l'événement, ayant délaissé en conséquent certaines de leur niches à des grands thérocéphales[14]. Après l'extinction des rubidgeinés dans leur territoire respectif qu'est l'Afrique, Inostrancevia a migré depuis la Russie pour prendre ainsi, pendant un temps limité, le rôle de superprédateur au sein de ce lieu. La présence de dicynodontes comme Lystrosaurus aurait été une opportunité pour être une proie, car ce dernier a prospéré durant toute l'extinction Permien-Trias[8].
Notes et références
Notes
- L'existence de ces taxons sont déjà mentionnés dans l'article décrivant I. alexandri[1], mais n'ont été officiellement nommé et décrit en détails qu'à partir de 1927[4].
- Précédemment identifié comme des molaires par Amalitsky[1], ce type de dents fut plus tard redécrit comme des dents postcanines, car ayant un manque de portée fonctionnelle[11].
Références
- (en) V. Amalitzky, « Diagnoses of the new forms of vertebrates and plants from the Upper Permian on North Dvina », Bulletin de l’Académie des Sciences de Russie, vol. 16, no 6, , p. 329-340 (lire en ligne)
- (en) Michael J. Benton, Mikhail A. Shishkin, David M. Unwin et Evgenii N. Kurochkin, The age of dinosaurs in Russia and Mongolia, Cambridge, Cambridge University Press, , 696 p. (ISBN 978-0-521-55476-3, lire en ligne)
- (en) « Inostrancevia », sur Paleofile (consulté le )
- (ru) P. A. Pravoslavlev, Gorgonopsidae from the North Dvina expedition of V. P. Amalitzki, vol. 3, Akademii Nauk SSSR, , 1-117 p.
- (en) Christian F. Kammerer et Vladimir Masyutin, « Gorgonopsian therapsids (Nochnitsa gen. nov. and Viatkogorgon) from the Permian Kotelnich locality of Russia », PeerJ, vol. 6, , e4954 (PMID 29900078, PMCID 5995105, DOI 10.7717/peerj.4954
 ).
). - (en) Mikhail F. Ivakhnenko, « Cranial morphology and evolution of Permian Dinomorpha (Eotherapsida) of eastern Europe », Paleontological Journal, vol. 42, , p. 859-995 (DOI 10.1134/S0031030108090013, S2CID 85114195)
- (ru) Leonid Tatarinov, Theriodonts of the USSR, vol. 143, Trudy Paleontologicheskogo Instituta, Akademiya Nauk SSSR, , 1-226 p. (lire en ligne)
- (en) Christian F. Kammerer, Pia A. Viglietti, Elize Butler et Jennifer Botha, « Rapid turnover of top predators in African terrestrial faunas around the Permian-Triassic mass extinction », Current Biology, (PMID 37220743, DOI 10.1016/j.cub.2023.04.007, S2CID 258835757)
- (en) Ivan Yefremov, « On the composition of the Severodvinian Permian Fauna from the excavation of V. P. Amalitzky. », Academy of Sciences of the Union of Soviet Socialist Republics, vol. 26, , p. 893-896
- (ru) Boris P. Vyushkov, « On gorgonopsians from the Severodvinian Fauna », Doklady Akademiya Nauk SSSR, vol. 91, , p. 397-400
- (en) Eva-Maria Bendel, Christian F. Kammerer, Nikolay Kardjilov, Vincent Fernandez et Jörg Fröbisch, « Cranial anatomy of the gorgonopsian Cynariops robustus based on CT-reconstruction », PLOS ONE, vol. 13, no 11, , e0207367 (PMID 30485338, PMCID 6261584, DOI 10.1371/journal.pone.0207367 , Bibcode 2018PLoSO..1307367B)
- (en) Christian F. Kammerer et Bruce S. Rubidge, « The earliest gorgonopsians from the Karoo Basin of South Africa », Journal of African Earth Sciences, vol. 194, , p. 104631 (DOI 10.1016/j.jafrearsci.2022.104631, Bibcode 2022JAfES.19404631K, S2CID 249977414)
- (en) A. P. Bystrow, « A gorgonopsian from the Upper Permian beds of the Volga », Voprosy Paleontologii, vol. 2, , p. 7-18
- (en) M. F. Ivakhnenko, « The first Whaitsiid (Therocephalia, Theromorpha) from the Terminal Permian of Eastern Europe », Paleontological Journal, vol. 42, no 4, , p. 409-413 (DOI 10.1134/S0031030108040102, S2CID 140547244)
- (en) Donald R. Prothero, « 20. Synapsids: The Origin of Mammals », dans Donald R. Prothero, Vertebrate Evolution: From Origins to Dinosaurs and Beyond, Boca Raton, CRC Press, (ISBN 978-0-36-747316-7, DOI 10.1201/9781003128205-4, S2CID 246318785)
- (en) Juan Carlos Cisneros, Fernando Abdala, Bruce S. Rubidge, Paula Camboim Dentzien-Dias et Ana de Oliveira Bueno, « Dental occlusion in a 260-million-year-old therapsid with saber canines from the Permian of Brazil », Science, vol. 331, no 6024, , p. 1603-1605 (PMID 21436452, DOI 10.1126/science.1200305, Bibcode 2011Sci...331.1603C, S2CID 8178585, lire en ligne)
- (en) Denise Sigogneau-Russell, Theriodontia I: Phthinosuchia, Biarmosuchia, Eotitanosuchia, Gorgonopsia, vol. 17 B/I, Stuttgart, Gustav Fischer Verlag, coll. « Encyclopedia of Paleoherpetology », (ISBN 978-3437304873), p. 127
- (en) Mikhail F. Ivakhnenko, « Taxonomy of East European Gorgonopia (Therapsida) », Paleontological Journal, vol. 36, no 3, , p. 283-292 (ISSN 0031-0301)
- (en) M. F. Ivakhnenko, « Eotherapsids from the East European placket », Paleontological Journal, vol. 37, no S4, , p. 339–465
- (en + de) E. V. I. Gebauer, « Phylogeny and Evolution of the Gorgonopsia with a Special Reference to the Skull and Skeleton of GPIT/RE/7113 », Eberhard-Karls University of Tübingen, (lire en ligne [archive du ] [PDF]).
- (en) Christian F. Kammerer, « Systematics of the Rubidgeinae (Therapsida: Gorgonopsia) », PeerJ, vol. 4, , e1608 (PMID 26823998, PMCID 4730894, DOI 10.7717/peerj.1608 )
- (en) Stephan Lautenschlager, Borja Figueirido, Daniel D. Cashmore, Eva-Maria Bendel et Thomas L. Stubbs, « Morphological convergence obscures functional diversity in sabre-toothed carnivores », Proceedings of the Royal Society B, vol. 287, no 1935, , p. 1-10 (ISSN 1471-2954, PMID 32993469, PMCID 7542828, DOI 10.1098/rspb.2020.1818 )
- (en) Julien Benoit, Claire Browning et Luke A. Norton, « The First Healed Bite Mark and Embedded Tooth in the Snout of a Middle Permian Gorgonopsian (Synapsida: Therapsida) », Frontiers in Ecology and Evolution, vol. 6, , p. 699298 (DOI 10.3389/fevo.2021.699298 , S2CID 235487002)
- (en) Massimo Bernardi, Fabio Massimo Petti, Evelyn Kustatscher, Matthias Franz, Christoph Hartkopf-Fröder, Conrad C. Labandeira, Torsten Wappler, Johanna H. A. Van Konijnenburg-Van Cittert, Brandon R. Peecook et Kenneth D. Angielczyk, « Late Permian (Lopingian) terrestrial ecosystems: A global comparison with new data from the low-latitude Bletterbach Biota », Earth-Science Reviews, vol. 175, , p. 18–43 (ISSN 0012-8252, DOI 10.1016/j.earscirev.2017.10.002 , Bibcode 2017ESRv..175...18B, S2CID 134260553)
- (en) E. Yu. Yakimenko, V. O. Targul’yan, N. M. Chumakov, M. P. Arefev et S. A. Inozemtsev, « Paleosols in Upper Permian sedimentary rocks, Sukhona River (Severnaya Dvina basin) », Lithology and Mineral Resources, vol. 35, no 2000, , p. 331–344 (DOI 10.1007/BF02782689, S2CID 140148404)
- (en) Elena Yakimenko, Svyatoslav Inozemtsev et Sergey Naugolnykh, « Upper Permian paleosols (Salarevskian Formation) in the central part of the Russian Platform: Paleoecology and paleoenvironment », Revista Mexicana de Ciencias Geológicas, vol. 21, no 1, , p. 110-119 (S2CID 59417568, lire en ligne [PDF])
- (en) D. A. Kukhtinov, V. R. Lozovsky, S. A. Afonin et E. A. Voronkova, « Non-marine ostracods of the Permian-Triassic transition from sections of the East European platform », Bollettino della Società Geologica Italiana, vol. 127, no 3, , p. 717-726 (lire en ligne)
- (en) Valeriy K. Golubev, « The faunal assemblages of Permian terrestrial vertebrates from Eastern Europe », Paleontological Journal, vol. 34, no 2, , p. 211-224 (lire en ligne)
- (en) Michael J. Benton, « Hyperthermal-driven mass extinctions: killing models during the Permian–Triassic mass extinction », Philosophical Transactions of the Royal Society A, vol. 376, no 2130, (PMID 30177561, PMCID 6127390, DOI 10.1098/rsta.2017.0076 , Bibcode 2018RSPTA.37670076B)
Voir aussi
Articles connexes
Liens externes
- (en) Roman Uchytel, « Reconstitution d'Inostrancevia »
- Ressources relatives au vivant :