Dimetrodon
Dimétrodon

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Clade | Sphenacodontia |
| Famille | † Sphenacodontidae |
| Sous-famille | † Sphenacodontinae |
Espèces de rang inférieur
Dimetrodon, également francisé sous le terme dimétrodon (littéralement « deux tailles de dents »), est un genre éteint d'amniotes appartenant à la famille également éteinte des sphénacodontidés, ayant vécu du début au milieu du Permien, il y a entre 295 et 272 millions d'années avant notre ère. La caractéristique la plus notable de Dimetrodon est sa grande voile d'épine neurale présente sur le dos, formée par des épines allongées s'étendant à partir des vertèbres. C'est un quadrupède ayant un crâne haut et incurvé, avec de grandes dents de différentes tailles placées sur le long des mâchoires. La plupart des fossiles ont été trouvés dans le Sud-Ouest des États-Unis, la majorité provenant de la formation géologique des lits rouges du Texas et de l'Oklahoma. D'autres fossiles ont été découverts au Canada et plus récemment en Allemagne. Plusieurs espèces ont été nommées depuis la description officielle du genre en 1878 par Edward Drinker Cope.
Bien que souvent identifié par erreur comme un dinosaure dans la culture populaire, le genre s'est éteint environ 40 millions d'années avant l'apparition de ces derniers. Malgré son apparence et sa physiologie, Dimetrodon est plus étroitement lié aux mammifères qu'à n'importe quel reptile et est classé comme un représentant non mammalien des synapsides, un groupe incorrectement appelé « reptiles mammaliens », aussi référencés sous différents termes tels que « proto-mammifères » ou « mammifères souches ». Cette classification regroupe Dimetrodon, les mammifères et leurs parents éteints dans un clade appelé Synapsida, tout en plaçant les reptiles, les dinosaures et les oiseaux dans un clade distinct, appelé Sauropsida. Des ouvertures uniques présentes dans le crâne et situés derrière chaque orbite de l'œil, connues sous le nom de fosses temporales, ainsi que d'autres caractéristiques distinguent ce groupe des sauropsides.
Dimetrodon aurait été l'un des principaux prédateurs des écosystèmes du Cisuralien, se nourrissant de poissons et de plus petits tétrapodes, notamment des reptiles et des amphibiens. Les espèces plus petites peuvent avoir eu différents rôles écologiques. La voile de l'animal peut avoir été utilisée pour stabiliser la colonne vertébrale ou pour chauffer et refroidir le corps, comme une forme de thermorégulation. Certaines études récentes affirment que la voile aurait été inefficace pour évacuer la chaleur du corps en raison de la découverte de grandes espèces avec de petites voiles et de petites espèces avec de grandes voiles, ce qui exclurait la régulation de température comme objectif principal. La voile aurait été très probablement utilisée dans les parades nuptiales avec diverses méthodes pour menacer des rivaux ou se montrer à des potentiels partenaires.
Description
Introduction
Dimetrodon est un synapside quadrupède caractérisé par la présence d'une grande voile sur le dos. La plupart des grandes espèces de Dimetrodon varient en des longueurs estimées de 1,7 à 4,6 mètres pour un poids hypothétique situé entre 28 et 250 kg[1]. La plus grande espèce connue du genre est D. angelensis, mesurant environ 4 m et la plus petite est D. teutonis, mesurant environ 60 cm[2] - [3]. Les plus grandes espèces de Dimetrodon figurent parmi les plus grands prédateurs du Permien inférieur, bien que l'étroitement apparenté Tappenosaurus (en), connu à partir de quelques fragments squelettiques découverts dans des roches légèrement plus jeunes, ait pu être encore plus grand avec une longueur corporelle totale estimée à 5,5 m[4] - [5]. Bien que certaines espèces de Dimetrodon puissent devenir très grandes, de nombreux spécimens juvéniles sont connus[6].
Présentation
Une seule grande ouverture de chaque côté de l'arrière du crâne relie Dimetrodon aux mammifères et le distingue de la plupart des premiers sauropsides, qui manquent d'ouvertures ou ont deux ouvertures selon les sous-groupes. Les chercheurs pensent que des caractéristiques telles que des crêtes à l'intérieur de la cavité nasale et une crête à l'arrière de la mâchoire inférieure font partie d'une progression évolutive des premiers vertébrés terrestres à quatre membres aux mammifères[7].
Le crâne de Dimetrodon est grand et comprimé latéralement. Les orbites sont positionnées haut et loin en arrière dans le crâne. Derrière chaque orbite se trouve un trou appelé fosse temporale. Un trou supplémentaire dans le crâne, la fenêtre supratemporale, peut être vu d'en haut. L'arrière du crâne est orienté à un léger angle vers le haut, une caractéristique qu'il partage avec tous les autres synapsides basaux. La marge supérieure du crâne s'incline vers le bas en un arc convexe jusqu'à l'extrémité du museau. La pointe de la mâchoire supérieure, formée par le prémaxillaire, s'élève au-dessus de la partie de la mâchoire formée par le maxillaire pour former une « marche » maxillaire. Au sein même de cette étape se trouve un diastème dans la rangée de dents. Dans son ensemble, le crâne de Dimetrodon est plus solidement construit que celui des dinosaures[7].




Dentition
Le nom générique Dimetrodon signifie littéralement « deux tailles de dents », en référence aux ensembles de dents qui varient considérablement en taille sur la longueur des mâchoires. Une ou deux paires de dents caniniformes, de grandes dents pointues ressemblant à des canines, saillent du maxillaire. De grandes incisives sont également présentes aux extrémités des mâchoires supérieure et inférieure, enracinées dans les prémaxillaires et l'os dentaire. De petites dents sont présentes autour de la « marche » maxillaire et derrière les caniniformes, devenant d'autant plus petites qu'elles s'éloignent dans la mâchoire[8].

De nombreuses dents sont plus larges à leur section médiane et se rétrécissent plus près des mâchoires, leur donnant l'apparence de larmes. Les dents en forme de larme sont uniques à Dimetrodon et à d'autres sphénacodontidés étroitement apparentés, et aident à les distinguer des autres synapsides basaux. Comme chez de nombreux autres synapsides basaux, les dents de la plupart des espèces de Dimetrodon sont dentelées sur leurs bords[3]. Les dentelures des dents de Dimetrodon sont si fines qu'elles ressemblent à de minuscules fissures. Le dinosaure Albertosaurus a des dentelures ressemblant à des fissures similaires, mais à la base de chaque dentelure se trouve un vide rond, qui aurait fonctionné pour répartir la force sur une plus grande surface et empêcher les contraintes de l'alimentation de provoquer la propagation de la fissure à travers la dent. Contrairement à Albertosaurus, les dents de Dimetrodon manquent d'adaptations qui empêcheraient la formation de fissures au niveau de leurs dentelures[9]. Les dents de D. teutonis n'ont pas de dentelures, mais ont toujours des arêtes vives[3].
Une étude publiée en 2014 démontre que les espèces de Dimetrodon auraient adopté différentes morphologies pour chasser divers types de proies. L'une des plus petites espèces, D. milleri, n'a pas de dentelures, car les preuves fossiles montrent qu'il se nourrissait de petites proies. Au fur et à mesure que les proies ont grossi, plusieurs espèces de Dimetrodon ont commencé à développer des dentelures sur leurs dents et à en augmenter la taille. Par exemple, D. limbatus ainsi que Secodontosaurus présentent des dentelures d'émail qui aurait été très utiles à couper la chair d'une proie. La deuxième plus grande espèce connu, D. grandis, a des dentelures similaires à celles des requins et des dinosaures théropodes, ce qui rend ses dents encore plus spécialisées pour trancher la chair. Les différentes proies du dimétrodon ont réagi en évoluant vers des tailles plus grandes et en développant des dents de plus en plus pointues, comme forme de défense au prédateur[10].
Cavité nasale
Sur la surface interne de la section nasale du crâne se trouvent des crêtes appelées nasoturbinaux, qui peuvent avoir soutenu du cartilage qui a augmenté la surface de l'épithélium olfactif, la couche de tissu qui détecte les odeurs. Ces crêtes sont beaucoup plus petites que celles des synapsides ultérieurs datant du Permien supérieur et du Trias, dont les grands nasoturbinaux sont considérés comme une preuve d'homéothermie car ils peuvent avoir soutenu des membranes muqueuses qui réchauffaient et humidifiaient l'air entrant. Ainsi, la cavité nasale du dimétrodon est transitionnelle entre celles des premiers vertébrés terrestres et des mammifères[11].
Articulation des mâchoires et des oreilles
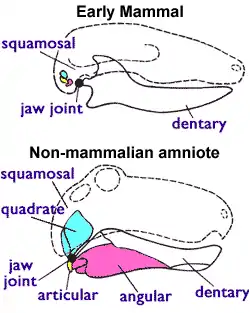
Une autre caractéristique transitoire de Dimetrodon est une crête située à l'arrière de la mâchoire appelée lame réfléchie, qui se trouve sur l'os articulaire, qui se connecte à l'os carré du crâne pour former l'articulation de la mâchoire. Chez les premiers mammifères apparus ultérieurement, l'articulaire et le quadratojugal sont séparés de l'articulation de la mâchoire, tandis que l'articulaire s'est développé pour donner le marteau de l'oreille moyenne. La lame réfléchie est devenue une partie d'un anneau appelé anneau tympanique, qui soutient le tympan chez tous les mammifères vivants[12].
Queue
La queue de Dimetrodon représente une grande partie de la longueur totale du corps de l'animal et comprend environ 50 vertèbres caudales. Les queues ont été manquantes ou incomplètes dans les premiers squelettes décrits de Dimetrodon, les seules vertèbres caudales connues étant les 11 les plus proches de la hanche. Étant donné que ces premières vertèbres caudales se rétrécissent rapidement à mesure qu'elles s'éloignent de la hanche, de nombreux paléontologues de la fin du XIXe siècle et du début du XXe siècle ont pensé que Dimetrodon aurait eu une queue très courte. La première description d'une queue en grande partie complète de Dimetrodon n'a été établie qu'en 1927[13].
Voile

La voile de Dimetrodon est formée d'épines neurales allongées faisant saillie à partir des vertèbres. Chaque épine varie en forme de coupe transversale de sa base à sa pointe dans ce que l'on appelle la différenciation « dimétrodonte »[14]. Près du corps vertébral, la section transversale de la colonne vertébrale est comprimée latéralement en une forme rectangulaire, et plus près de la pointe, elle prend une forme de « 8 » lorsqu'une rainure longe chaque côté de la colonne vertébrale. La forme en huit est censée renforcer la colonne vertébrale, empêchant la flexion et les fractures[15]. Une coupe transversale de la colonne vertébrale d'un spécimen de D. giganhomogenes est de forme rectangulaire mais conserve des anneaux en forme de « 8 » près de son centre, indiquant que la forme des épines peut changer à mesure que les individus vieillissent[16]. L'anatomie microscopique de chaque vertèbre varie de la base à la pointe, indiquant où elle fut intégrée dans les muscles du dos et où est exposée dans le cadre d'une voile. La partie inférieure ou proximale de la colonne vertébrale a une surface rugueuse qui aurait servi de point d'ancrage pour les muscles épaxiaux (en) du dos, et possède également un réseau de tissus conjonctifs appelés fibres de Sharpey qui indiquent qu'il fut intégré dans le corps. Plus haut, sur la partie externe de la colonne vertébrale, la surface osseuse est plus lisse. Le périoste, une couche de tissu entourant l'os, est recouvert de petites rainures qui auraient soutenu vraisemblablement les vaisseaux sanguins qui vascularisaient la voile[17].
Les chercheurs ont pensé autrefois que la grande rainure qui s'étend sur toute la longueur de la colonne vertébrale aurait été un canal pour les vaisseaux sanguins, mais comme l'os ne contient pas de canaux vasculaires, on ne pense pas que la voile ait été aussi fortement vascularisée comme précédemment admis. Certains spécimens de Dimetrodon conservent des zones déformées des épines neurales qui semblent être des fractures cicatrisées. L'os cortical qui s'est développé au cours de ces ruptures est fortement vascularisé, suggérant que des tissus mous devaient être présents sur la voile pour alimenter le site en vaisseaux sanguins[15]. L'os lamellaire en couches constitue la majeure partie de la section transversale de la colonne vertébrale neurale et contient des lignes de croissance arrêtée qui peuvent être utilisées pour déterminer l'âge de chaque individu au décès[18]. Dans de nombreux spécimens de l'espèce D. gigashomogenes, les parties distales des épines se plient fortement, indiquant que la voile aurait eu un profil irrégulier lorsque l'animal fut vivant. Leur courbure suggère que les tissus mous ne se sont peut-être pas étendus jusqu'aux extrémités des épines, ce qui signifie que la sangle de la voile n'aurait peut-être pas été aussi étendue qu'on l'imagine généralement[14].
Peau
Aucune preuve fossile de la peau de Dimetrodon n'a encore été trouvée. Les impressions de la peau d'un animal apparenté, Estemmenosuchus, indiquent qu'elle aurait été lisse et bien pourvue de glandes, mais cette forme de peau peut ne pas s'appliquer à Dimetrodon, car sa lignée est assez éloignée[19]. Dimetrodon peut également avoir eu de grandes écailles sur le dessous de sa queue et de son ventre, comme d'autres synapsides[20] - [21]. Les preuves du varanopidé Ascendonanus (en) suggèrent que certains synapsides basaux peuvent avoir eu des écailles similaires à celles des squamates[22], cependant, certaines études récentes ont rapproché taxonomiquement les varanopidés comme étant plus proches des reptiles diapsides, rendant cette théorie incertaine[23] - [24].
Historique des découvertes
Premières découvertes
_(1886)_(20001979533).jpg.webp)
La première découverte de fossiles de Dimetrodon concerne un maxillaire récupéré en 1845 par un homme du nom de Donald McLeod, vivant dans la colonie britannique de l'Île-du-Prince-Édouard, au Canada. Ces fossiles ont été achetés par John William Johnson, un géologue canadien, puis décrits par Joseph Leidy durant le début des années 1850 comme la mandibule de Bathygnathus borealis, un grand carnivore apparenté selon lui au dinosaure Thecodontosaurus[25] - [26] - [27], bien qu'il ait ensuite été reclassé comme une espèce de Dimetrodon en 2015, sous le nom de D. borealis[28].
Premières descriptions par Edward Drinker Cope
Les fossiles maintenant attribués à Dimetrodon ont été étudiés pour la première fois par le paléontologue américain Edward Drinker Cope durant les années 1870. Cope a obtenu les fossiles du dimétrodon ainsi que ceux de nombreux autres tétrapodes datant du Permien auprès de plusieurs collectionneurs qui ont exploré un groupe de roches présents au Texas appelé « les lits rouges ». Parmi ces collectionneurs figurent le naturaliste suisse Jacob Boll, le géologue texan W. F. Cummins et le paléontologue amateur Charles Hazelius Sternberg[29]. La plupart des spécimens de Cope ont été déplacés au musée américain d'histoire naturelle ou au musée de Walker de l'université de Chicago (la majeure partie de la collection de fossiles de Walker étant maintenant conservée au musée Field).
Sternberg a envoyé certains de ses propres spécimens au paléontologue allemand Ferdinand Broili à l'université de Munich, bien que Broili n'ait pas été aussi prolifique que Cope dans la description des spécimens. Le rival de Cope, Othniel Charles Marsh, a également recueilli des ossements de Dimetrodon, qu'il a envoyés au musée de Walker[30]. La première utilisation du nom Dimetrodon date de 1878 lorsque Cope nomme les espèces D. incisivus, D. rectiformis et D. gigas dans la revue scientifique Proceedings of the American Philosophical Society (en)[31].
La première description d'un fossile de Dimetrodon par Cope date d'un an plus tôt, cependant il y nomme le taxon Clepsydrops limbatus[32]. C. limbatus sera plus tard reclassé comme une espèce de Dimetrodon en 1940, ce qui signifie que l'article de Cope datant de 1877 est le premier enregistrement du genre[33].
Cope a été le premier à décrire un synapside ayant une voile sur le dos avec le nom de C. natalis dans son article de 1878, bien qu'il considère la voile comme une nageoire et la compare aux crêtes du basilic. Les voiles ne sont pas conservées dans les spécimens de D. incisivus et D. gigas que Cope a décrits dans son article de 1878, mais des épines allongées sont présentes dans le spécimen de D. rectiformis, qu'il a également décrit[31]. Cope commente le but de la voile en 1886, en écrivant :
« L'utilité est difficile à imaginer. À moins que l'animal n'ait des habitudes aquatiques et nage sur le dos, la crête ou la nageoire doit avoir gêné les mouvements actifs [...] Les membres ne sont pas assez longs ni les griffes assez aiguës pour démontrer des habitudes arboricoles, comme dans le genre existant Basilicus, où une crête similaire existe[trad 1]. »
Descriptions du début du XXe siècle

Au cours des premières décennies du XXe siècle, le paléontologue américain Ermine Cowles Case (en) est l'auteur de nombreuses études sur Dimetrodon et décrit plusieurs nouvelles espèces. Il reçoit un financement de la Carnegie Institution pour son étude de nombreux spécimens de Dimetrodon dans les collections du musée américain d'histoire naturelle et de plusieurs autres musées[30]. Beaucoup de ces fossiles ont été collectés par Cope mais n'ont pas été décrits en détail, car ce dernier était connu pour ériger de nouvelles espèces sur la base de seulement quelques fragments d'os.
À partir de la fin des années 1920, le paléontologue Alfred Sherwood Romer réétudie de nombreux spécimens de dimétrodons et nomme plusieurs nouvelles espèces. En 1940, Romer co-écrit une grande étude avec Llewellyn Ivor Price, intitulée Review of the Pelycosauria, dans laquelle les espèces de Dimetrodon nommées par Cope et Case sont réévaluées[33]. La plupart des noms d'espèces considérés comme valides par Romer et Price sont encore reconnus de nos jours[17].
Nouveaux spécimens
Dans les décennies qui ont suivi la monographie de Romer et Price, de nombreux spécimens de Dimetrodon ont été décrits dans des localités situées en dehors du Texas et de l'Oklahoma. Le premier a été décrit dans la région des Four Corners (Utah) en 1966[34], tandis qu'un autre a été décrit d'Arizona en 1969[35]. En 1975, Olson publie un article dans lequel il rapporte du matériel de Dimetrodon ayant été découvert dans l'Ohio[36]. En 1977, l'espèce D. occidentalis est décrite à partir de fossiles découverts au Nouveau-Mexique[37]. Les spécimens trouvés dans l'Utah et dans l'Arizona appartiendraient probablement aussi à D. occidentalis[38].
Avant ces découvertes, une théorie proposait qu'une voie maritime du milieu du continent avait séparé de ce qui est maintenant le Texas et l'Oklahoma des terres plus occidentales au début du Permien, isolant Dimetrodon dans une petite région d'Amérique du Nord, tandis que le genre apparenté Sphenacodon dominait la zone ouest. Bien que cette voie maritime ait probablement existé, la découverte de fossiles répertoriés en dehors de ces deux régions montre que son étendue était limitée et qu'elle n'a pas été une barrière efficace à la distribution de Dimetrodon[37] - [39].
En 2001, une nouvelle espèce du genre, D. teutonis, ayant été décrite dans la localité de Bromacker, située dans la forêt de Thuringe en Allemagne, étend pour la première fois l'aire de répartition géographique de Dimetrodon en dehors d'Amérique du Nord[1].
Espèces
Une vingtaine d'espèces attribuées au genre Dimetrodon ont été nommées depuis la première description du taxon en 1878. Beaucoup ont été synonymisés avec d'autres espèces ayant été nommées antérieurement, et certaines appartiennent maintenant à des genres différents.
Listes
| Espèce | Auteur | Lieu de découverte | Statut | Synonymes | Images |
|---|---|---|---|---|---|
| Dimetrodon angelensis | Olson, 1962 | Texas | Valide |  | |
| Dimetrodon borealis | Leidy, 1853 | Canada | Valide | Bathygnathus borealis Leidy, 1853 |  |
| Dimetrodon booneorum | Romer, 1937 | Texas | Valide | ||
| Dimetrodon dollovianus | Case, 1907 | Texas | Valide | Embolophorus dollovianus Cope, 1888 | |
| Dimetrodon giganhomogenes | Case, 1907 | Texas | Valide |  | |
| Dimetrodon grandis | Romer & Price, 1940 | Oklahoma, Texas | Valide | Clepsydrops gigas Cope, 1878 Dimetrodon gigas Cope, 1878 Theropleura grandis Case, 1907 Bathyglyptus theodori Case, 1911 Dimetrodon maximus Romer, 1936 |  |
| ?Dimetrodon kempae | Romer, 1937 | Texas | Possible nomen dubium | ||
| Dimetrodon limbatus | Romer & Price, 1940 | Oklahoma Texas | Valide | Clepsydrops limbatus Cope, 1877 Dimetrodon incisivus Cope, 1878 Dimetrodon rectiformis Cope, 1878 Dimetrodon semiradicatus Cope, 1881 |  |
| Dimetrodon loomisi | Romer, 1937 | Texas, Oklahoma | Valide |  | |
| Dimetrodon macrospondylus | Case, 1907 | Texas | Valide | Clepsydrops macrospondylus Cope, 1884 Dimetrodon platycentrus Case, 1907 | |
| Dimetrodon milleri | Romer, 1937 | Texas | Valide |  | |
| Dimetrodon natalis | Romer, 1936 | Texas | Valide | Clepsydrops natalis Cope, 1878 |  |
| Dimetrodon occidentalis | Berman, 1977 | Nouveau Mexique | Valide | ||
| Dimetrodon teutonis | Berman et al., 2001 | Allemagne | Valide | ||
Dimetrodon borealis
Dimetrodon borealis a été nommé pour la première fois par Leidy en 1853 sous le nom de Bathygnathus borealis à partir d'un maxillaire incomplet ayant été découvert dans la province de l'Île-du-Prince-Édouard, au Canada[25]. Le fossile, catalogué ANSP 9524, sera décrit plus en détail un an plus tard par le même auteur, qu'il considère comme appartenant à un saurien bipède[26]. L'animal étant le tout premier synapside non-mammalien à avoir été découvert et décrit, son histoire taxonomique fut donc très complexe et intrigante au yeux des paléontologues du XIXe siècle, ayant été décrit à plusieurs reprises[28]. Le spécimen à d'abord été considéré comme un dinosaure[27], jusqu'à ce qu'il soit reclassé par Richard Owen en 1876 comme un thériodonte[40] - [N 1], avant d'être correctement identifié comme étant le museau d'un sphenacodontidé durant le début du XXe siècle[28]. Case (en) suggère en 1907 que le maxillaire appartiendrait à un grand Dimetrodon ou à un Naosaurus[30]. En 1940, Romer et Price suggèrent que le spécimen est similaire à D. limbatus et à Sphenacodon ferocior en raison de la taille du maxillaire et des dents[33] mais, en raison du manque de détails de l'arrière du crane, B. borealis fut maintenu comme taxon distinct en attendant d'obtenir une recherche plus approfondie[28].
C'est en 2015 qu'une étude reclasse le spécimen au sein du genre Dimetrodon, le renommant par conséquent D. borealis, et le considérant comme le taxon frère de D. grandis. Le genre Bathygnathus, ayant été nommé 24 ans plus tôt que Dimetrodon, devrait avoir la priorité du nom définitif selon les règlements du CINZ[28]. Cependant, étant donné la large utilisation et la familiarité du nom générique Dimetrodon dans la littérature scientifique et la culture populaire[41], un compromis a donc été trouvé avec la CINZ afin de préserver le nom le plus connu[28].
Dimetrodon limbatus
Dimetrodon limbatus a été décrit pour la première fois par Edward Drinker Cope en 1877 sous le nom de Clepsydrops limbatus[32]. Le genre Clepsydrops a été proposé pour la première fois par Cope en 1875 pour nommer les restes de sphénacodontidés découverts dans le comté de Vermilion, dans l'Illinois, et a ensuite été utilisé pour de nombreux spécimens de sphénacodontidés du Texas, de nombreuses nouvelles espèces de sphénacodontidés de la région ayant été attribuées à Clepsydrops ou à Dimetrodon à la fin du XIXe siècle et au début du XXe siècle. Basé sur un spécimen provenant des lits rouges du Texas, il s'agissait du premier synapside connu ayant une voile sur le dos. En 1940, les paléontologues Romer et Price ont réaffecté C. limbatus à Dimetrodon, faisant de D. limbatus l'espèce type du genre[33].
La première utilisation du nom Dimetrodon est publiée en 1878 lorsque Cope nomme l'espèce D. incisivus avec D. rectiformis et D. gigas. D. rectiformis a été nommé aux côtés de D. incisivus dans l'article de Cope de 1878, et était la seule des trois espèces nommées à préserver des épines neurales allongées[31]. Trois ans plus tard, en 1881, Cope décrit sur la base des os de la mâchoire supérieure sa dernière espèce, D. semiradicatus[42]. En 1907, lorsque Case synonymise D. rectiformis et D. semiradicatus avec D. incisivus[30]. De nos jours, l'espèce D. incisivus est considérée comme un synonyme de D. limbatus[17].
Dimetrodon dollovianus
Dimetrodon dollovianus a été décrit pour la première fois par Cope en 1888 sous le nom Embolophorus dollovianus. En 1903, Case publie une longue description d'E. dollovianus, qu'il référa plus tard au genre Dimetrodon[43].
Dimetrodon grandis
Dans son article de 1878 sur les fossiles du Texas, Cope nomme Clepsydrops gigas avec les premières espèces nommées de Dimetrodon, D. limbatus, D. incisivus et D. rectiformis[31]. Case reclasse C. gigas comme une nouvelle espèce de Dimetrodon en 1907[30]. Case a également décrit un crâne très bien conservé de Dimetrodon en 1904, l'attribuant à l'espèce D. gigas[44]. En 1919, Charles W. Gilmore attribua un spécimen presque complet de Dimetrodon à D. gigas[45]. Actuellement, D. gigas est considéré comme un synonyme de D. grandis[46].
Dans son article datant de 1907, Case nomme une nouvelle espèce de synapside à dos à voile, Theropleura grandis[30]. En 1940, Romer et Price ont réaffecté T. grandis à Dimetrodon, érigeant donc l'espèce D. grandis[33].
Dimetrodon giganhomogenes
Dimetrodon giganhomogenes a été nommé par Case en 1907 et est toujours considéré comme une espèce valide de Dimetrodon[17] - [30].
Dimetrodon macrospondylus
Dimetrodon macrospondylus a été décrit pour la première fois par Cope en 1884 sous le nom de Clepsydrops macrospondylus. En 1907, Case le reclasse comme D. macrospondylus[30]. Toujours dans son article, Case nomme également l'espèce D. platycentrus[30], mais ce taxon est actuellement considéré comme un synonyme de D. macrospondylus[17].
Dimetrodon natalis
Le paléontologue Alfred Romer érige l'espèce Dimetrodon natalis en 1936, précédemment décrite sous le nom de Clepsydrops natalis. D. natalis était la plus petite espèce connue de Dimetrodon à cette époque et a été trouvée aux côtés des restes de la plus imposante espèce D. limbatus[47].
Dimetrodon booneorum
Dimetrodon booneorum a été décrit pour la première fois par Romer en 1937 sur la base de restes découverts au Texas[47].
"Dimetrodon" kempae
Dimetrodon kempae a été nommé par Romer en 1937 dans le même article où il décrivit D. booneorum, D. loomisi et D. milleri[47]. Le taxon a été nommé sur la base d'un seul humérus et de quelques vertèbres, et peut donc être un nomen dubium qui ne peut être distingué comme une espèce unique de Dimetrodon[1]. En 1940, Romer et Price ont évoqué la possibilité que "D". kempae ne relèverait pas du genre Dimetrodon, préférant de le classer comme un incertae sedis[33].
Dimetrodon loomisi
Dimetrodon loomisi a été décrit pour la première fois par Romer en 1937 avec D. booneorum, "D". kempae et D. milleri. Des restes ont été retrouvés au Texas et en Oklahoma[47].
Dimetrodon milleri
Dimetrodon milleri a été décrit par Romer en 1937[47]. C'est l'une des plus petites espèces de Dimetrodon connues d'Amérique du Nord et elle peut être étroitement liée à D. occidentalis, une autre espèce de taille similaire[38]. D. milleri est connu à partir de deux squelettes, un presque complet (MCZ 1365) et un autre moins complet mais plus grand (MCZ 1367). D. milleri est actuellement la plus ancienne espèce connue de Dimetrodon[47].
Outre sa petite taille, D. milleri diffère des autres espèces de Dimetrodon en ce que ses épines neurales sont circulaires plutôt qu'en forme de huit en coupe transversale. Ses vertèbres sont également plus courtes en hauteur par rapport au reste du squelette que celles des autres espèces de Dimetrodon. Le crâne est haut et le museau est court par rapport à la région temporale. Les vertèbres courtes et un crâne haut sont également observés chez les espèces D. booneorum, D. limbatus et D. grandis, ce qui suggère que D. milleri pourrait être le premier d'une progression évolutive entre ces espèces[38].
Dimetrodon angelensis
Dimetrodon angelensis a été nommé par le paléontologue Everett C. Olson en 1962[48]. Des spécimens de l'espèce ont été signalés dans la formation de San Angelo (en) au Texas et est actuellement la plus grande espèce connu du genre[49].
Dimetrodon occidentalis
Dimetrodon occidentalis a été nommé en 1977 à partir de fossiles découverts au Nouveau-Mexique[37]. L'épithète spécifique fait référence pour son statut de première espèce nord-américaine connue de Dimetrodon, celle-ci ayant été découverte à l'ouest du Texas et de l'Oklahoma. Il a été nommé sur la base d'un squelette unique appartenant à un individu relativement petit. La petite taille de D. occidentalis est similaire à celle de D. milleri, suggérant une relation étroite. Des spécimens de Dimetrodon trouvés dans l'Utah et dans l'Arizona appartiendraient probablement aussi à D. occidentalis[38].
Dimetrodon teutonis
Dimetrodon teutonis a été décrit en 2001 à partir de restes fossiles découverts dans la forêt de Thuringe en Allemagne et est la première espèce connue du genre à être décrite hors d'Amérique du Nord. C'est aussi la plus petite espèce du genre Dimetrodon[1].
Dimetrodon cruciger
En 1878, Cope publie un article intitulé « The Theromorphous Reptilia », dans lequel il décrit Dimetrodon cruciger[50]. Selon lui, D. cruciger se distingue par les petites projections qui s'étendent de chaque côté des épines neurales comme les branches d'un arbre[51]. En 1886, Cope déplace D. cruciger dans le genre Naosaurus, car il considère ses épines si différentes de celles des autres espèces de Dimetrodon que l'espèce mérite son propre genre[52]. Naosaurus sera plus tard un synonyme d'Edaphosaurus, un genre que Cope nomma en 1882 sur la base de crânes qui appartenaient manifestement à des animaux herbivores compte tenu de leurs dents broyeuses émoussées[53].
Dimetrodon longiramus
Case nomme l'espèce Dimetrodon longiramus en 1907 sur la base d'une omoplate et d'une mandibule allongée provenant de la formation de Belle Plains au Texas[30]. En 1940, Romer et Price reconnaissent que le matériel de D. longiramus appartient au même taxon qu'un autre spécimen décrit par le paléontologue Samuel Wendell Williston en 1916, qui comprend une mandibule allongée de la même manière et un long maxillaire[33]. Williston n'a pas considéré son spécimen comme appartenant à Dimetrodon mais l'a plutôt classé comme un ophiacodontidé[54]. Romer et Price attribuent les spécimens de Case et Williston à un genre et une espèce nouvellement érigée, Secodontosaurus longiramus, qui est étroitement lié à Dimetrodon[33] - [55].
Classification
Classification classique
Dimetrodon est l'un des premiers membres d'un groupe désigné sous le nom de Synapsida, un clade qui comprend les mammifères et leurs parents aujourd'hui disparus, bien qu'il ne soit l'ancêtre d'aucun d'eux, car appartenant à une lignée distincte. Il est souvent considéré comme un dinosaure dans la culture populaire, bien qu'il ait disparu environ 40 millions d'années avant l'apparition des premiers dinosaures au Trias. En tant que synapside, Dimetrodon est plus étroitement lié aux mammifères qu'aux sauropsides, qu'ils soient vivants ou éteints. Au début des années 1900, la plupart des paléontologues ont considéré Dimetrodon comme un reptile conformément à la taxonomie linnéenne, qui place Reptilia en tant que classe et Dimetrodon en tant que genre au sein du groupe. Les mammifères ont été affectés à une classe distincte et Dimetrodon a été régulièrement décrit comme un « reptile mammalien ». Les paléontologues ont émis l'hypothèse que les mammifères ont évolué à partir de ce groupe dans ce qu'ils ont appelé une transition « reptile à mammifère »[41].
Classification phylogénétique
Selon la classification phylogénétique, les descendants du dernier ancêtre commun de Dimetrodon et de tous les reptiles vivants incluraient tous les mammifères car Dimetrodon est plus étroitement lié à ces derniers qu'aux sauropsides. Ainsi, si l'on souhaite éviter le clade qui contient à la fois les mammifères et les reptiles vivants, alors Dimetrodon ne devrait pas être inclus dans ce clade, ni aucun autre « reptile mammalien ». Les descendants du dernier ancêtre commun des mammifères et des reptiles, qui sont apparus vers environ 310 millions d'années au Carbonifère supérieur, sont donc scindés en deux clades : Synapsida (qui comprend les mammifères et leurs parents éteints, incluant Dimetrodon) et Sauropsida (qui comprend tous les reptiles et les oiseaux, qu'ils soient vivants ou éteints)[41].
Au sein du clade Synapsida, Dimetrodon fait partie du clade Sphenacodontia, qui a été proposé pour la première fois comme un sous-groupe de synapsides basaux en 1940 par les paléontologues Alfred Sherwood Romer et Llewellyn Ivor Price, avec les groupes Ophiacodontia et Edaphosauria[33]. Les trois groupes sont connus des archives fossiles datant du Carbonifère supérieur et du Permien inférieur. Romer et Price les distinguent principalement par des caractéristiques postcrâniennes telles que la forme des membres et des vertèbres. Les Ophiacodontia ont été considérés comme le groupe le plus primitif en raison du fait que leurs membres semblent les plus reptiliens, et les Sphenacodontia étant plus avancé parce que leurs membres ressemblent le plus à ceux des thérapsides, groupe qui comprend notamment les ancêtres et parents les plus proches des mammifères. Romer et Price ont placé un autre groupe de synapsides précoces appelés Varanopidae au sein des Sphenacodontia, les considérant comme étant plus primitifs que d'autres membres du groupe comme Dimetrodon. Les chercheurs ont supposé que les varanopidés et les sphénacodontes tels Dimetrodon avaient été étroitement liés puisque que les deux groupes sont carnivores, bien que les varanopidés soient beaucoup plus petits et ressemblent davantage à des lézards[56].
La vision actuelle des relations des synapsides a été proposée par le paléontologue Robert R. Reisz (en) en 1986, dont l'étude comprend des caractéristiques principalement trouvées dans le crâne plutôt que dans le squelette postcrânien[57]. Dimetrodon est toujours considéré comme un sphénacodonte sous cette phylogénie, mais les varanopidés sont généralement considérés comme des synapsides plus basaux placés en dehors du clade Sphenacodontia, bien que certaines études ultérieures contestent ces affirmations, préférant classer cette famille au sein des reptiles diapsides[23] - [24]. Au sein des Sphenacodontia se trouve le clade Sphenacodontoidea, qui à son tour contient les Sphenacodontidae et les Therapsida. Les sphénacodontidés forment la famille contenant Dimetrodon et plusieurs autres synapsides ayant une voile sur le dos comme Sphenacodon et Secodontosaurus, tandis que les thérapsides comprennent les mammifères et leurs parents disparus ayant principalement vécu durant le Permien et le Trias[3] - [7] - [12].
Ci-dessous, un cladogramme des synapsides établi par R. J. Benson en 2012[56], à noter que la position phylogénétique des varanopidés reste contestée[23] - [24] :
| ◄ Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Le cladogramme ci-dessous montre les relations de quelques espèces de Dimetrodon selon Brink et al., (2015)[28] :
| ◄ Sphenacodontidae |
| ||||||||||||||||||||||||||||||
Paléobiologie
Fonctions des épines neurales

Les paléontologues ont proposé de nombreuses façons dont la voile aurait pu fonctionner chez l'animal. Certains des premiers à réfléchir à son utilité suggèrent qu'elle aurait pu servir de camouflage parmi les roseaux pendant que le dimétrodon attendait une proie, ou comme une véritable voile semblable à celle d'un voilier, qui aurait attrapé le vent pendant que l'animal était dans l'eau[58]. D'autres suggèrent que les longues épines neurales auraient pu stabiliser le tronc en limitant les mouvements de haut en bas, ce qui aurait permis un mouvement latéral plus efficace pendant la marche[15].
En 1940, Romer et Price proposent que la voile aurait eu une fonction thermorégulatrice, permettant aux individus de se réchauffer le corps avec la chaleur du soleil[33]. Dans les années suivantes, de nombreux modèles ont été créés pour estimer l'efficacité de la thermorégulation chez Dimetrodon. Par exemple, dans un article publiée en 1973 dans la revue Nature, les paléontologues C. D. Bramwell et P. B. Fellgett estiment qu'il aurait fallu environ une heure et demie à un individu de 200 kilos pour que sa température corporelle passe de 26 à 32 °C[59]. En 1986, Steven C. Haack conclut que le réchauffement serait plus lent qu'on ne le pense et que le processus prenait probablement quatre heures. En utilisant un modèle basé sur une variété de facteurs environnementaux et des aspects physiologiques hypothétiques de Dimetrodon, Haack affirme que la voile aurait permis à l'animal de se réchauffer plus rapidement le matin et d'atteindre une température corporelle légèrement plus élevée pendant la journée, mais qu'elle était inefficace pour libérer l'excès de chaleur et n'aurait pas permis à l'animal de conserver une température corporelle plus élevée la nuit[60]. En 1999, un groupe d'ingénieurs en mécanique crée un modèle informatique pour analyser la capacité de la voile à réguler la température corporelle au cours des différentes saisons et en conclut que la voile aurait été bénéfique pour capter et libérer la chaleur à tout moment de l'année[61].
.jpg.webp)
La plupart de ces études attribuent deux rôles thermorégulateurs à la voile de Dimetrodon : l'un comme moyen de se réchauffer rapidement le matin, et l'autre comme moyen de se rafraîchir lorsque la température corporelle devient élevée. Dimetrodon et tous les autres tétrapodes du Permien inférieur sont supposés avoir été des animaux à sang froid, s'appuyant sur le soleil pour maintenir une température corporelle élevée. En raison de sa grande taille, Dimetrodon avait une inertie thermique élevée, ce qui signifie que les changements de température corporelle s'y produisaient plus lentement que chez les animaux de plus petite taille. À mesure que les températures augmentaient le matin, les proies de petite taille de Dimetrodon pouvaient réchauffer leurs corps beaucoup plus rapidement que quelque chose de la taille de Dimetrodon. De nombreux paléontologues, dont Haack, ont proposé que la voile de Dimetrodon lui eût permis de se réchauffer rapidement le matin afin de suivre le rythme de sa proie[60]. La grande surface de la voile signifie également que la chaleur pouvait se dissiper rapidement dans l'environnement, ce qui aurait été utile si l'animal avait besoin de libérer l'excès de chaleur produit par le métabolisme ou absorbé par le soleil. Dimetrodon peut avoir incliné sa voile loin du soleil pour se rafraîchir ou restreindre le flux sanguin vers la voile pour maintenir la chaleur durant la nuit[58].
En 1986, J. Scott Turner et C. Richard Tracy proposent que l'évolution d'une voile chez le dimétrodon était liée à l'évolution du sang chaud chez les ancêtres des mammifères. Ils pensent que la voile de Dimetrodon lui permettait d'être homéothermique, en maintenant une température corporelle constante, quoique basse. Les mammifères sont également homéothermes, bien qu'ils diffèrent de Dimetrodon en ce qu'ils sont endothermiques, contrôlant leur température corporelle en interne grâce à un métabolisme accru. Turner et Tracy notent que les premiers thérapsides, un groupe plus avancé de synapsides incluant les mammifères, ont eu de longs membres qui pouvaient libérer de la chaleur d'une manière similaire à celle de la voile de Dimetrodon. L'homéothermie qui s'est développée chez des animaux comme Dimetrodon peut s'être propagée aux thérapsides par une modification de la forme du corps, qui finirait par se transformer en sang chaud chez les mammifères[62].

Des études récentes sur la voile de Dimetrodon et d'autres sphénacodontidés soutiennent l'affirmation de Haack (1986) selon laquelle la voile serait mal adaptée pour libérer de la chaleur et maintenir une température corporelle stable. La présence de voiles chez les espèces de Dimetrodon à petit corps telles que D. milleri et D. teutonis ne correspond pas à l'idée que le but de la voile était la thermorégulation car les petites voiles sont moins capables de transférer la chaleur et parce que les petits corps peuvent facilement absorber et libérer de la chaleur par eux-mêmes. De plus, les proches parents de Dimetrodon tels que Sphenacodon ont des crêtes très basses qui auraient été inutiles comme dispositifs de thermorégulation[17]. Les chercheurs pensent que la grande voile de Dimetrodon s'est développée progressivement à partir de ces crêtes plus petites, ce qui signifie que pendant la majeure partie de l'histoire évolutive de la voile, la thermorégulation n'aurait pas pu remplir une fonction importante[63].
Bien que la fonction de la voile de Dimetrodon reste incertaine, les sphénacodontidés auraient été probablement des endothermes du corps entier, caractérisés par un métabolisme énergétique élevé (tachymétabolisme) et probablement une capacité à maintenir une température corporelle élevée et stable. Cette conclusion faisait partie d'une étude à l'échelle des amniotes qui a révélé que l'endothermie tachymétabolique était répandue partout et probablement plésiomorphe pour les synapsides et les sauropsides. Pour Dimetrodon, la preuve est la taille endothermique indicative des foramens à travers lesquels le sang était acheminé vers leurs os longs et l'hypertension artérielle qui aurait été nécessaire pour fournir du sang au sommet des épines bien vascularisées soutenant la voile[64].
Les spécimens plus gros de Dimetrodon ont des voiles plus grandes par rapport à leur taille, un exemple d'allométrie positive. L'allométrie positive peut être bénéfique pour la thermorégulation car cela signifie que, à mesure que les individus grandissent, la surface augmente plus rapidement que la masse. Les animaux de plus grande taille génèrent une grande quantité de chaleur par le métabolisme, et la quantité de chaleur qui doit être dissipée de la surface du corps est nettement supérieure à celle qui doit être dissipée par les animaux de plus petite taille. La dissipation thermique efficace peut être prédite sur de nombreux animaux différents avec une seule relation entre la masse et la surface. Cependant, une étude publiée en 2010 sur l'allométrie chez Dimetrodon a trouvé une relation différente entre sa voile et sa masse corporelle : l'exposant de mise à l'échelle réel de la voile est beaucoup plus grand que l'exposant attendu chez un animal adapté à la dissipation thermique. Les chercheurs concluent que la voile de Dimetrodon grandissait à un rythme beaucoup plus rapide que nécessaire pour la thermorégulation et suggèrent que la sélection sexuelle était la principale raison de son évolution[63].
Dimorphisme sexuel
Dimetrodon peut avoir été sexuellement dimorphe, ce qui signifie que les mâles et les femelles ont eu des tailles corporelles légèrement différentes. Certains spécimens de Dimetrodon ont été supposés être des mâles parce qu'ils ont des os plus épais, des voiles plus grandes, des crânes plus longs et des « marches » maxillaires plus prononcées que d'autres. Sur la base de ces différences, les squelettes montés du musée américain d'histoire naturelle (AMNH 4636) et du musée Field peuvent être des mâles et les squelettes du musée de la nature et des sciences de Denver (MCZ 1347) et du musée d'histoire naturelle de l'université du Michigan (en) peuvent être des femelles[33].


Paléoécologie
Environnement

Les fossiles de Dimetrodon sont connus des États-Unis (plus précisément dans les états du Texas, d'Oklahoma, du Nouveau-Mexique, d'Arizona, d'Utah et de l'Ohio), du Canada et d'Allemagne, des régions qui ont fait partie du supercontinent laurasien durant le Permien inférieur. Aux États-Unis, presque tout le matériel attribué à Dimetrodon provient des lits rouges du Texas et de l'Oklahoma[65] - [66]. La plupart des découvertes de fossiles font partie d'écosystèmes de plaine qui, au cours du Permien, auraient été de vastes zones humides. En particulier, les lits rouges du Texas sont une zone de grande diversité de tétrapodes fossiles. En plus de Dimetrodon, les tétrapodes les plus courants dans les lits rouges et dans les dépôts du Permien inférieur du Sud-Ouest des États-Unis sont les amphibiens Archeria, Diplocaulus, Eryops et Trimerorhachis, le reptiliomorphe Seymouria, le sauropside Captorhinus et les synapsides Ophiacodon et Edaphosaurus. Ces tétrapodes constituaient un groupe d'animaux que le paléontologue Everett C. Olson surnomme la « chronofaune Permo-Carbonifère », une faune qui a dominé l'écosystème continental laurasien pendant plusieurs millions d'années[67]. Sur la base de la géologie de gisements comparables à ceux des lits rouges, les chercheurs pensent que la faune aurait habité un écosystème deltaïque de plaine bien végétalisé[68].
Réseau trophique

Olson a fait de nombreuses inférences sur la paléoécologie des lits rouges du Texas et de l'Oklahoma et le rôle de Dimetrodon dans son écosystème. Il propose plusieurs principaux types d'écosystèmes dans lesquels vivaient les premiers tétrapodes. Dimetrodon appartenait à l'écosystème le plus primitif, qui s'est développé à partir des réseaux trophiques aquatiques. Les plantes aquatiques y étaient les principales productrices et étaient largement consommés par les poissons et les invertébrés marins. La plupart des vertébrés terrestres se nourrissaient de ces consommateurs primaires aquatiques. Dimetrodon était probablement le superprédateur de l'écosystème des lits rouges, se nourrissant d'une variété d'organismes tels que le chondrichtyen Xenacanthus, les amphibiens aquatiques Trimerorhachis et Diplocaulus, et les tétrapodes terrestres tels que Seymouria et Acheloma. Les insectes sont connus des premiers lits rouges du Permien et étaient probablement impliqués dans une certaine mesure dans le même réseau trophique que Dimetrodon, nourrissant de petits reptiles comme Captorhinus. L'assemblage des lits rouges comprenait également certains des premiers grands herbivores terrestres comme Edaphosaurus et Diadectes. Se nourrissant principalement de plantes terrestres, ces herbivores ne tiraient pas leur énergie des réseaux trophiques aquatiques. Selon Olson, le meilleur analogue moderne de l'écosystème ayant été habité par Dimetrodon sont les Everglades, situé en Floride[68]. Le mode de vie exact du dimétrodon a longtemps été controversé, mais l'anatomie osseuse de l'animal soutient un mode de vie terrestre, ce qui implique qu'il se serait nourri principalement sur terre, sur les berges ou dans des eaux très peu profondes[69]. Il existe également des preuves que Dimetrodon se nourrissait de Diplocaulus estivant pendant les périodes de sécheresse, avec trois Diplocaulus juvéniles partiellement dévorés dans un terrier avec huit marques de dents portant d'un dimétrodon qui les a déterrés et tués[70]. La formation de Orby Head (en) au Canada, lieu d'où D. borealis est connu et datant d'un âge similaire aux lits rouges, conserve des fossiles de tétrapodes comparables ainsi que divers plantes palynomorphes. Des traces de pas fossiles de vertébrés et d'invertébrés ont été également identifiés dans la zone[28].
La seule espèce du genre Dimetrodon ayant été découverte en dehors du États-Unis est D. teutonis, connu d'Allemagne. Ses restes ont été trouvés dans la formation de Tambach, située dans un site fossilifère appelé la localité de Bromacker. L'assemblage de tétrapodes du Permien inférieur de Bromacker est inhabituel en ce sens qu'il existe peu de synapsides de grande taille jouant le rôle de superprédateurs. La taille estimé de D. teutonis est de 1,7 mètre de longueur, trop petit pour s'attaquer aux grands herbivores de la famille des diadectidés qui sont abondants dans la zone de Bromacker. Il se serait plus probablement attaqué aux petits vertébrés et aux insectes. Seuls trois fossiles peuvent être attribués à de grands prédateurs, et on pense qu'ils étaient soit de grands varanopidés, soit de petits sphénacodontes, qui pourraient tous deux s'en prendre à D. teutonis lors des chasses. Contrairement aux lits rouges deltaïques des basses terres du Texas, les gisements de Bromacker représentaient un environnement de hautes terres sans espèces aquatiques. Il est possible que les carnivores de grande taille ne fassent pas partie de l'assemblage de Bromacker car ils dépendaient de grands amphibiens aquatiques pour se nourrir[1].
Dimétrodon dans la culture

Le dimétrodon est l'un des animaux préhistoriques les plus célèbre du grand public pour son aspect facilement différentiable des grands sauropsides connus, notamment pour sa voile dorsale qui fait sa popularité, comme celle de Spinosaurus ou d'Ouranosaurus. Cependant, l'animal étant souvent mentionné dans divers lieux public et présenté au côtés de dinosaures. Cela a conduit à l'idée majoritairement répandue que Dimetrodon serait l'un d'eux, alors qu'il est plus proche des mammifères que de n'importe quel reptile. Il est ainsi le synapside non mammifère le plus connu du grand public et on le retrouve dans plusieurs éléments de la culture populaire, notamment dans des jouets, livres et même dans divers médias incluant dessins-animés, documentaires et films[41].
Notes et références
Notes
- Les thériodontes forment un groupe avancée de thérapsides comprenant notamment les gorgonopsiens, les thérocéphales ainsi que les cynodontes, ces derniers étant les ancêtres des mammifères[41].
Références
- (en + fr) David S. Berman, Robert R. Reisz, Thomas Martens et Amy C. Henrici, « A new species of Dimetrodon (Synapsida: Sphenacodontidae) from the Lower Permian of Germany records first occurrence of genus outside of North America », Canadian Journal of Earth Sciences, vol. 38, no 5, , p. 803-812 (DOI 10.1139/cjes-38-5-803, Bibcode 2001CaJES..38..803B, S2CID 52268104, lire en ligne [PDF])
- (en) « The Kungurian Age » [archive du ], sur Palaeos (consulté le )
- (en) J. Fröbisch, R. R. Schoch, J. Müller, T. Schindler et D. Schweiss, « A new basal sphenacodontid synapsid from the Late Carboniferous of the Saar-Nahe Basin, Germany », Acta Palaeontologica Polonica, vol. 56, no 1, , p. 113–120 (DOI 10.4202/app.2010.0039
 , S2CID 45410472, lire en ligne [PDF])
, S2CID 45410472, lire en ligne [PDF]) - (en) E. C. Olson et J. R. Beerbower, « The San Angelo Formation, Permian of Texas, and its vertebrates », The Journal of Geology, vol. 61, no 5, , p. 389–423 (DOI 10.1086/626109, Bibcode 1953JG.....61..389O, S2CID 128681671)
- (en) E. C. Olson, « Parallelism in the evolution of the Permian reptilian faunas of the Old and New Worlds », Fieldiana, vol. 37, no 13, , p. 385–401 (lire en ligne)
- (en) C. W. Sternberg, « The skeleton of an immature pelycosaur, Dimetrodon cf. grandis, from the Permian of Texas », Journal of Paleontology, vol. 16, no 4, , p. 485–486 (JSTOR 1298848)
- (en) M. Laurin et R. R. Reisz, « Synapsida: mammals and their extinct relatives », sur Tree of Life Web Project, (consulté le )
- (en) G. Baur et E. C. Case, « The history of the Pelycosauria, with a description of the genus Dimetrodon, Cope », Transactions of the American Philosophical Society, vol. 20, no 1, , p. 5-62 (DOI 10.2307/1005488, JSTOR 1005488, hdl 2027/uc1.32106020416696 , S2CID 133464941)
- (en) D. H. Tanke, K. Carpenter et M. W. Skrepnick, Mesozoic Vertebrate Life, Bloomington, Indiana University Press, , 84-89 p. (ISBN 978-0-253-33907-2), « A kerf-and-drill model of tyrannosaur tooth serrations »
- (en) Kirstin S. Brink et Robert R. Reisz, « Hidden dental diversity in the oldest terrestrial apex predator Dimetrodon », Nature Communications, vol. 5, no 1, , p. 3269 (PMID 24509889, DOI 10.1038/ncomms4269 , Bibcode 2014NatCo...5.3269B, S2CID 10705383)
- (en) T. S. Kemp, « The origin and early radiation of the therapsid mammal‐like reptiles: a palaeobiological hypothesis », Journal of Evolutionary Biology, vol. 19, no 4, , p. 1231-1247 (PMID 16780524, DOI 10.1111/j.1420-9101.2005.01076.x , S2CID 3184629).
- (en) M. Laurin et R. R. Reisz, « Autapomorphies of the main clades of synapsids », sur Tree of Life Web Project, (consulté le )
- (en) A. S. Romer, « Notes on the Permo-Carboniferous reptile Dimetrodon », The Journal of Geology, vol. 35, no 8, , p. 673–689 (DOI 10.1086/623462, JSTOR 30060393, Bibcode 1927JG.....35..673R, S2CID 140679339)
- (en) E. A. Rega, K. Noriega, S. S. Sumida, A. Huttenlocker, A. Lee et B. Kennedy, « Healed Fractures in the Neural Spines of an Associated Skeleton of Dimetrodon: Implications for Dorsal Sail Morphology and Function », Fieldiana Life and Earth Sciences, vol. 5, , p. 104–111 (DOI 10.3158/2158-5520-5.1.104, S2CID 108887164)
- (en) E. Rega, S. Sumida, Noriega K., Pell C. et Lee A., « Evidence-based paleopathology I: Ontogenetic and functional implications of dorsal sails in Dimetrodon », Journal of Vertebrate Paleontology, vol. 25, no S3, , p. 103A (DOI 10.1080/02724634.2005.10009942, S2CID 220413556)
- (en) S. Sumida, E. Rega et K. Noriega, « Evidence-based paleopathology II: Impact on phylogenetic analysis of the genus Dimetrodon », Journal of Vertebrate Paleontology, vol. 25, no S3, , p. 120A (DOI 10.1080/02724634.2005.10009942, S2CID 220413556)
- (en) A. K. Huttenlocker, E. Rega et S. S. Sumida, « Comparative anatomy and osteohistology of hyperelongate neural spines in the sphenacodontids Sphenacodon and Dimetrodon (Amniota: Synapsida) », Journal of Morphology, vol. 271, no 12, , p. 1407-1421 (PMID 20886514, DOI 10.1002/jmor.10876, S2CID 40899700)
- (en) Elizabeth A. Rega, Ken Noriega, Stuart S. Sumida et Andrew Lee, « Histological analysis of traumatic injury to multiple neural spines of an associated skeleton of Dimetrodon: Implications for healing response, dorsal sail morphology and age-at-death in a Lower Permian synapsid », Integrative and Comparative Biology, vol. 44, , p. 628
- (en) Petr Konstantinovich Tchudinov, « New Facts about the Fauna of the Upper Permian of the USSR », The Journal of Geology, vol. 73, no 1, , p. 117–30 (DOI 10.1086/627048, Bibcode 1965JG.....73..117C, S2CID 129428120)
- (en) J. Botha-Brink et S. P. Modesto, « A mixed-age classed 'pelycosaur' aggregation from South Africa: earliest evidence of parental care in amniotes? », Proceedings of the Royal Society B, vol. 274, no 1627, , p. 2829–2834 (PMID 17848370, PMCID 2288685, DOI 10.1098/rspb.2007.0803, lire en ligne [PDF])
- (en) G. Niedźwiedzki et M. Bojanowski, « A Supposed Eupelycosaur Body Impression from the Early Permian of the Intra-Sudetic Basin, Poland », Ichnos, vol. 19, no 3, , p. 150–155 (DOI 10.1080/10420940.2012.702549, S2CID 129567176)
- (en) Frederik Spindler, Ralf Werneburg, Joerg W. Schneider, Ludwig Luthardt, Volker Annacker et Ronny Rößler, « First arboreal 'pelycosaurs' (Synapsida: Varanopidae) from the early Permian Chemnitz Fossil Lagerstätte, SE Germany, with a review of varanopid phylogeny », PalZ, vol. 92, no 2, , p. 315–364 (DOI 10.1007/s12542-018-0405-9, S2CID 133846070)
- (en) David P. Ford et Roger B. J. Benson, « A redescription of Orovenator mayorum (Sauropsida, Diapsida) using high-resolution μCT, and the consequences for early amniote phylogeny », Papers in Palaeontology, vol. 5, no 2, , p. 197–239 (ISSN 2056-2802, DOI 10.1002/spp2.1236, S2CID 92485505, lire en ligne [PDF])
- (en) Sean P. Modesto, « Rooting about reptile relationships », Nature Ecology & Evolution, vol. 4, no 1, , p. 10–11 (ISSN 2397-334X, PMID 31900449, DOI 10.1038/s41559-019-1074-0, S2CID 209672518)
- (en) Joseph Leidy, « Remarks on a fragment of a jaw of an extinct Saurian, from Prince Edward’s Island », Proceedings of the Academy of Natural Sciences of Philadelphia, vol. 6, , p. 404
- (en) Joseph Leidy, « On Bathygnathus borealis, an extinct saurian of the New Red sandstone of Prince Edward’s Island », Journal of the Academy of Natural Sciences of Philadelphia, vol. 2, , p. 327-330
- (en + fr) D. A. E. Spalding, Vertebrate Fossils and the Evolution of Scientific Concepts, Taylor & Francis US, , 245-254 p. (ISBN 2881249965), « Bathygnathus, Canada's first "dinosaur" »
- (en + fr) Kirstin S. Brink, Hillary C. Maddin, David C. Evans, Robert R. Reisz et Hans-Dieter Sues, « Re-evaluation of the historic Canadian fossil Bathygnathus borealisfrom the Early Permian of Prince Edward Island », Canadian Journal of Earth Sciences, vol. 52, no 12, , p. 1109–1120 (DOI 10.1139/cjes-2015-0100 , Bibcode 2015CaJES..52.1109B, S2CID 130774660, lire en ligne [PDF])
- (en) W. D. Matthew, « Review of Case's Revision of the Pelycosauria of North America », Science, vol. 27, no 699, , p. 816-818 (DOI 10.1126/science.27.699.816, Bibcode 1908Sci....27..816M, S2CID 178711873)
- (en) E.C. Case, Revision of the Pelycosauria of North America, Washington, D.C., Carnegie Institution of Washington, , 176 p. (lire en ligne)
- (en) E. D. Cope, « Descriptions of extinct Batrachia and Reptilia from the Permian formation of Texas », Proceedings of the American Philosophical Society, vol. 17, no 101, , p. 505-530 (DOI 10.5962/bhl.title.104569 , JSTOR 982652, S2CID 130656729, lire en ligne)
- (en) E. D. Cope, « Descriptions of extinct Vertebrata from the Permian and Triassic Formations of the United States », Proceedings of the American Philosophical Society, vol. 17, no 100, , p. 182-193 (JSTOR 982295, S2CID 135111418)
- (en) A. S. Romer et Price, « Review of the Pelycosauria », Geological Society of America Special Paper, vol. 28, , p. 1-538 (DOI 10.1130/spe28-p1, S2CID 86826161)
- (en) P. P. Vaughn, « Comparison of the Early Permian vertebrate faunas of the Four Corners region and north-central Texas », Contributions in Science, Los Angeles County Museum of Natural History, vol. 105, , p. 1-13 (lire en ligne [archive du ] [PDF], consulté le )
- (en) P.P. Vaughn, « Early Permian vertebrates from southern New Mexico and their paleozoogeographic significance », Contributions in Science, Los Angeles County Museum of Natural History, vol. 166, , p. 1-22 (lire en ligne [archive du ] [PDF], consulté le )
- (en) E. C. Olson, « Permo-Carboniferous paleoecology and morphotypic series », American Zoologist, vol. 15, no 2, , p. 371-389 (DOI 10.1093/icb/15.2.371 , S2CID 85014773)
- (en) D. S. Berman, « A new species of Dimetrodon (Reptilia, Pelycosauria) from a non-deltaic facies in the Lower Permian of north-central New Mexico », Journal of Paleontology, vol. 51, no 1, , p. 108-115 (JSTOR 1303466, S2CID 133330419)
- (en) K. Madalena, S. Sumida, K. Zeigler et E. Rega, « A new record of the Early Permian pelycosaurian-grade synapsid Dimetrodon (Eupelycosauria: Sphenacodontidae) from the Lower Cutler Group (Early Permian) of Jemez Pueblo, north-central New Mexico », Journal of Vertebrate Paleontology, vol. 27, no 3, Suppl, , p. 110A (DOI 10.1080/02724634.2007.10010458)
- (en) P. P. Vaughn, Vertebrates from the Cutler Group of Monument Valley and vicinity, vol. 24, New Mexico Geological Society, coll. « New Mexico Geological Society Guidebook », , 99-105 p., PDF (lire en ligne)
- (en) Richard Owen, « Evidences of theriodonts in Permian deposits elsewhere than in South Africa », Quarterly Journal of the Geological Society, vol. 32, , p. 352-363 (DOI 10.1144/GSL.JGS.1876.032.01-04.41, S2CID 140711697)
- (en) K. D. Angielczyk, « Dimetrodon is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution », Evolution: Education and Outreach, vol. 2, no 2, , p. 257-271 (DOI 10.1007/s12052-009-0117-4 , S2CID 24110810)
- (en) E. D. Cope, « On some new Batrachia and Reptilia from the Permian Red Beds of Texas », Bulletin of the United States Geological Survey, vol. 6, , p. 79-82
- (en) E. C. Case, « The osteology of Embolophorus dollovianus, Cope, with an attempted restoration », The Journal of Geology, vol. 11, no 1, , p. 1–28 (DOI 10.1086/621055 , JSTOR 30056735, Bibcode 1903JG.....11....1C, S2CID 128537086)
- (en) E. C. Case, « The osteology of the skull of the pelycosaurian genus, Dimetrodon », The Journal of Geology, vol. 12, no 4, , p. 304-311 (DOI 10.1086/621157 , JSTOR 30055825, Bibcode 1904JG.....12..304C, lire en ligne)
- (en) C. W. Gilmore, « A mounted skeleton of Dimetrodon gigas in the United States National Museum, with notes on the skeletal anatomy », Proceedings of the U.S. National Museum, vol. 56, no 2300, , p. 525-539 (DOI 10.5479/si.00963801.56-2300.525 , S2CID 40833898, lire en ligne [PDF])
- (en) A. C. Henrici, Zeigler, K.E., S. G. Lucas, A. B. Heckert, L. F. Rinehart et Zeigler K. E., The Nonmarine Permian, vol. 30, Albuquerque, New Mexico Museum of Natural History and Science, coll. « New Mexico Museum of Natural History and Science Bulletin », , 106-110 p. (lire en ligne), « The carpus and tarsus of the Early Permian synapsid Sphenacodon ferox (Eupelycosauria: Sphenacodontidae) »
- (en) A. S. Romer, « New genera and species of pelycosaurian reptiles », Proceedings of the New England Zoological Club, vol. 16, , p. 89–97 (lire en ligne [PDF])
- (en) E. C. Olson, « Late Permian terrestrial vertebrates, USA and USSR », Transactions of the American Philosophical Society, vol. 52, no 2, , p. 1–224 (DOI 10.2307/1005904, JSTOR 1005904)
- (en) B. Battail, « A comparison of Late Permian Gondwanan and Laurasian amniote faunas », Journal of African Earth Sciences, vol. 31, no 1, , p. 165–174 (DOI 10.1016/S0899-5362(00)00081-6, Bibcode 2000JAfES..31..165B, S2CID 140664898)
- (en) E. D. Cope, « The Theromorphous Reptilia », The American Naturalist, vol. 12, no 12, , p. 829-830 (DOI 10.1086/272251 )
- (en) E. D. Cope, « Second contribution to the history of the vertebrata of the Permian Formation of Texas », Proceedings of the American Philosophical Society, vol. 19, no 107, , p. 38-58 (JSTOR 982605, lire en ligne [PDF])
- (en) E. D. Cope, « The long-spined Theromorpha of the Permian Epoch », The American Naturalist, vol. 20, no 6, , p. 544-545 (DOI 10.1086/274275 )
- (en) E. D. Cope, « Third contribution to the history of the vertebrata of the Permian Formation of Texas », Proceedings of the American Philosophical Society, vol. 20, no 112, , p. 447-461 (JSTOR 982692)
- (en) S. W. Williston, « The osteology of some American Permian vertebrates, II », Contribution from the Walker Museum, vol. 1, , p. 165-192
- R. R. Reisz, D. S. Berman et D. Scott, « The cranial anatomy and relationships of Secodontosaurus, an unusual mammal-like reptile (Synapsida: Sphenacodontidae) from the early Permian of Texas », Zoological Journal of the Linnean Society, vol. 104, no 2, , p. 127-184 (DOI 10.1111/j.1096-3642.1992.tb00920.x, S2CID 56425294)
- (en) R. J. Benson, « Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies », Journal of Systematic Palaeontology, vol. 10, no 2, , p. 601-624 (DOI 10.1080/14772019.2011.631042, S2CID 84706899)
- (de) R. R. Reisz, « Pelycosauria », dans R. R. Reisz, Handbuch der Paläoherpetologie, vol. 17A, Sues, H.-D., , 1–102 p. (ISBN 978-3-89937-032-4)
- (en) G. A. Florides, Kalogirou, S. A. Tassou et L. Wrobel, « Natural environment and thermal behaviour of Dimetrodon limbatus », Journal of Thermal Biology, vol. 26, no 1, , p. 15-20 (PMID 11070340, DOI 10.1016/S0306-4565(00)00019-X, S2CID 22905981)
- (en) C. D. Bramwell et P. B. Fellgett, « Thermal regulation in sail lizards », Nature, vol. 242, no 5394, , p. 203-205 (DOI 10.1038/242203a0, Bibcode 1973Natur.242..203B, S2CID 4159825)
- (en) S. C. Haack, « A thermal model of the sailback pelycosaur », Paleobiology, vol. 12, no 4, , p. 450-458 (DOI 10.1017/S009483730000316X, S2CID 124339088)
- (en) G. A. Florides, L. C. Wrobel, S. A. Kalogirou et S. A. Tassou, « A thermal model for reptiles and pelycosaurs », Journal of Thermal Biology, vol. 24, no 1, , p. 1-13 (DOI 10.1016/S0306-4565(98)00032-1, S2CID 83677371)
- (en) J. S. Turner et C. R. Tracy, « Body size, homeothermy and the control of heat exchange in mammal-like reptiles », dans N. III. Hotton, P. D. Maclean, J. J. Roth, E. C. Roth, The Ecology and Biology of Mammal-like Reptiles, Washington, D.C., , 326 p., PDF (lire en ligne)
- (en) J. L. Tomkins, N. R. LeBas, M. P. Witton, D. M. Martill et S. Humphries, « Positive allometry and the prehistory of sexual selection », The American Naturalist, vol. 176, no 2, , p. 141-148 (PMID 20565262, DOI 10.1086/653001, S2CID 36207, lire en ligne [archive du ] [PDF])
- (en) Gordon Grigg, Julia Nowack, José Eduardo Pereira Wilken Bicudo, Naresh Chandra Bal, Holly N. Woodward et Roger S. Seymour, « Whole‐body endothermy: ancient, homologous and widespread among the ancestors of mammals, birds and crocodylians », Biological Reviews, vol. 97, no 2, , p. 766–801 (ISSN 1464-7931, PMID 34894040, PMCID 9300183, DOI 10.1111/brv.12822 )
- (en) S. G. Lucas, J. A. Spielmann, L. F. Rinehart et T. Martens, Dimetrodon (Amniota: Synapsida: Sphenacodontidae) from the Lower Permian Abo Formation, Socorro County, New Mexico, vol. 60, New Mexico Geological Society, coll. « New Mexico Geological Society Guidebook », , 281–284 p., PDF (lire en ligne)
- (en) W. John Nelson, Robert W. Hook et Dan S. Chaney, « Lithostratigraphy of the Lower Permian (Leonardian) Clear Fork Formation of North-Central Texas », New Mexico Museum of Natural History and Science, , p. 286-311 (lire en ligne)
- (en) C. Sullivan, R. R. Reisz et W. J. May, « Large dissorophoid skeletal elements from the Lower Permian Richards Spur fissures, Oklahoma, and their paleoecological implications », Journal of Vertebrate Paleontology, vol. 20, no 3, , p. 456-461 (DOI 10.1671/0272-4634(2000)020[0456:LDSEFT]2.0.CO;2, JSTOR 4524117, S2CID 140709673)
- (en) E. C. Olson, « Community evolution and the origin of mammals », Ecology, vol. 47, no 2, , p. 291-302 (DOI 10.2307/1933776, JSTOR 1933776, S2CID 86728985)
- (en) A. Kriloff, D. Germain, A. Canoville, P. Vincent, M. Sache et M. Laurin, « Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference », Journal of Evolutionary Biology, vol. 21, no 3, , p. 807-826 (PMID 18312321, DOI 10.1111/j.1420-9101.2008.01512.x , S2CID 6102313)
- (en) « Finned Monster Chomped Heads off Ancient Amphibians », sur Live Science,
Citations originales
- « The utility is difficult to imagine. Unless the animal had aquatic habits, and swam on its back, the crest or fin must have been in the way of active movements [...] The limbs are not long enough nor the claws acute enough to demonstrate arboreal habits, as in the existing genus Basilicus, where a similar crest exists. »
Voir aussi
Articles connexes
Vidéos
- (en) [vidéo] Dimetrodon: Our Most Unlikely Ancestor sur YouTube.
Liens externes
- (en) « Dimetrodon », sur Palaeos
- (en) « Introduction to the Pelycosaurs », sur le site web de musée de paléontologie de l'université de Californie.
- Ressources relatives au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :