Dendrobatidae
Les Dendrobatidae, communément appelés dendrobates, sont une famille d'amphibiens[1] qui sont originaires de l'Amérique centrale et du Sud. Ces espèces de grenouilles de petite taille sont diurnes et ont souvent des corps de couleur vive. Tous les dendrobates sauvages sont toxiques, bien que leur toxicité varie considérablement d'une espèce à l'autre et d'une population à une autre. Ces amphibiens sont appelés « poison-dart frogs » en anglais (grenouilles à flèches empoisonnées) en raison de l'utilisation faite par les Amérindiens de leurs sécrétions toxiques pour empoisonner les extrémités de leurs flèches de sarbacane. Cependant, seulement quatre espèces provenant du genre Phyllobates, hautement toxique, ont été documentées comme étant utilisées à cet effet [2] - [3]. Elles font l'objet d'un commerce notamment en France en tant que nouveaux animaux de compagnie. De nombreuses espèces sont menacées.

- Phyllobatidae Fitzinger, 1843
- Eubaphidae Bonaparte, 1850
- Hylaplesidae Günther, 1858
- Colostethidae Cope, 1867
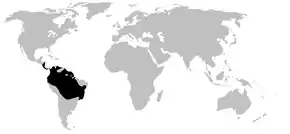
Répartition géographique
Description
Anatomie
La plupart des espèces de Dendrobatidae sont de petite taille à l'âge adulte, allant de moins de 15 mm de long pour moins d'1 gramme à 6 cm de long pour environ 28 g[4]. La plupart des Dendrobatidae affichent des motifs aposématiques aux couleurs vives avertissant les prédateurs potentiels de leur dangerosité. Leur coloration vive est associée à leur toxicité et les niveaux d'alcaloïdes contenus dans leur peau. Les grenouilles du genre Dendrobates ont des niveaux élevés d'alcaloïdes, alors que les espèces du genre Colostethus sont cryptiques et ne sont pas toxiques[5].
Les Dendrobatidae sont un exemple d'organisme aposématique. Leur coloration vive annonce leur inappétence pour les prédateurs potentiels. On pense que l'aposématisme est apparu au moins quatre fois au sein de la famille des Dendrobatidae selon les arbres phylogénétiques, et ces grenouilles ont depuis subi des divergences spectaculaires - à la fois interspécifiques et intraspécifiques - dans leur coloration. Ceci est surprenant étant donné que ce type de mécanisme de défense dépend de la fréquence à laquelle ces animaux sont présents[6] - [7].
Couleurs
Certaines espèces de Dendrobatidae présentent un certain nombre de colorations différentes au sein d'une même espèce, et ce polymorphisme semblent être apparu récemment, il y a 6 000 ans[8]. Par conséquent, les espèces telles que Dendrobates tinctorius, Oophaga pumilio et O. granulifera peuvent avoir un motif de couleur qui peut varier suivant les croisements (les couleurs sont sous contrôle polygénique, tandis que les motifs sont probablement contrôlés par un seul locus)[9]. Les différences de coloration ont historiquement conduit à identifier les individus d'une même espèce comme appartenant à deux espèces différentes, et cela explique le grand nombre de controverses parmi les taxonomistes pour la classification de ces espèces[10].
Les variations dans les régimes de prédation peuvent avoir influencé l'évolution du polymorphisme chez O. granulifera[11], tandis que la sélection sexuelle semble avoir contribué à la différenciation entre les populations de Oophaga pumilio à Bocas del Toro[12] - [13] - [14].
Biologie et écologie
Reproduction
Les espèces de la famille des Dendrobatidae présentent un spectre de comportements reproductifs marqués par un rituel nuptial et des soins parentaux contrastant avec la majorité des anoures[15].
Les Dendrobatidae se reproduisent plusieurs fois dans l'année, la période de reproduction commence typiquement au début de la saison des pluies.
Les mâles sont particulièrement territoriaux lors de cette période et se disputent en luttant les emplacements à partir desquels ils émettent leur cri ainsi que les sites de ponte. Des luttes entre femelles sont également présentes chez certaines espèces et conditionnent l'accès aux mâles et aux sites de ponte[16]. Les femelles inspectent les mâles et choisissent alors leur partenaires en fonction de son apparence et de l'emplacement choisi pour l'appeler[8]. Chez la majorité des espèces, le mâle amène ensuite la femelle au site de ponte. Certains Dendrobatidae forment alors un amplexus céphalique caractéristique, le mâle s'accrochant au dos de la femelle en la maintenant par la tête, tandis que d'autres ne forment pas d'amplexus[17]. Les œufs sont pondus hors de l'eau dans des endroits humides, comme sur les feuilles, dans les plantes, sur des racines exposées, les œufs sont fertilisés par fécondation extérieure.
Une fois éclos, les têtards sont déplacés individuellement ou en groupe sur le dos d'un parent pour être acheminés jusqu'à un milieu aquatique où ils se développeront. En fonction des espèces, la dépôt des têtards a lieu au niveau du sol dans un plan d'eau stagnante ou un ruisseau, ou bien dans les phytotelmes de plantes terrestres ou épiphytes tels que les réserves d'eau des broméliacées [18]. Chez la majorité des espèces les soins parentaux persistent sous diverses formes telles que le déplacement des têtards lorsque la réserve d'eau s'assèche ou le nourrissage des têtards par la femelle avec des œufs infertiles, jusqu'à la métamorphose des têtards [19].
La métamorphose survient entre 50 et 80 jours après la ponte et les jeunes atteignent la maturité sexuelle au cours de leur première année.
| Espèce | Agressions | Structure parentale | Soins parentaux | Nombre de têtards | Site de dépôt | ||
|---|---|---|---|---|---|---|---|
| Sexe(s) | Type(s) | ||||||
| Colostethus panamensis |  |
Territoriaux envers tous sexes. | Polygame | Femelle | Dépôt | 20-35 | Ruisseau |
| Oophaga pumilio |  |
Mâles-Mâles | Polygame | Femelle | Dépôt,
Déplacement, Nourrissage
|
1-4 | Phytotelmes de broméliacées. |
| Ranitomeya imitator | Mâles-Mâles | Monogame | Femelle
Mâles |
Dépôt(♂), Déplacement(♂), Nourrissage (♀) | 1 | Phytotelmes de plantes terrestres comme Heliconia, Dieffenbachia, ou Xanthosoma[21] | |
Alimentation
Les Dendrobatidae adultes sont carnivores et se nourrissent de petits arthropodes qu'ils chassent activement durant la journée. Des spécialisations alimentaires sont apparues à plusieurs reprises dans la famille des Dendrobatidae et sont fortement corrélées à la toxicité et à l'aposématisme des espèces spécialistes. Celles-ci sont en effet capable de séquestrer les alcaloïdes présents dans leur alimentation, principalement composée de fourmis et d'acariens oribates et développent ainsi une toxicité les protégeant des prédateurs [22] - [23].
Les têtards sont principalement carnivores à forte tendance cannibales. Ils se nourrissent d'arthropodes aquatiques et de têtards de la même espèce ou d'autres amphibiens[24]. Différentes recherches montrent que cette prédation permet également aux têtards d'éliminer les autres têtards et larves d'insectes prédatrices lorsque ces derniers sont encore trop petits pour les attaquer et les manger. Chez certaines espèces, l'alimentation est complétée par des œufs infertiles pondus par la femelle, voir majoritairement comprise de ces œufs chez les Dendrobatidae les plus spécialisés[19].
Répartition et habitat
Les Dendrobatidae sont endémiques des environnements tropicaux humides d'Amérique Centrale et du Sud[2]. Ces grenouilles sont généralement rencontrées dans les forêts tropicales, notamment en Bolivie, au Costa Rica, au Brésil, en Colombie, en Équateur, au Venezuela, au Suriname, en Guyane française, au Pérou, au Panama, au Guyana, au Nicaragua, et à Hawaii (où ils sont introduits)[2] - [25].
Leurs habitats naturels comprennent les forêts subtropicales et tropicales humides de plaine, les terres arbustives subtropicales ou tropicales de haute altitude, les montagnes, rivières, marais d'eau douce, lacs et marécages sous un climat subtropicale ou tropicale. Certaines espèces peuvent être rencontrées dans des prairies saisonnièrement humides ou des plaines inondées, les terres arables, les pâturages, les jardins, les plantations, la savane humide et les anciennes forêts fortement dégradées. Les Dendrobatidae ont tendance à vivre sur ou près du sol, mais aussi dans les arbres jusqu'à 10 m au-dessus du sol[26].
Taxonomie
Les Dendrobatidae forment avec leur famille sœur Aromobatidae la superfamille des Dendrobatoidae. La famille des Dendrobatidae contient selon Amphibian Species of the World (04 avril 2020)[27] 17 genres, avec 199 espèces[28] - [29], elle est cependant l'objet de diverses études phylogénétiques et la taxonomie des espèces lui appartenant évolue fréquemment[30] .
| Genres selon Amphibian Species of the World (04 avril 2020)[27] | Cladogramme dérivé, selon Grant et al. (2017)[31] |
|---|---|
|
Toxicité et médecine

Beaucoup de grenouilles de la famille des Dendrobatidae sécrètent des toxines alcaloïdes lipophiles à travers leur peau. Les alcaloïdes contenus dans les glandes de la peau des grenouilles servent comme moyen de défense chimique contre les prédateurs, et ils sont donc en mesure d'être actifs face aux prédateurs potentiels pendant la journée. Environ 28 classes structurelles d'alcaloïdes sont connues chez ces grenouilles[2] - [32]. Parmi ces espèces la plus toxique est Phyllobates terribilis. Certains scientifiques pensent que les Dendrobatidae ne synthétisent pas leurs poisons, mais séquestrent les produits chimiques de certains arthropodes qu'ils consomment, comme les fourmis, mille-pattes et acariens[33] - [34]. Pour cette raison, les animaux élevés en captivité ne présentent pas les niveaux significatifs de toxines de ceux vivant dans le milieu naturel, car leurs régimes sont différents. En fait, de nouvelles études suggèrent que les femelles de certaines espèces pondent des œufs non fécondés, qui présentent des traces d'alcaloïdes, pour nourrir les têtards[35]. Ce comportement montre que les poisons sont introduits à partir d'un très jeune âge. Néanmoins, les grenouilles élevées en captivité conservent la capacité d'accumuler des alcaloïdes quand on leur fourni une alimentation en contenant[36]. Malgré les toxines utilisées par certaines Dendrobatidae, des prédateurs ont développé la capacité de leur résister. Par exemple le serpent Liophis epinephelus a développé une immunité au poison[37].
Les molécules chimiques extraites de la peau d'Epipedobates tricolor ont une valeur médicinale. Les scientifiques utilisent ce poison comme analgésique[38]. Parmi les molécules extraites on trouve en effet un analgésique 200 fois plus puissant que la morphine, l'épibatidine. Cependant, la dose thérapeutique est très proche de la dose mortelle[39]. Un dérivé, ABT-594, développé par les Laboratoires Abbott, appelée Tebanicline est entré en deuxième phase de test chez l'Homme[40], mais son développement a été stoppé en raison des effets secondaires gastro-intestinaux trop importants[41]. Les sécrétions des Dendrobatidae peuvent également être intéressantes comme relaxants musculaires, stimulants cardiaques et anorexigènes[42]. La plus toxique de ces grenouilles, Phyllobates terribilis, dispose d'assez de toxines en moyenne pour tuer dix à vingt hommes ou environ dix mille souris[43]. La plupart des autres Dendrobatidae, même si elles sont colorées et suffisamment toxiques pour décourager les prédateurs, comportent beaucoup moins de dangers pour les humains ou d'autres animaux de grande taille.
Évolution de la coloration de la peau et de la toxicité
Une première étude menée par Summers et Clough (2000) sur l'évolution de la coloration de la peau et de la toxicité dans la famille des Dendrobatidae a indiqué que l'évolution de la toxicité de la peau était significativement corrélée avec l'évolution de la coloration vive[44]. Par ailleurs, une étude réalisée par Santos et Cannatella (2011) a confirmé cela en expliquant qu'il y avait une corrélation remarquable entre la coloration et la spécialisation de l'alimentation, la masse corporelle, la capacité aérobie, et la défense chimique chez les Dendrobatidae[7]. Santos et Cannatella (2011) proposent deux scénarios évolutifs qui expliquent ces corrélations. Une possibilité est que l'aposématisme et la capacité aérobie ont précédé une plus grande collecte de ressources, les grenouilles consommant plus de fourmis et d'acariens et spécialisant leur alimentation. Ceci est contraire à la théorie aposématique classique, qui considère que la toxicité de l'alimentation est apparue avant la coloration vive. La deuxième hypothèse suggère que la spécialisation de l'alimentation a précédé la capacité aérobie supérieure et que l'aposématisme a évolué afin de permettre aux Dendrobatidae de partir à la recherche de nourriture sans risquer la prédation.
D'autres données indiquent que la coloration et la toxicité peuvent être inversement proportionnelles. Une étude récemment publiée par Wang et Shaffer (2011) soutient cette idée. Cette étude a révélé que chez les grenouilles Dendrobatidae polymorphes, les animaux aux coloris les moins visibles sont plus toxiques que les espèces les plus brillantes et les plus visibles[45]. Les dépenses énergétiques pour produire les toxines et les pigments de couleurs vives conduisent à des compromis possibles entre la toxicité et coloration vive[46]. Les proies avec de fortes défenses secondaires (comme les grenouilles toxiques dans l'étude Wang et Shaffer 2011) ont moins à gagner avec une peau colorée coûteuse en énergie. Par conséquent, les populations de grenouilles très toxiques manifestent des signaux extérieurs de dangerosité moins évidents[47]. Cette théorie conteste le principe que l'augmentation de la coloration de la peau évolue toujours avec une toxicité accrue.
La mobilité des proies pourrait également expliquer le développement initial de la signalisation aposématique. Si les proies ont des caractéristiques qui les rendent plus exposées aux prédateurs, telles que la taille ou l'habitat, ils ont de bonnes raisons de développer de l'aposématisme. Un exemple de cela est le passage du comportement nocturne au comportement diurne chez certaines Dendrobatidae[6]. Après le passage à un comportement diurne, les grenouilles avaient de plus grandes possibilités en termes de biologie, et en conséquence la spécialisation alimentaire est apparue[48]. Ainsi, l'aposématisme est non seulement un système de signalisation, mais un moyen pour les organismes d'avoir un meilleur accès aux ressources et à accroître leur succès comme reproducteurs.
Marples et al., cependant, a montré que les habitudes alimentaires (néophobie à long terme) des prédateurs pourraient faciliter l'évolution de la coloration d'avertissement si les prédateurs évitent les nouvelles formes pour une période de temps suffisamment longue[49].
De nombreuses études ont suggéré que la sélection sexuelle joue un rôle dans la diversification de la couleur et des motifs de la peau des Dendrobatidae[50] - [51] - [52] - [53]. Comme les préférences des femelles jouent alors un rôle important, la coloration des mâles devrait évoluer plus rapidement. La sélection sexuelle est influencée par beaucoup de choses. L'investissement parental peut expliquer à un certain point l'évolution de la coloration par rapport au choix de la femelle. Par exemple, chez Oophaga pumilio les deux sexes ne sont pas impliqués de la même manière dans le soin aux jeunes. La femelle pond les œufs et s'occupe de la progéniture pendant plusieurs semaines tandis que les mâles déposent le sperme (ce qui demande moins d'énergie) et ne s'occupent des jeunes que pendant quelques jours. Cette différence entre sexes indique qu'il y aura une forte incidence du choix des femelles. La sélection sexuelle fait fortement augmenter la variation phénotypique chez les espèces. Tazzyman et Iwasa (2010) ont constaté qu'il y avait des populations de O. pumilio qui utilisaient la sélection sexuelle, et d'autres non. Dans les premières populations, le polymorphisme phénotypique était évident[54].
L'absence de fort dimorphisme sexuel dans certaines populations de Dendrobatidae suggère que la sélection sexuelle n'est pas une hypothèse valable de l'aposématisme de ces grenouilles[55]. Ce pourrait être parce que la mesure de protection principale des populations de grenouilles aposématiques est leur capacité à avertir les prédateurs de leur toxicité. Rûdh et al. (2011) ont prédit que la préférence des femelles pour les mâles plus vifs augmenterait la vivacité de la coloration chez les deux sexes et accroîtrait ainsi leurs aptitudes à avertir les agresseurs. S'il y avait un dimorphisme sexuel entre les mâles et les femelles au sein d'une population aposématique il y aurait une différence au niveau de la prédation des mâles et des femelles qui provoquerait un déséquilibre dans le sex-ratio, ce qui aurait une incidence sur les stratégies d'accouplement et, finalement modifierait le comportement d'accouplement de l'espèce.
Relations avec l'Homme
Statut de sauvegarde

De nombreuses espèces de Dendrobatidae ont connu récemment une disparition de leur habitat (en raison de l'exploitation forestière et l'agriculture), et sont touchées par des maladies telle que la chytridiomycose. Certaines espèces sont donc répertoriées comme menacées ou en danger[56]. Les zoos ont essayé d'enrayer cette maladie en traitant les grenouilles en captivité avec un agent antifongique qui est normalement utilisé pour soigner les pieds des athlètes[57].
Élevage en captivité
Toutes les espèces de grenouilles venimeuses sont d'origine néotropicale. Les spécimens capturés dans la nature peuvent maintenir la toxicité pendant un certain temps, de sorte qu'il faut prendre de grandes précautions lors de la manipulation de ces animaux[58]. En captivité, la plupart des espèces vivent correctement lorsque l'humidité est maintenue constante vers 80 à 100% et lorsque la température est d'environ 22 °C à 27 °C pendant la journée et pas inférieure à 16 °C à 18 °C pendant la nuit. Certaines espèces tolèrent des températures plus basses mieux que d'autres.
Publication originale
- Cope, 1865 : Sketch of the Primary Groups of Batrachia Salientia. The Natural History Review, New Serie, vol. 5, p. 97–120 (texte intégral).
Liens externes
- (en) Référence Animal Diversity Web : Dendrobatidae (consulté le )
- (en) Référence AmphibiaWeb : famille Dendrobatidae (+ liste espèces) (consulté le )
- (en) Référence Amphibian Species of the World : Dendrobatidae Cope, 1865 (consulté le )
- (en) Référence Catalogue of Life : Dendrobatidae Cope, 1865 (consulté le )
- (fr+en) Référence ITIS : Dendrobatidae Cope, 1865 (consulté le )
- (en) Référence NCBI : Dendrobatidae (taxons inclus) (consulté le )
- (en) Référence Tree of Life Web Project : Dendrobatidae (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023
- (en) Référence Wild Herps : photographies de Dendrobatidae (consulté le )
Notes et références
- Amphibian Species of the World, consulté lors d'une mise à jour du lien externe
- « AmphibiaWeb – Dendrobatidae », AmphibiaWeb (consulté le )
- Heying, H., « Dendrobatidae », Animal Diversity Web, (consulté le )
- (en) Ted R. Kahn, Enrique La Marca, Stefan Lötters et al., Aposematic poison frogs (Dendrobatidae) of the Andean countries : Bolivia, Colombia, Ecuador, Perú and Venezuela, Arlington, Virginia, Conservation International., , 588 p. (ISBN 978-1-934151-27-3)
- Caldwell, J.P., « The evolution of myrmecophagy and its correlates in poison frogs (Family Dendrobatidae). », Journal of Zoology, vol. 240, no 1, , p. 75–101 (DOI 10.1111/j.1469-7998.1996.tb05487.x)
- J.C. Santos, L.A. Coloma et D.C. Cannatella, « Multiple, recurring origins of aposematism and diet specialization in poison frogs », Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no 22, , p. 12792–12797 (PMID 14555763, PMCID 240697, DOI 10.1073/pnas.2133521100)
- J. C. Santos et D. C. Cannatella, « Phenotypic integration emerges from aposematism and scale in poison frogs », Prodeedings of the National Academy of Sciences of the United States of America, vol. 108, no 15, , p. 6175–6180 (PMID 21444790, PMCID 3076872, DOI 10.1073/pnas.1010952108)
- (en) Summers, K; Symula, R; Clough, M; Cronin, T, « Visual mate choice in poison frogs », Proceedings of the Royal Society of London B: Biological Sciences, vol. 266, no 1434, , p. 2141–5 (PMID 10649631, PMCID 1690338, DOI 10.1098/rspb.1999.0900)
- Summers K., Cronin T. W., Kennedy T., « Cross-breeding of distinct color morphs of the strawberry poison frog (Dendrobates pumilio) from the Bocas del Toro Archipelago, Panama », Journal of Herpetology, vol. 38, no 1, , p. 1–8 (DOI 10.1670/51-03A)
- Kok, PJR; MacCulloch, RD; Gaucher, P; Poelman, EH; Bourne, GR; Lathrop, A; Lenglet, GL, « A new species of Colostethus (Anura, Dendrobatidae) from French Guiana with a redescription of Colostethus beebei (Noble, 1923) from its type locality », Phyllomedusa, vol. 5, no 1, , p. 43–66 (DOI 10.11606/issn.2316-9079.v5i1p43-66, lire en ligne)
- I. J. Wang, « Inversely related aposematic traits: reduced conspicuousness evolves with increased toxicity in a polymorphic poison-dart frog », Evolution, vol. 65, no 6, , p. 1637–1649 (PMID 21644954, DOI 10.1111/j.1558-5646.2011.01257.x)
- M. E. Maan et M. E. Cummings, « Female preferences for aposematic signal components in a polymorphic poison frog », Evolution, vol. 62, no 9, , p. 2234–2345 (PMID 18616568, DOI 10.1111/j.1558-5646.2008.00454.x)
- R. G. Reynolds et B. M. Fitzpatrick, « Assortative mating in poison-dart frogs based on an ecologically important trait », Evolution, vol. 61, no 9, , p. 2253–2259 (PMID 17767594, DOI 10.1111/j.1558-5646.2007.00174.x)
- I.J. Tazzyman et Y. Iwassa, « Sexual selection can increase the effect of random genetic drift – a quantitative genetic model of polymorphism in Oophaga pumilio, the strawberry poison-dart frog », Evolution, vol. 64, no 6, , p. 1719–1728 (PMID 20015236, DOI 10.1111/j.1558-5646.2009.00923.x)
- (en) Martha L. Crump, « Parental care among the Amphibia », Advances in the Study of Behavior, vol. 25, , p. 109–144 (DOI 10.1016/S0065-3454(08)60331-9)
- (en) Summers, « Mating strategies in two species of dart-poison frogs: a comparative study », Animal Behaviour, , p. 907-919 (DOI 10.1016/S0003-3472(06)80004-7)
- (en) Heather Heying, « Dendrobatidae », sur animaldiversity.org (consulté le )
- (en) Kentwood D. Wells, The Ecology and Behavior of Amphibians, University of Chicago Press, , 1400 p. (ISBN 9780226893334 et 0226893332, DOI 10.7208/chicago/9780226893334.001.0001), Chapitre 11 : p524-526
- (en) Sexual Selection : Perspectives and Models from the Neotropics, , 466 p. (ISBN 978-0-12-416028-6, DOI 10.1016/C2011-0-05695-8), chap. 11, p. 295-297
- (en) Sexual Selection : Perspectives and Models from the Neotropics, , 466 p. (ISBN 978-0-12-416028-6, DOI 10.1016/C2011-0-05695-8), chap. 11
- (en) « Ranitomeya imitator », sur www.dendrobates.org (consulté le )
- (en) Catherine Darst, Pablo A Menéndez-Guerrero, Luis A. Coloma et David Cannatella, « Evolution of Dietary Specialization and Chemical Defense in Poison Frogs (Dendrobatidae): A Comparative Analysis », The American Naturalist 165(1), , p. 56-69 (DOI 10.1086/426599, lire en ligne)
- (en) A. Saporito, « Arthropod Alkaloids in Poison Frogs: A Review of the “Dietary Hypothesis. », Heterocycles, 79(1), , p. 277 (DOI 10.3987/REV-08-SR(D)11)
- (en) Janalee P. Caldwell et Maria Carmozina de Araujo, « Cannibalistic Interactions Resulting from Indiscriminate Predatory Behavior in Tadpoles of Poison Frogs (Anura: Dendrobatidae) », Biotropica, 30(1), , p. 92-103 (DOI 10.1111/j.1744-7429.1998.tb00372.x)
- « Poison Dart Frogs in Hawaii », Explore Biodiversity (consulté le )
- Hurme, Kristiina; Gonzalez, Kittzie; Halvorsen, Mark; Foster, Bruce and Moore, Don, « Environmental Enrichment for Dendrobatid Frogs », Journal of Applied Animal Welfare Science, Lawrence Erlbaum Associates, Inc., vol. 6, no 4, , p. 285–299 (PMID 14965783, DOI 10.1207/s15327604jaws0604_3)
- Amphibian Species of the World, consulté le 04 avril 2020
- « Amphibian Species of the World », The American Museum of Natural History (consulté le )
- Pough, Frederick Harvey; Andrews, Robin M.; Cadle, John E. and Crump, Martha L., Herpetology, Upper Saddle River, NJ, Pearson/Prentice Hall, (ISBN 0-13-100849-8), p. 92
- T. Grant, D.R. Frost, J.P. Caldwell, R. Gagliardo, C.F.B. Haddad, P.J.R. Kok, D.B. Means, B.P. Noonan, W.E. Schargel et W.C. Wheeler, « Phylogenetic systematics of dart-poison frogs and their relatives (Amphibia: Athesphatanura: Dendrobatidae) », Bulletin of the American Museum of Natural History, American Museum of Natural History, vol. 299, no 299, , p. 1–262 (DOI 10.1206/0003-0090(2006)299[1:PSODFA]2.0.CO;2, lire en ligne)
- (en) Grant et al., « Phylogenetic Systematics of Dart-Poison Frogs and Their Relatives Revisited (Anura: Dendrobatoidea) », South American Journal of Herpetology 12(s1), (ISSN 1982-355X, lire en ligne)
- David Cannatella, « Dendrobatidae. Poison-arrow frogs, Dart-poison frogs, Poison-dart frogs », The Tree of Life Project, (consulté le )
- Darst, C., Menendez-Guerreor, P., Coloma, L. and Cannatella, D., « Evolution of Dietary Specialization and Chemical Defense in Poison Frogs (Dendrobatidae): A Comparative Analysis », The American Natural, vol. 165, no 1, , p. 56–69 (PMID 15729640, DOI 10.1086/426599)
- Daly, J.W., Gusovsky, F., Myers, C.W., Yotsuyamashita, M., and Yasumoto, T., « First occurrence of tetrodotoxin in a dendrobatid frog (Colostethus inguinalis), with further reports for the bufonid genus Atelopus », Toxicon, vol. 32, no 3, , p. 279–285 (PMID 8016850, DOI 10.1016/0041-0101(94)90081-7)
- Jennifer Stynoski, Yaritbel Torres-Mendoza, Mahmood Sasa-Marin et Ralph Saporito, « Evidence of maternal provisioning of alkaloid-based chemical defenses in the strawberry poison frog Oophaga Pumilio », Ecology, vol. 95, no 3, , p. 587–93 (PMID 24804437, DOI 10.1890/13-0927.1)
- R. Saporito, M. Donnelly, R. Norton, H. Garraffo, T. Spande et J. Daly, « Oribatid mites as a major dietary source for alkaloids in poison frogs », Proceedings of the National Academy of Sciences of the United States of America, vol. 104, no 21, , p. 8885–8890 (PMID 17502597, PMCID 1885597, DOI 10.1073/pnas.0702851104)
- Myers, C.W.; Daly, J.W. and Malkin, B., « A dangerously toxic new frog (Phyllobates) used by the Emberá Indians of western Colombia, with discussion of blowgun fabrication and dart poisoning », Bulletin of the American Museum of Natural History, vol. 161, no 2, , p. 307–365 + color pls. 1–2
- John Emsley, « Science: Potent painkiller from poisonous frog », New Scientist, (version du 7 avril 2010 sur Internet Archive)
- R.J. Prince et S.M. Sine, « Epibatidine activates muscle acetylcholine receptors with unique site selectivity », Biophysical Journal, Biophysical Journal, vol. 75, no 4, , p. 1817–27 (PMID 9746523, PMCID 1299853, DOI 10.1016/S0006-3495(98)77623-4)
- M. Decker, M. Meyer et J. Sullivan, « The therapeutic potential of nicotinic acetylcholine receptor agonists for pain control », Expert Opinion on Investigational Drugs, vol. 10, no 10, , p. 1819–1830 (PMID 11772288, DOI 10.1517/13543784.10.10.1819)
- Meyer MD. Neuronal Nicotinic Acetylcholine Receptors as a Target for the Treatment of Neuropathic Pain. Drug Development Research. 2006; 67: 355-359.
- « San Diego Zoo's Animal Bytes: Poison Frog », Zoological Society of San Diego (consulté le )
- « Most poisonous creature on earth could be a mystery insect », Wonderquest.com (consulté le )
- K. Summers et M. Clough, « The evolution of coloration and toxicity in the poison frog family (Dendrobatidae) », Proceedings of the National Academy of Sciences of the United States of America, vol. 98, no 11, , p. 6227–6232 (PMID 11353830, PMCID 33450, DOI 10.1073/pnas.101134898)
- I. Wang et H.B. Shaffer, « Rapid Color Evolution in an Aposematic Species: A Phylogenetic Analysis of Color Variation in the Strikingly Polymorphic Strawberry Poison-Dart Frog », Evolution, vol. 62, no 11, , p. 2742–2759 (PMID 18764916, DOI 10.1111/j.1558-5646.2008.00507.x)
- I Speed, M.A. Brockhurst et G.D. Ruxton, « The dual benefits of aposematism: Predator avoidance and enhanced resource collection », Evolution, vol. 64, no 6, , p. 1622–1633 (PMID 20050915, DOI 10.1111/j.1558-5646.2009.00931.x)
- I. Speed, G.D. Ruxton, J.D. Blount et P.A. Stephens, « Diversification of honest signals in a predator-prey system », Ecology Letters, vol. 13, no 6, , p. 744–753 (PMID 21644954, DOI 10.1111/j.1558-5646.2011.01257.x)
- K. Summers, « Convergent evolution of bright coloration and toxicity in frogs », Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no 22, , p. 12533–12534 (PMID 14569014, PMCID 240648, DOI 10.1073/pnas.2335928100)
- N.M. Marples, DJ Kelly et RJ Thomas, « Perspective: The evolution of warning coloration is not paradoxical », Evolution, vol. 59, no 5, , p. 933–940 (PMID 16136793, DOI 10.1111/j.0014-3820.2005.tb01032.x)
- M.E. Mann et M.E. Cummings, « Sexual dimorphism and directional sexual selection on aposematic signals in a poison frog », Proceedings of the National Academy of Sciences of the United States of America, vol. 106, no 45, , p. 19072–19077 (PMID 19858491, PMCID 2776464, DOI 10.1073/pnas.0903327106)
- K Summers, L. Bermingham, S. Weigt, S. McCafferty et L. Dahlstrom, « Phenotypic and genetic divergence in three species of dart-poison frogs with contrasting parental behavior », The Journal of Heredity, vol. 88, no 1, , p. 8–13 (PMID 9048443, DOI 10.1093/oxfordjournals.jhered.a023065)
- A. Rudh, B. Rogell et J. Hoglund, « Non-gradual variation in color morphs of the strawberry poison frog Dendrobates pumilio: genetic and geographical isolation suggest a role for selection in maintaining polymorphism », Molecular Ecology, vol. 16, no 20, , p. 4282–4294 (PMID 17868297, DOI 10.1111/j.1365-294X.2007.03479.x)
- M. E. Maan et M. E. Cummings, « Sexual dimorphism and directional selection on aposematic signals in a poison frog », Proceedings of the National Academy of Sciences of the United States of America, vol. 106, no 45, , p. 19072–19077 (PMID 19858491, PMCID 2776464, DOI 10.1073/pnas.0903327106)
- S.J. Tazzyman et Y. Iwasa, « Sexual selection can increase the effect of random genetic drift-a quantitative genetic model of polymorphism in oophaga pumilio, the strawberry poison-dart frog », Evolution, vol. 64, no 6, , p. 1719–1728 (PMID 20015236, DOI 10.1111/j.1558-5646.2009.00923.x)
- Andreas Rudh, B. Rogell, O. Håstad et =A. Qvarnström, « Rapid population divergence linked with co-variation between coloration and sexual display in strawberry poison frogs », Evolution, vol. 65, no 65, , p. 1271–1282 (DOI 10.1111/j.1558-5646.2010.01210.x)
- P Daszak, L Berger, AA Cunningham, AD Hyatt, DE Green et R. Speare, « Emerging infectious diseases and amphibian population declines », Emerg. Infect. Dis., vol. 5, no 5, , p. 735–48 (DOI 10.3201/eid0506.990601)
- « Poison Dart Frog Fact Sheet – National Zoo| FONZ », National Zoological Park (consulté le )
- Lötters Stefan, Jungfer, Henkel et Schmidt, Poison Frogs : Biology, Species, & Captive Husbandry, Serpent's Tale, , 668 p. (ISBN 978-3-930612-62-8 et 3-930612-62-3)