Anapsida
Les anapsides (Anapsida) forment une sous-classe de vertébrés tétrapodes. Le terme anapside leur est attribué à cause de l'absence de fentes temporales au niveau du crâne et par opposition aux animaux synapsides (1 fosse) et diapsides (2 fosses). On considérait auparavant que les tortues, l'ordre des Testudines, appartenait à cette sous-classe. Aujourd'hui certains auteurs considèrent les testudines comme des diapsides ayant perdu leurs fosses temporales[1].
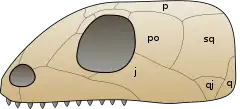
j: jugal, p: pariétal, po: postorbitaire, q: carré,
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Clade | Amniota |
| Clade | Sauropsida |
| Classe | Reptilia |
Taxons de rang inférieur
- † Mesosauria
- † Millerettidae
- † Nyctiphruretidae
- † Pareiasauridae
- † Procolophonidae
- Testudines ?
- Sauropsides
- Diapsides
- Anapsides
Ancrage des tortues au sens des anapsides
Alors que l'on parlait traditionnellement de reptiles anapsides comme s'il s'agissait d'un groupe monophylétique, il a été suggéré que plusieurs groupes de reptiles ayant un crâne d'anapside pourraient être liés de façon lointaine. Les scientifiques discutent encore de la relation exacte entre les reptiles basaux (originaux) apparus pour la première fois dans le Carbonifère tardif et les divers reptiles du Permien qui avaient un crâne anapside et les tortues. Cependant, il a été suggéré par la suite que le crâne de tortue ressemblant à une anapside était dû à la réversion plutôt qu'à une descente anapside. La majorité des paléontologues modernes croient que les Testudines sont des descendants de reptiles diapsides ayant perdu leur fenêtre temporale. Des études phylogénétiques morphologiques plus récentes dans cet esprit ont placé les tortues fermement dans les diapsides[2] - [3] - [4], certaines tortues locales constituant un groupe-frère pour les archosaures existants[5] - [6] ou, plus communément, chez les Lepidosauromorpha[7] - [8] - [9] - [10] - [11].
Position phylogénetique

Toutes les études moléculaire ont fortement confirmé le placement des tortues chez les diapsides, et plus précisément chez les Archosauria (groupe incluant les Dinosauria) ou en faire un groupe-frère[12] - [13] - [14] - [15] - [16] - [17]. Cependant, l'une des études moléculaires les plus récentes, publiée le 23 février 2012, suggère que les tortues sont des diapsides lépidosauromorphes, plus étroitement apparentées aux Lepidosauria, Lacerticia ou encore les Serpentes[18].
La réanalyse de phylogénies antérieures suggère qu’elles classent les tortues comme des anapsides, à la fois parce qu’elles ont assumé cette classification (la plupart d’entre elles étudient le type de tortues anapsides) et parce qu’elles n’ont pas échantillonné suffisamment de taxons existants et fossiles pour construire le cladogramme. Il est suggéré que Testudines se soit écarté d'autres diapsides, il y a entre 200 et 279 millions d'années, bien que le débat soit loin d'être réglé[7] - [13] - [19]. Bien que procolophonidés ait réussi à survivre dans le Trias, la plupart des autres reptiles dotés de crânes anapsides, y compris les millerettidés, nycteroleteridés, et pareiasaures, se sont éteints lors de l'extinction Permien-Trias.
Malgré les études moléculaires, certaines preuves contredisent leur classification en tant que diapsides. Tous les diapsides connus excrètent de l'acide urique sous forme de déchets azotés, et il n'y a aucun cas connu de diapside revenant à l'excrétion d'urée, même lorsqu'ils reviennent à un mode de vie semi-aquatique. Les crocodiliens, par exemple, sont toujours uricotéliques, bien qu’ils soient également en partie ammonotéliques, ce qui signifie qu’ils excrètent une partie de leurs déchets sous forme d’ammoniac. L'uréotélisme semble être la condition ancestrale chez les amniotes primitifs et il est conservé par les mammifères, qui ont probablement hérité de l'uréotélisme de leurs ancêtres synapsides. L'ulotélisme suggérerait donc que les tortues étaient plus susceptibles d'être des anapsides que des diapsides. Le seul chélonien uricotélique connu est la tortue du désert, qui l'a probablement développée récemment pour s'adapter aux habitats du désert. Certains mammifères désertique sont aussi des uricotéliciens. Ainsi, puisque pratiquement tous les mammifères connus sont des uréotéliques, l'adaptation uricotélique est probablement le résultat de la convergence entre les espèces du désert. Par conséquent, les tortues devraient être le seul cas connu de reptile uricotelique redevenant uréotélique.
Notes
- (en) M. deBraga & O. Rieppel, « Reptile phylogeny and the interrelationships of turtles », Zoological Journal of the Linnean Society, vol. 120, no 3, , p. 281-354 (résumé)
- M. deBraga et O. Rieppel, « Reptile phylogeny and the interrelationships of turtles », Zoological Journal of the Linnean Society, vol. 120, no 3, , p. 281–354 (DOI 10.1111/j.1096-3642.1997.tb01280.x, lire en ligne)
- Linda A. Tsuji et Johannes Muller, « Assembling the history of the Parareptilia: phylogeny, diversification, and a new definition of the clade », Fossil Record, vol. 12, no 1, , p. 71–81 (DOI 10.1002/mmng.200800011)
- Marcello Ruta, Juan C. Cisneros, Torsten Liebrect, Linda A. Tsuji et Johannes Muller, « Amniotes through major biological crises: faunal turnover among Parareptiles and the end-Permian mass extinction », Palaeontology, vol. 54, no 5, , p. 1117–1137 (DOI 10.1111/j.1475-4983.2011.01051.x)
- Susan E. Evans, « An early kuehneosaurid reptile (Reptilia: Diapsida) from the Early Triassic of Poland », Paleontologica Polonica, vol. 65, , p. 145–178 (lire en ligne).
- Magdalena Borsuk−Białynicka; and Susan E. Evans, « A long−necked archosauromorph from the Early Triassic of Poland », Paleontologica Polonica, vol. 65, , p. 203–234 (lire en ligne)
- Rieppel O, DeBraga M, « Turtles as diapsid reptiles », Nature, vol. 384, no 6608, , p. 453–5 (DOI 10.1038/384453a0, Bibcode 1996Natur.384..453R)
- Chun Li, Xiao-Chun Wu, Olivier Rieppel, Li-Ting Wang et Li-Jun Zhao, « An ancestral turtle from the Late Triassic of southwestern China », Nature, vol. 456, no 7221, , p. 497–501 (PMID 19037315, DOI 10.1038/nature07533, Bibcode 2008Natur.456..497L)
- Constanze Bickelmann, Johannes Müller and Robert R. Reisz, « The enigmatic diapsid Acerosodontosaurus piveteaui (Reptilia: Neodiapsida) from the Upper Permian of Madagascar and the paraphyly of younginiform reptiles », Canadian Journal of Earth Sciences, vol. 49, no 9, , p. 651–661 (DOI 10.1139/E09-038, Bibcode 2009CaJES..46..651S)
- Robert R. Reisz, Sean P. Modesto and Diane M. Scott, « A new Early Permian reptile and its significance in early diapsid evolution », Proceedings of the Royal Society B, vol. 278, no 1725, , p. 3731–3737 (PMID 21525061, PMCID 3203498, DOI 10.1098/rspb.2011.0439)
- Chun Li, Olivier Rieppel, Xiao-Chun Wu, Li-Jun Zhao et Li-Ting Wang, « A new Triassic marine reptile from southwestern China », Journal of Vertebrate Paleontology, vol. 31, no 2, , p. 303–312 (DOI 10.1080/02724634.2011.550368, lire en ligne)
- Hideyuki Mannena et Li, Steven S. -L., « Molecular evidence for a clade of turtles », Molecular Phylogenetics and Evolution, vol. 13, no 1, , p. 144–148 (PMID 10508547, DOI 10.1006/mpev.1999.0640)
- R. Zardoya et A. Meyer, « Complete mitochondrial genome suggests diapsid affinities of turtles », Proc Natl Acad Sci U S A, vol. 95, no 24, , p. 14226–14231 (ISSN 0027-8424, PMID 9826682, PMCID 24355, DOI 10.1073/pnas.95.24.14226, Bibcode 1998PNAS...9514226Z)
- N. Iwabe, Hara, Y., Kumazawa, Y., Shibamoto, K., Saito, Y., Miyata, T. et Katoh, K., « Sister group relationship of turtles to the bird-crocodilian clade revealed by nuclear DNA-coded proteins », Molecular Biology and Evolution, vol. 22, no 4, , p. 810–813 (PMID 15625185, DOI 10.1093/molbev/msi075)
- Jonas Roos, Aggarwal, Ramesh K. et Janke, Axel, « Extended mitogenomic phylogenetic analyses yield new insight into crocodylian evolution and their survival of the Cretaceous–Tertiary boundary », Molecular Phylogenetics and Evolution, vol. 45, no 2, , p. 663–673 (PMID 17719245, DOI 10.1016/j.ympev.2007.06.018)
- Y. Katsu, Braun, E. L., Guillette, L. J. Jr. et Iguchi, T., « From reptilian phylogenomics to reptilian genomes: analyses of c-Jun and DJ-1 proto-oncogenes », Cytogenetic and Genome Research, vol. 127, nos 2–4, , p. 79–93 (PMID 20234127, DOI 10.1159/000297715)
- N. G. Crawford, B. C. Faircloth, J. E. McCormack, R. T. Brumfield, K. Winker et T. C. Glenn, « More than 1000 ultraconserved elements provide evidence that turtles are the sister group of archosaurs », Biology Letters, vol. 8, no 5, , p. 783–6 (PMID 22593086, PMCID 3440978, DOI 10.1098/rsbl.2012.0331)
- Tyler R. Lyson, Erik A. Sperling, Alysha M. Heimberg, Jacques A. Gauthier, Benjamin L. King et Kevin J. Peterson, « MicroRNAs support a turtle + lizard clade », Biology Letters, vol. 8, no 1, , p. 104–107 (PMID 21775315, PMCID 3259949, DOI 10.1098/rsbl.2011.0477)
- M. J. Benton, Vertebrate Paleontology, Londres, Blackwell Science Ltd, , 2e éd., 452 p. (ISBN 978-0-632-05614-9), 3rd ed. 2004 (ISBN 0-632-05637-1)
Articles connexes
Liens externes
- (en) Référence Tree of Life Web Project : Anapsida
- (en) Référence Paleobiology Database : Anapsida Williston 1917
- (fr+en) Référence ITIS : Anapsida Non valide
- (fr+en) Référence ITIS : Reptilia Laurenti, 1768
- (en) Référence Animal Diversity Web : Anapsida
- (en) Référence NCBI : Testudines (taxons inclus)