Mammouth laineux
Mammuthus primigenius, Elephas primigenius

Südostbayerisches Naturkunde- und Mammut-Museum de Siegsdorf, Allemagne.
Le mammouth laineux (Mammuthus primigenius ou Elephas primigenius) est une espèce éteinte de la famille des éléphantidés qui a vécu durant le Pléistocène et, pour ses derniers représentants, au cours de l'Holocène il y a seulement 4 000 ans. Le genre Mammuthus était apparu avec Mammuthus subplanifrons au début du Pliocène.
Le mammouth laineux a divergé du mammouth des steppes il y a environ 800 000 ans dans l'est de l'Asie. Son plus proche parent est l'éléphant d'Asie. L'apparence et le comportement de cette espèce sont parmi les mieux étudiés pour un animal préhistorique du fait de la découverte de carcasses prises dans les glaces en Sibérie et en Alaska, ainsi que de squelettes, de dents, de contenu stomacal, de défenses et de représentations sur les parois des grottes. Des traces fossilisées de mammouths ont été trouvées en Asie bien avant que les Européens n'en prennent connaissance au XVIIe siècle. Les origines de ces fossiles ont fait l'objet de nombreux débats, et on les a souvent associés à des restes de créatures légendaires. Le mammouth a été identifié comme une espèce éteinte d'éléphant par Georges Cuvier en 1796.
Le mammouth laineux avait une taille relativement proche de celle de l'éléphant d'Afrique actuel. Les mâles atteignaient une hauteur aux épaules comprise entre 2,7 et 3,4 m et pesaient jusqu'à 6 tonnes. La taille des femelles variaient entre 2,6 et 2,9 m pour un poids qui pouvait atteindre 4 tonnes. Un nouveau-né pesait environ 90 kg.
Le mammouth laineux était bien adapté à l'environnement froid de la dernière période glaciaire. Il était couvert de fourrure, avec de longs poils de jarre et une sous-couche de poils de bourre. La couleur de sa fourrure était plus ou moins sombre. Les oreilles et la queue étaient courtes, limitant les risques de gelure et de déperdition de chaleur. Il avait de longues défenses incurvées, et quatre molaires qui étaient remplacées six fois au cours de la vie d'un individu. Son comportement était similaire à celui des éléphants modernes, et il utilisait ses défenses et sa trompe pour manipuler des objets, se battre et s'alimenter. L'alimentation du mammouth laineux était principalement composée d'herbe et de cypéracées. Les animaux pouvaient probablement atteindre l'âge de 60 ans. Ils vivaient dans les steppes de l'Eurasie et de l'Amérique du Nord.
Le mammouth laineux était contemporain des premiers Hommes, qui utilisaient ses os et ses défenses pour faire des outils, des objets décoratifs ou des habitations, et cette espèce était également chassée pour la nourriture. Elle a disparu de son aire de répartition continentale à la fin du Pléistocène il y 10 000 ans, certainement du fait du changement climatique et de la réduction de son habitat qui en a découlé, voire de la chasse par l'Homme. Des populations isolées ont survécu sur l'île Saint-Paul jusqu'à il y a 6 400 ans et sur l'île Wrangel jusqu'à il y a 4 000 ans. Après son extinction, les Hommes ont continué à utiliser son ivoire, une tradition qui se poursuit encore de nos jours. La possibilité de récréer cette espèce par clonage a été évoquée à plusieurs reprises, mais cela semble improbable du fait du stade de dégradation important du matériel génétique retrouvé à ce jour.
Description
Apparence générale

L'apparence du mammouth laineux est probablement la mieux connue de tous les animaux préhistoriques car plusieurs individus aux tissus préservés ont été retrouvés pris dans la glace. De plus cet animal a été très fréquemment représenté par les premiers hommes. Les mâles adultes pouvaient atteindre une taille aux épaules de 2,7 à 3,4 m et pesaient jusqu'à 6 tonnes. Il est presque aussi grand que les éléphants d'Afrique modernes, qui atteignent communément 3 à 3,4 m, mais plus petit que les espèces de mammouths plus primitives tels que le mammouth méridional (Mammuthus meridionalis) et le mammouth des steppes (Mammuthus trogontherii), ou que son contemporain le mammouth de Colomb (Mammuthus columbi). La raison pour cette taille inférieure n'est pas connue. Les femelles mesuraient en moyenne 2,6 à 2,9 m de hauteur aux épaules et étaient un peu plus frêles que les mâles, pesant au plus 4 tonnes. Un nouveau-né pesait environ 90 kg. Ces tailles sont estimées à partir de celles des éléphants modernes de taille similaire[1]. Peu de spécimens retrouvés congelés avaient leurs organes génitaux intacts, et le sexe est souvent déterminé à partir du squelette. La meilleure indication pour le déterminer est ainsi la ceinture pelvienne, car l'ouverture qui permet le passage du nouveau-né est toujours plus large chez les femelles que chez les mâles[2]. Bien que les mammouths de l'île de Wrangel aient été plus petits que ceux du continent, leur taille était variable et ils n'étaient pas suffisamment petits pour être qualifiés de « nains »[3]. Il a été dit que les dernières populations de Mammouths laineux avaient décliné en taille et présentaient un dimorphisme sexuel plus marqué, mais cela a été contredit par une étude de 2012[4].
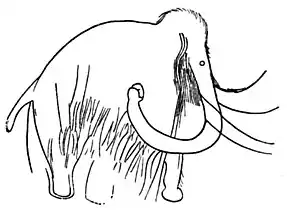
Les Mammouths laineux présentent diverses adaptations au froid, dont la plus notable est la robe de fourrure qui couvrait l'intégralité de son corps. Par ailleurs leurs oreilles étaient nettement plus petites que celles des éléphants modernes, mesurant environ 38 cm de long et entre 18 et 28 cm de large. L'oreille d'un jeune de 6 à 12 mois retrouvé congelé, « Dima », mesurait moins de 13 cm de long. Cette petitesse des oreilles permettait de limiter l'exposition au froid et donc les déperditions de chaleur ou les gelures. Pour les mêmes raisons, la queue était courte, mesurant seulement 36 cm chez le « Berezovka mammoth ». La queue comprenait 21 vertèbres, tandis que celle des éléphants modernes en présentent de 28 à 33. Tous ces points illustrent typiquement la règle d'Allen.
Leur peau n'était pas plus épaisse que celle des éléphants, entre 1,25 et 2,5 cm. Ils disposaient d'une couche de graisse pouvant atteindre 10 cm d'épaisseur sous la queue, qui les aidait à conserver leur chaleur corporelle. Les Mammouths laineux avaient de larges lambeaux de peau sous leur queue qui couvraient l'anus, une caractéristique que l'on retrouve chez les éléphants modernes[5].


Parmi les autres caractéristiques dépeintes sur les parois des grottes, on note une grande et haute tête, et un dos incliné avec des épaules très hautes. Cette forme résulte des vertèbres du dos dont la longueur diminue du devant vers l'arrière. Ces caractéristiques ne sont pas visibles chez les jeunes, qui ont un dos convexe comme les éléphants d'Asie. Les peintures rupestres ont également permis d'identifier une caractéristique de la trompe des mammouths qui a pu être confirmée par la découverte d'un spécimen pris dans la glace en 1924 et surnommé « Middle Kolyma mammoth », préservé avec une trompe intacte. À la différence de l'extrémité de la trompe des éléphants modernes, le « doigt » supérieur de l'extrémité de la trompe des mammouths a un lobe pointu et allongé mesurant 10 cm, tandis que le « pouce » inférieur mesure 5 cm et est plus large. La trompe de « Dima » mesurait 76 cm de long, tandis que celle du « mammouth de Lyakhov » (dont le squelette est exposé au Muséum national d'histoire naturelle à Paris) faisait 2 m de long[5]. La trompe très bien préservée d'un spécimen juvénile surnommé « Yuka » a été décrite en 2015 ; elle présentait une excroissance de chair un tiers au-dessus de son extrémité. Contrairement au reste de la trompe qui a une section de forme ovale, cette partie a une section ellipsoïdale et un diamètre deux fois plus important. Cette caractéristique a été retrouvée chez deux autres spécimens de différents sexes et âges[6].
Fourrure

La fourrure est composée d'une couche extérieure de « poils de jarre », qui mesuraient 30 cm sur la partie supérieure du corps et jusqu'à 90 cm de long sur les flancs pour 0,5 mm de diamètre, et d'une couche dense de poils de bourre plus courts et légèrement bouclés mesurant 8 cm de long et 0,05 mm de diamètre. Les poils du haut de la jambe pouvaient atteindre 38 cm de long et ceux du pied mesuraient 15 cm de long, atteignant les orteils. Les poils de la tête étaient relativement courts mais plus longs sous le dessous et sur les côtés de la trompe. La queue était prolongée par des poils grossiers pouvant atteindre 60 cm de long, qui étaient plus épais que les poils de jarre. Vraisemblablement, les mammouths muaient saisonnièrement et la fourrure la plus lourde tombait au printemps. Comme les carcasses de mammouths retrouvées prises dans la glace l'ont certainement été à l'automne, les spécimens retrouvés portaient probablement tous la fourrure hivernale. Les éléphants modernes ont beaucoup moins de poils, bien que les jeunes aient une couverture pileuse un peu plus développée que les adultes[7]. Des comparaisons entre les poils des mammouths laineux et ceux des éléphants ont montré que leur morphologie était par contre relativement semblable[8].
Les Mammouths laineux avaient dans leur peau de nombreuses glandes sébacées qui sécrétaient des huiles dans leur fourrure, améliorant son isolation, évitant que l'eau ne la pénètre et lui donnant un aspect brillant[9].
La fourrure des Mammouths laineux préservés est brun-orangée, mais cela semble être lié à un blanchiment des pigments durant leur conservation. La quantité de pigments varie d'un poil à l'autre ainsi qu'à l'intérieur d'un même poil[5]. Une étude de 2006 a séquencé le gène MC1R (qui influence la couleur du poil chez les mammifères) à partir d'os de Mammouths laineux. Deux allèles ont été découverts : un dominant et l'autre récessif. Chez les mammifères, l'allèle récessif MC1R résulte en des poils clairs. Les mammouths qui naissaient hétérozygotes avec au moins un exemplaire de l'allèle dominant avaient des fourrures sombres, tandis que ceux qui avaient deux exemplaires de l'allèle récessif avaient des fourrures plus claires[10]. Une étude de 2011 a cependant montré que les individus clairs étaient rares[11]. En 2014, une autre étude a montré que la coloration d'un individu pouvait varier d'une dépigmentation des poils supérieurs, ou d'une robe bicolore mêlant poils de jarre rouge-brun et non pigmentés, à des robes avec les poils de bourre non pigmentés, donnant à chaque fois une coloration globale claire[12].
Denture
.jpg.webp)
Les Mammouths laineux avaient de très longues défenses, plus incurvées que celles des éléphants modernes. La plus grande défense jamais retrouvée mesurait 4,2 m de long pour un poids de 91 kg, mais en général elles mesuraient entre 2,4 et 2,7 m pour un poids de 45 kg. Les défenses des femelles étaient plus petites et plus fines, mesurant en moyenne entre 1,5 et 1,8 m pour un poids de 9 kg. Les fourreaux des défenses étaient parallèles et proches l'un de l'autre. Environ un quart de la longueur était situé à l'intérieur du fourreau. Les défenses croissaient en spirales, dans des directions opposées à leur base et se continuaient en formant une courbe jusqu'à ce que leurs extrémités pointent l'une en face de l'autre, se croisant parfois. De cette façon, la majeure partie du poids des défenses était proche du crâne, et il y avait moins de moment que si elles étaient droites. Les défenses étaient généralement asymétriques, de manière parfois importante, certaines défenses se développant parfois vers le bas plutôt que vers le haut, et certaines se cassant. Les jeunes développaient de petites défenses temporaires longues de quelques centimètres, qui étaient remplacées par des défenses permanentes un an plus tard. La croissance des défenses se poursuivait tout au long de la vie de l'animal, même si elle ralentissait lorsque celui-ci avait atteint l'âge adulte. Les défenses croissaient d'environ 2,5 à 15 cm chaque année. Certaines peintures rupestres représentent des Mammouths laineux avec de courtes défenses, voire sans défenses, mais on ne sait pas s'il s'agit d'une vue d'artiste ou d'une observation réelle. Les femelles Éléphants d'Asie n'ont pas de défenses, mais aucun fossile de mammouth n'a été retrouvé dépourvu de ces attributs[13] - [14].

Les Mammouths laineux avaient quatre molaires fonctionnelles à la fois, deux au niveau de la mâchoire supérieure et deux au niveau de la mâchoire inférieure. Environ 23 cm de leur couronne étaient à l'intérieur de la mâchoire et 2,5 cm en émergeaient. Les dents étaient continuellement poussées vers l'avant, de la même façon qu'un convoyeur à bande. Elles ont jusqu'à 26 crêtes distinctes d'émail, elles-mêmes couvertes de « prismes » dirigés vers la surface utilisée pour mâcher. Elles étaient relativement résistantes et rendues solidaires par le cément et la dentine. Un mammouth avait six ensembles de molaires durant sa vie, qui étaient remplacées cinq fois, mais quelques spécimens pouvaient avoir un septième ensemble de dents. Dans ce dernier cas, la durée de vie de l'animal pouvait être allongée. La première molaire était de la taille de celle d'un humain, 1,3 cm, la troisième mesurait 15 cm, de long, et la sixième environ 30 cm de long pour un poids de 1,8 kg. Les molaires grossissaient et présentaient plus de crêtes à chaque fois qu'elles étaient remplacées[15]. On considère que le Mammouth laineux avait les molaires les plus complexes de tous les éléphants[14].
Les déformations des molaires sont les problèmes de santé les mieux connus chez les fossiles de Mammouths laineux. Parfois, leur remplacement n'avait pas lieu convenablement et les molaires poussaient dans des positions anormales, toutefois, certains animaux semblent avoir pu survivre à cela. Des dents collectées en Grande-Bretagne ont montré que 2 % des spécimens présentaient une maladie parodontale, la moitié d'entre elles comportant des caries. Les dents avaient également parfois des croissances cancéreuses[16].
Paléobiologie
Comportement

Les Mammouths laineux adultes pouvaient efficacement se défendre des prédateurs avec leurs défenses, leur trompe et leur taille. Mais les jeunes et les adultes affaiblis étaient vulnérables à des animaux chassant en meute comme les loups, les humains, les Hyènes des cavernes et les grands félins. Les défenses étaient peut-être également utilisées pour des combats entre mammouths, pour défendre un territoire ou pour convoiter un partenaire. Les grandes défenses des mâles leur servaient ainsi peut-être à attirer les femelles et intimider leurs rivaux. Du fait de leur forme fortement incurvée, elles n'étaient pas adaptées pour empaler, mais plutôt pour frapper, comme le montrent certaines blessures retrouvées sur des omoplates de fossiles. Les très longs poils de sa queue permettaient certainement de compenser la petite taille de cette dernière, le rendant capable de s'en servir pour chasser les mouches, comme le font les éléphants modernes. Comme chez ces derniers, la trompe musculeuse et sensitive était utilisée comme un membre et avait diverses fonctions. Elle était utilisée pour manipuler des objets, et lors des interactions sociales[17]. Le pied bien préservé du mammouth mâle adulte de Yukagir a permis d'observer que la plante des pieds présentait plusieurs craquelures qui lui permettait de mieux s'accrocher aux surfaces lorsque l'animal marchait. Comme les éléphants modernes, les Mammouths laineux marchaient sur leurs orteils, et avaient de grands coussinets charnus sous les orteils[5].

Comme les éléphants modernes, les Mammouths laineux étaient vraisemblablement très sociaux et vivaient au sein de groupes familiaux matriarcaux. Cette théorie a été émise à partir de peintures rupestres représentant des groupes et de fossiles retrouvés ensemble. C'est pourquoi il est probable que le comportement social de ces animaux était proche de celui des éléphants modernes. On ne sait pas combien de mammouths vivaient au même endroit au même moment, car les dépôts de fossiles sont souvent des accumulations d'individus morts sur une longue période de temps. Ce nombre variait certainement selon la saison ou divers éléments du cycle de vie de ces animaux. Les éléphants modernes forment de grands troupeaux, parfois composés de plusieurs groupes familiaux, et ces troupeaux peuvent comporter des milliers d'animaux migrant ensemble. Les mammouths ont pu former de grands troupeaux très fréquemment, car ce comportement est souvent plus facile à mettre en œuvre dans des milieux ouverts que dans des milieux forestiers[18]. Des traces laissées par un troupeau de Mammouths laineux il y a entre 11 300 et 11 000 ans et retrouvées dans le Saint Mary Reservoir au Canada, ont montré que ce groupe comportait dans ce cas des nombres similaires d'individus adultes, subadultes et juvéniles. Les adultes faisaient des enjambées de 2 m, et les jeunes couraient pour rester au contact[19].

Le Mammouth laineux était probablement le membre le plus spécialisé de la famille des Elephantidae. Il avait une lipopexie (stockage de graisse) au niveau de son cou et de son garrot, en prévision de périodes où la disponibilité en nourriture serait insuffisante comme en hiver, et ses trois premières molaires croissaient plus rapidement que chez les jeunes éléphants modernes. On pense que l'expansion identifiée sur la trompe de « Yuka » et d'autres spécimens servait de « gant en fourrure » ; l'extrémité de la trompe n'était pas couverte de fourrure, mais était utilisée pour manger durant l'hiver, et aurait pu être réchauffée en étant glissée dans cette expansion. On imagine aussi que cette extension aurait pu être utilisée pour faire fondre de la neige afin d'approvisionner l'animal en eau, car la faire fondre directement dans sa gueule aurait pu perturber l'équilibre thermique de l'animal[6]. Comme chez le renne et le bœuf musqué, l'hémoglobine du Mammouth laineux était adaptée au froid, avec trois mutations permettant d'améliorer la libération de l'oxygène dans le corps et d'éviter le gel. Cette caractéristique aidait peut-être les mammouths à vivre à de hautes latitudes[20].
Dans une étude de 2015, des séquences de génome de très bonne qualité provenant de trois Éléphants d'Asie et de deux Mammouths laineux ont été comparées. Approximativement 1,4 million de différences au niveau des nucléotides d'ADN ont été trouvées entre mammouths et éléphants, qui affectent les séquences de plus de 1 600 protéines. Des différences ont été notées au niveau des gènes sur plusieurs aspects de physiologie et de biologie qui semblent liés à la survie dans l'Arctique, comme le développement de la peau et des poils, le stockage et le métabolisme du tissu adipeux et la perception de la température. Les gènes liés au ressenti de la température et la transmission de la sensation au cerveau sont modifiés. Un des gènes responsables de la sensibilité à la chaleur permet la production d'une protéine, TRPV3, trouvée dans la peau et qui affecte aussi la croissance des poils. Quand elle est insérée dans une cellule humaine, la version de la protéine présente chez le mammouth se révèle moins sensible à la chaleur que celle de l'éléphant. Par ailleurs, d'autres observations avaient montré que les souris qui n'avaient pas de TRPV3 active passaient plus de temps dans les endroits de la cage plus froids que les autres, et avaient une fourrure plus ondulée. Il y a également plusieurs altérations au niveau des gènes agissant sur l'horloge circadienne, peut-être pour mieux supporter les variations de la durée du jour extrêmes au niveau du pôle. Des mutations similaires sont connues chez d'autres mammifères arctiques, comme le renne[21] - [22].
Alimentation

De la nourriture à différents stades de digestion a été retrouvée dans les intestins de plusieurs Mammouths laineux, donnant une bonne idée de leur régime alimentaire. Les Mammouths laineux se nourrissaient de plantes, principalement de l'herbe et des carex, ainsi que de diverses autres plantes herbacées, plantes en fleurs, buissons, mousses et parties d'arbres. La composition exacte de leur régime alimentaire variait d'un endroit à l'autre. Les Mammouths laineux avaient besoin d'une diète diversifiée pour assurer leur croissance, comme les éléphants modernes. Un adulte de six tonnes devait avoir besoin de consommer 180 kg tous les jours, et devait s'alimenter durant 20 heures chaque jour. Les deux doigts de la trompe étaient probablement adaptés pour attraper les herbes courtes qui poussaient lors de la dernière ère glaciaire (glaciations quaternaires, il y a 2,58 millions d'années) en les enveloppant, tandis que les éléphants modernes enroulent leur trompe autour de la végétation des environnements tropicaux, nettement plus longue. La trompe pouvait également être utilisée pour arracher de grandes touffes d'herbe, cueillir délicatement des bourgeons et des fleurs, et arracher des feuilles et des branches quand des arbres ou des buissons étaient présents. Le « Mammouth du Yukagir » avait ingéré de la matière végétale contenant des spores de champignons coprophiles[23]. L'analyse des isotopes montre que les Mammouths laineux se nourrissaient principalement de plantes en C3, à la différence des chevaux et des rhinocéros[24].

Les scientifiques ont identifié du lait dans l'estomac et des matières fécales dans l'intestin du jeune mammouth « Lyuba »[25]. Les matières fécales ont peut-être été ingérées pour aider au développement de la flore microbienne nécessaire pour la digestion des végétaux, comme c'est le cas chez les éléphants modernes[26]. Une analyse des isotopes de Mammouths laineux du Yukon a montré que les jeunes étaient allaités durant trois ans, puis étaient sevrés. Leur régime alimentaire incluait progressivement les plantes à partir de l'âge de deux à trois ans. Ils étaient donc plus tardifs que chez les éléphants modernes, peut-être du fait du plus grand risque d'attaques par des prédateurs, ou de la difficulté de trouver de la nourriture durant les longues périodes d'hiver à des latitudes élevées[27].
Les molaires étaient adaptées à leur alimentation à base d'herbes grossières de la toundra, avec plus de plaques d'émail et une couronne plus grande que leurs ancêtres qui vivaient plus au sud. Le Mammouth laineux mâchait sa nourriture en utilisant les puissants muscles de ses mâchoires pour bouger ses mandibules vers l'avant en fermant la bouche puis vers l'arrière en l'ouvrant ; les arêtes émaillées aiguisées se rejoignaient et permettaient de broyer les aliments. Les arêtes étaient résistantes à l'usure pour permettre à l'animal de mâcher de grandes quantités de nourriture, qui parfois contenait du sable. Les Mammouths laineux utilisaient peut-être leurs défenses comme des pelles pour écarter la neige et trouver la végétation ensevelie dessous, et pour briser la glace pour pouvoir boire. Ceci est démontré par la présence sur des défenses bien conservées d'une section plate et polie d'environ 30 cm de long à l'endroit où les défenses devaient toucher terre. Celles-ci permettaient d'obtenir de la nourriture par d'autres biais, comme en déterrant des plantes ou en arrachant de l'écorce[28].
Cycle de vie

La longévité des mammifères est liée à leur taille. Puisque les éléphants modernes peuvent vivre jusqu'à 60 ans, on pense qu'il en était de même pour les Mammouths laineux qui étaient de taille similaire. L'âge d'un mammouth peut être déterminé en comptant les anneaux de croissance de ses défenses lorsque celles-ci sont sectionnées, mais cela ne fonctionne pas pour les premières années de l'animal, qui sont représentées par les extrémités des défenses, qui sont généralement usées. Dans la partie restante de la défense, chaque ligne majeure représente une année, et des lignes hebdomadaires et journalières peuvent être observées entre. Les bandes sombres correspondent aux étés, et il est donc possible de déterminer en quelle saison est mort un mammouth. La croissance des défenses ralentit quand il est plus difficile de s'alimenter, notamment l'hiver, lors d'une période de maladie ou quand un mâle a été banni d'un troupeau (les mâles éléphants vivent au sein du troupeau jusqu'à l'âge de dix ans). Les défenses des mammouths datant de la période la plus froide de la dernière ère glaciaire il y a 25 à 20 000 ans présentent des taux de croissance plus lents[29].
Les Mammouths laineux continuent leur croissance après avoir atteint l'âge adulte, comme les autres éléphants. Des os des membres non fusionnés ont montré que les mâles poursuivaient leur croissance jusqu'à l'âge de 40 ans, et les femelles jusqu'à 25 ans. Le jeune mammouth « Dima » retrouvé congelé mesurait 90 cm à sa mort à l'âge de 6 à 12 mois. À cet âge, la seconde série de molaires commençait à apparaître, et la première série était généralement usée à l'âge de 18 mois. La troisième série de molaires pouvait durer 10 ans, et le processus se renouvelait jusqu'à l'apparition du sixième et dernier jeu de molaires vers l'âge de 30 ans. Quand la dernière série de molaires était usée, l'animal devenait incapable de mâcher et ne pouvait donc plus se nourrir. Il mourrait alors rapidement de faim. Une étude sur les mammouths nord-américains a montré qu'ils mourraient souvent durant l'hiver ou au printemps, la période la plus dure pour la survie des animaux nordiques[29].

La tête la mieux conservée d'un spécimen de mammouth adulte est celle d'un mâle appelé « Mammouth du Yukagir ». Elle a montré que les Mammouths laineux disposaient de glandes temporales entre les oreilles et les yeux[30]. Ces caractéristiques indiquent que, comme les éléphants, les mâles Mammouths laineux entraient en « musth », une période durant laquelle ils étaient plus agressifs. Les glandes étaient utilisées en particulier par les mâles pour produire une substance huileuse à l'odeur très marquée, la frontaline. La fourrure permettait certainement de diffuser cette odeur plus longtemps[31].
L'examen approfondi de jeunes préservés a montré qu'ils étaient tous nés durant le printemps ou l'été. Les éléphants modernes ayant une gestation d'une durée de 21 à 22 mois, il est probable que la saison de reproduction avait lieu de l'été à l'automne[32]. L'analyse d'isotopes de δ15N d'une dent de « Lyuba » a montré son développement prénatal, et indiqué que sa durée de gestation était similaire à celle d'un éléphant moderne, et qu'elle était née au printemps[33].
Des traces de plusieurs maladies osseuses différentes ont été observées chez les Mammouths laineux. La plus commune de ces maladies était l'arthrose, que l'on rencontre chez 2 % des spécimens. Un spécimen de Suisse avait plusieurs vertèbres fusionnées à la suite de cette maladie. Le « Mammouth du Yukagir » souffrait de spondylodiscite dans deux vertèbres, et des cas d'ostéomyélite ont été observés sur plusieurs spécimens. D'autres présentaient des fractures guéries, montrant que ces animaux pouvaient survivre à ces blessures[34]. Un nombre de vertèbres cervicales anormal a été observé chez 33 % des spécimens de la région de la Mer du Nord, probablement du fait de la consanguinité dans une population déclinante[35]. Des mouches parasites et des protozoaires ont été identifiés dans l'intestin de « Dima »[36].
L'analyse en 2021 de la composition isotopique du strontium d'une défense d'un mammouth laineux mâle, longue de 1,7 m et vieille d'environ 17 100 ans, montre qu'au cours de sa vie (plus de 28 ans) il a parcouru presque 80 000 km à travers l'actuel Alaska, des déplacements sur des distances différentes selon les périodes de sa vie et qui se sont très fortement restreints pendant les derniers 18 mois, pendant lesquels il a souffert de famine (cause probable de sa mort)[37] - [38].
Distribution et habitat

L'habitat principal du Mammouth laineux était la steppe à mammouths ou steppe-toundra. Cet environnement se rencontrait à travers le nord de l'Asie, diverses parties de l'Europe et la partie septentrionale de l'Amérique du Nord durant la dernière ère glaciaire. Il se rapprochait des steppes herbeuses de la Russie actuelle, mais comportait une flore plus diverse, abondante, et qui poussait plus rapidement. Des herbes, des carex, des buissons étaient présents, et parfois quelques arbres isolés, notamment dans les régions les plus au sud. Cet habitat n'était pas dominé par la glace et la neige, comme on le croit communément, car ces régions étaient protégées par un anticyclone. Le Mammouth laineux partageait son habitat avec d'autres herbivores comme le Rhinocéros laineux, des chevaux sauvages et le bison[39]. Une étude de 2014 a conclu que les plantes herbacées non graminoïdes étaient plus importantes dans les steppes que ce que l'on pensait auparavant, et qu'elles représentaient une importante source de nourriture pour la mégafaune de l'ère glaciaire[40].
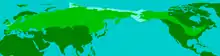
L'aire de répartition du Mammouth laineux correspondait donc à la présence de ces steppes à mammouths, et s'étendait sur toute l'Eurasie septentrionale et l'Amérique du Nord[41]. Le spécimen de Mammouth laineux trouvé le plus au sud était dans la province de Shandong en Chine et datait d'il y a 33 000 ans[42]. En Europe le spécimen le plus méridional a été retrouvé dans la dépression de Grenade en Espagne et datait de la même époque[43]. Le Mammouth laineux a également été ponctuellement présent au Japon il y a entre 20 000 ans et 40 000 ans. Leur répartition variait suivant les ères glaciaires. Lorsque le climat s'adoucissait, le Mammouth laineux gagnait les régions les plus septentrionales de l'Europe, comme les plaines scandinaves et la Sibérie, comme lors de la dernière période interglaciaire il y a entre 130 000 et 114 000 ans. Il redescendait dans l'Europe continentale, les parties plus méridionales de la Russie et le nord de la Chine lors des glaciations[41]. Durant la période du Dryas récent, les Mammouths laineux se sont brièvement étendus dans le nord-est de l'Europe, après quoi la population continentale s'est éteinte[44].
Originaire d'Eurasie, le Mammouth laineux a gagné l'Amérique du Nord via le détroit de Béring il y a 100 000 ans[41]. Les populations nord-américaines ont ainsi rejoint une autre espèce de mammouth, le Mammouth de Colomb, et des fossiles de ces deux espèces ont été trouvés ensemble dans quelques endroits d'Amérique du Nord, comme à proximité de l'actuelle Hot Springs dans le Dakota du Sud où leurs aires de répartition se superposaient. On ne sait pas si les deux espèces étaient sympatriques et vivaient ici simultanément, ou si les Mammouths laineux ne pénétraient dans ces terres que pendant les périodes où les Mammouths de Colomb n'y étaient pas[39]. Des travaux récents sur des isotopes stables chez des spécimens de mammouths de Sibérie et du Nouveau Monde ont montré qu'il y avait aussi des différences dans les conditions climatiques de chaque côté du détroit de Béring, la Sibérie étant plus uniformément froide et sèche tout au long du Pléistocène tardif[45]. Les deux populations ont divergé en deux sous-espèces distinctes, comme ont pu le montrer des études ADN récentes. Une analyse génétique de 2008 a montré que certains Mammouths laineux, qui étaient arrivés en Amérique du Nord en provenance d'Asie via le détroit de Bering, ont effectué une migration inverse il y a environ 60 000 ans à la faveur d'une période de refroidissement[41]. Après un temps de cohabitation ces mammouths nord-américains ont remplacé la population asiatique il y a 40 000 ans, peu de temps avant que l'espèce ne disparaisse complètement[46]. Cette version est confirmée par une étude ADN de 2008 qui a montré qu'il y avait deux groupes différents de Mammouths laineux : un qui s'est éteint il y a 45 000 ans et un autre il y a 12 000 ans. Les deux groupes semblent avoir suffisamment divergé pour qu'on les considère comme deux sous-espèces différentes. Le groupe qui s'est éteint le plus précocement vivait dans une zone limitée comprise entre les rivières Léna et Kolyma, dans le centre de la Sibérie Arctique, tandis que l'autre avait une aire de répartition plus étendue[47].
Taxonomie
Étymologie

À la suite de l'identification de Cuvier, Johann Friedrich Blumenbach donne au Mammouth laineux son nom scientifique, Elephas primigenius, en 1799, le plaçant dans le même genre que l'Éléphant d'Asie. Ce nom signifie en latin « premier éléphant ». Cuvier utilise le nom Elephas mammonteus quelques mois plus tard, mais c'est le premier nom qui est utilisé par la suite[48]. En 1828, Joshua Brookes utilise le nom Mammuthus borealis pour désigner les fossiles de Mammouths laineux de sa collection qu'il met en vente, créant ainsi un nouveau nom de genre[49]. On ne sait pas réellement d'où provient le mot « mammouth ». Selon l'Oxford English Dictionary, il viendrait d'un ancien mot mansi, mēmoŋt signifiant « corne de la terre »[50]. Cela pourrait être également une déclinaison de mehemot, le mot arabe pour désigner le « Béhémoth », monstre décrit dans le livre de Job[51]. Une autre origine possible est l'estonien, où maa signifie « terre », et mutt désigne la « taupe ». Le naturaliste Pallas appuie cette hypothèse sur le fait que certaines légendes sibériennes narraient l'existence d'une taupe géante vivant sous la terre, et que le parallèle aurait été fait avec les fossiles de mammouths retrouvés sous terre[51]. Le mot « mammouth » a été utilisé pour la première fois en Europe à la fin du XVIIe siècle, alors que les termes « mamant » ou « mamont » étaient jusque-là largement usités pour désigner les animaux auxquels appartenaient les fossiles découverts en Sibérie[51] - [52]. Thomas Jefferson, qui s'intéressait à la paléontologie, est en partie responsable de la transformation du mot mammouth d'un nom pour décrire les éléphants préhistoriques en un adjectif pour qualifier quelque chose de particulièrement grand. Ainsi la première utilisation connue du mot « mammouth » comme adjectif est une description d'un fromage (le « Cheshire Mammoth Cheese ») faite par Jefferson en 1802[53].
Classification

Des fossiles de diverses espèces d'éléphants disparues sont connus des Européens depuis des siècles, mais ils les voyaient, s'appuyant sur des récits bibliques, comme des restes de créatures légendaires comme le Béhémoth ou les géants. Certains faisaient également l'hypothèse qu'il s'agissait de fossiles d'éléphants modernes qui avaient été amenés en Europe durant la République romaine, comme les éléphants de guerre d'Hannibal et Pyrrhus Ier, ou d'animaux qui avaient divagué vers le nord[54].
Les premières traces fossilisées de Mammouth laineux étudiées par des scientifiques européens l'ont été par Hans Sloane en 1728 et comprenaient des dents et défenses venant de Sibérie. Sloane a été le premier à identifier que ces fossiles pouvaient provenir d'un éléphant[55]. Sloane trouva une explication biblique à la présence d'éléphants dans l'Arctique, considérant qu'ils avaient été ensevelis ici durant le Déluge, et que la Sibérie avait eu autrefois un climat tropical avant de grands changements climatiques[56]. D'autres auteurs ont interprété les conclusions de Sloane différemment, indiquant que le courant avait dû transporter les carcasses d'éléphants des tropiques vers l'Arctique. Les écrits de Sloane étaient basés sur des descriptions faites par des voyageurs et quelques os collectés en Sibérie et en Grande-Bretagne. Il discute avant tout sur la possibilité que ces restes soient ceux d'éléphants, mais ne tirent pas de conclusion claire[57]. En 1738, Johann Philipp Breyne indique que les fossiles de mammouths correspondent à une certaine forme d'éléphants. Cependant, il ne sait pas expliquer comment un animal tropical peut se rencontrer dans des régions froides comme la Sibérie, et suggère qu'ils ont dû y être transportés durant le Déluge[58]. En 1796, l'anatomiste français Georges Cuvier est le premier à identifier les fossiles de Mammouth laineux comme une nouvelle espèce, distincte des éléphants modernes. Il indique par ailleurs que cette espèce est éteinte et n'existe donc plus à l'heure actuelle, un concept qui n'est alors pas accepté par tous[54] - [59].
La taxonomie des éléphants disparus se complique au début du XXe siècle, et en 1942 un ouvrage posthume d'Henry Fairfield Osborn sur les Proboscidea est publié, dans lequel il utilise différents noms de taxon qui avaient auparavant été proposés pour les espèces de mammouths, et remplace notamment Mammuthus par Mammonteus, considérant que le premier nom n'avait pas été publié de manière valide[60]. Cette taxonomie est ensuite simplifiée par différents chercheurs au cours des années 1970, et toutes les espèces sont alors placées dans le genre Mammuthus et les écarts morphologiques notés pour différencier les espèces sont interprétés comme des variations intraspécifiques[61]. Osborn choisit deux molaires (trouvées en Sibérie et dans l'Osterode) de la collection de Blumenbach à l'université de Göttingen comme spécimen lectotype du Mammouth laineux, puisqu'aucun holotype n'avait été désigné du temps de Blumenbach. Véra Gromova a ensuite proposé que la première d'entre elles soit considérée comme le lectotype tandis que la suivante serait le paralectotype. Ces deux molaires ont été perdues dans les années 1980, et le « Mammouth de Taimyr », plus complet, découvert en Sibérie en 1948, a été proposé comme spécimen néotype en 1990. La validité du nom générique Mammuthus et la désignation de l'espèce type E. primigenius ont également été largement débattues à cette époque[62]. La molaire paralectotype (spécimen GZG.V.010.018) a depuis été retrouvée dans la collection de l'université de Göttingen, identifiée par comparaison avec l'illustration qu'Osborn en avait fait[48] - [63].
Évolution
Les premiers membres connus des Proboscidea, le clade qui comprend les éléphants modernes, existaient il y a environ 55 millions d'années autour de Téthys. Leurs plus proches parents sont les siréniens (dugongs et lamantins) et les hyracoïdes (un ordre de petits mammifères herbivores). La famille des Elephantidae existe depuis six millions d'années en Afrique et inclut les éléphants modernes et les mammouths. Parmi plusieurs clades aujourd'hui éteints, le Mastodonte (Mammut) est un lointain parent des mammouths, et fait partie d'une famille différente, celle des Mammutidae, qui a divergé 25 millions d'années avant que les mammouths n'évoluent[64]. Le cladogramme suivant montre le placement du genre Mammuthus parmi d'autres.
Phylogénie des genres d'éléphants ainsi que d'autres familles proches, d'après les caractéristiques de leur os hyoïde, selon Shoshani et al. (2007)[65] :
| Elephantimorpha |
| ||||||||||||||||||||||||||||||

En 2005, les chercheurs assemblent le profil complet du génome mitochondrial du Mammouth laineux, qui leur permet d'étudier les relations évolutives entre les mammouths et les Éléphants d'Asie (Elephas maximus)[66]. Une étude de 2015 sur cet ADN a confirmé que l'Éléphant d'Asie était le plus proche parent encore existant du Mammouth laineux[67]. L'Éléphant d'Afrique (Loxodonta africana) a divergé de ce clade il y a environ 6 millions d'années, soit à peu près au même moment que les chimpanzés ont divergé des Hommes. Avant la publication du génome de Neandertal, plusieurs chercheurs pensaient que le premier génome nucléaire d'une espèce éteinte complètement séquencé serait celui du mammouth[68]. Une étude de 2010 confirme ces relations, et suggère que les lignées des mammouths et des Éléphants d'Asie ont divergé il y a 5,8 à 7,8 millions d'années, tandis que les éléphants d'Afrique ont divergé d'un ancêtre commun il y a 6,6 à 8,8 millions d'années[69]. En 2008, la plupart de l'ADN chromosomique du Mammouth laineux était séquencé. Les analyses ont montré que le Mammouth laineux et l'éléphant d'Afrique avaient 98,55 % à 99,40 % de leur patrimoine génétique en commun[70]. Le génome nucléaire du Mammouth laineux a été séquencé à partir de l'ADN issu des follicules pileux d'un mammouth de 20 000 ans retrouvé dans le permafrost, et d'un autre mort il y a 60 000 ans[71]. En 2012, les protéines ont été identifiées de manière fiable pour la première fois, à partir d'un Mammouth laineux de 43 000 ans[72].

Grâce aux nombreuses découvertes de fossiles de mammouth, il est possible de reconstruire l'histoire évolutive de ce genre au travers d'études morphologiques. Les espèces de mammouths peuvent être identifiées à partir du nombre de crêtes émaillées (ou lamelles) sur leurs molaires : les espèces primitives ont peu de crêtes, et ce nombre augmente progressivement au fur et à mesure que les espèces ont évolué pour se nourrir d'aliments plus abrasifs. Dans un même temps, les couronnes des dents et les crânes deviennent plus profonds et la longueur du crâne mesurée du front à la nuque se réduit pour minimiser le poids de la tête[73] - [74]. Les crânes courts et hauts du Mammouth laineux et du Mammouth de Colomb (Mammuthus columbi) constituent le stade ultime de ce processus évolutif[75].
Les premiers membres connus du genre Mammuthus sont les espèces africaines Mammuthus subplanifrons qui vivait au Pliocène, et Mammuthus africanavus qui vivait au Pléistocène. La première était certainement un ancêtre de la seconde. Les mammouths sont arrivés en Europe il y a environ trois millions d'années. Le premier mammouth européen a été nommé Mammuthus rumanus ; il s'est répandu à travers l'Europe et jusqu'en Chine. Il n'est connu qu'à partir de ses molaires, qui présentent entre 8 et 10 crêtes émaillées. La population a évolué pour former une nouvelle espèce qui comptait 12 à 14 crêtes émaillées, devenant Mammuthus meridionalis, il y a environ 2 à 1,7 million d'années. Cette espèce a été à son tour remplacée par le Mammouth des steppes (Mammuthus trogontherii) avec 18 à 20 crêtes, qui a évolué dans l'est de l'Asie il y a environ 1 million d'années[73]. Le Mammouth de Colomb a évolué à partir d'une population de M. trogontherii qui a traversé le détroit de Béring et est entré en Amérique du Nord il y a environ 1,5 million d'années. Les mammouths dérivés de M. trogontherii ont développé des molaires avec 26 crêtes molaires il y a environ 400 000 ans en Sibérie pour devenir le Mammouth laineux. Les Mammouths laineux sont entrés en Amérique du Nord il y a environ 100 000 ans[75].
Des individus et des populations montrant des morphologies intermédiaires entre les espèces connues, ainsi que des espèces primitives et dérivées ont coexisté au moins jusqu'à ce que l'ancienne espèce disparaisse. Les différentes espèces et leurs formes intermédiaires ont été appelées « chrono-espèce ». Plusieurs taxons intermédiaires entre M. primigenius et les autres mammouths ont également été proposés, mais leur validité est incertaine. Suivant leur auteur, ils sont soit considérés comme des formes primitives d'une espèce évoluée ou comme les formes avancées d'une espèce primitive[73]. La distinction et la détermination de ces formes intermédiaires étaient considérées comme l'un des plus longs et plus complexes problèmes à résoudre pour la paléontologie du quaternaire. Des espèces régionales et intermédiaires incluant M. intermedius, M. chosaricus, M. p. primigenius, M. p. jatzkovi, M. p. sibiricus, M. p. fraasi, M. p. leith-adamsi, M. p. hydruntinus, M. p. astensis, M. p. americanus, M. p. compressus et M. p. alaskensis ont été reconnues[60] - [76] - [77].
Une étude de 2011 a placé deux spécimens de Mammouth de Colomb dans un sous-clade des Mammouths laineux. Cela laisse à penser que les deux populations pouvaient s'hybrider et donner une progéniture fertile. Un type nord-américain appelé à l'origine M. jeffersonii pourrait être un hybride entre ces deux espèces[78]. Une étude de 2015 suggère que les animaux qui vivaient dans les zones où M. columbi et M. primigenius cohabitaient formaient une métapopulation d'hybrides aux morphologies diverses. Elle suggère également que le M. primigenius eurasiatique avait des relations similaires avec M. trogontherii dans les zones où leurs aires de répartition se chevauchaient[79].
Relations avec l'Homme
Coexistence

Les humains ont coexisté avec le Mammouth laineux durant le Paléolithique supérieur, quand l'Homme est arrivé en Europe en provenance d'Afrique. Avant cela, l'Homme de Néandertal a coexisté avec les mammouths durant le Paléolithique moyen : ils ont notamment utilisé les os de mammouths pour faire des outils et construire des matériaux. Le Mammouth laineux était très important pour l'Homme durant l'ère glaciaire, car il était une source de nourriture, d'os, de graisse, de fourrure en quantités importantes[80]. Les preuves de cette coexistence ont été découvertes durant le XIXe siècle. William Buckland publie sa découverte du squelette de la dame rouge de Paviland en 1823, qui a été retrouvé dans une grotte avec des os de mammouths, mais dans un premier temps il ne considère pas qu'ils sont contemporains. En 1864, Édouard Lartet découvre une gravure d'un Mammouth laineux sur un morceau d'ivoire provenant de cet animal dans l'Abri de la Madeleine en Dordogne. Cette gravure est la première preuve largement acceptée de la coexistence de l'Homme avec des animaux préhistoriques éteints et est la première description contemporaine d'une telle créature pour la science moderne[81].

Le Mammouth laineux est la troisième espèce la plus fréquemment dépeinte dans les peintures de l'âge de glace, après les chevaux et les bisons, et ces peintures ont été réalisées entre 35 000 et 11 500 ans avant notre ère. Aujourd'hui, plus de 500 représentations de Mammouths laineux sont connues, sous la forme de peintures et gravures rupestres situées dans 46 grottes en Russie, France et Espagne, d'engravures et de sculptures (que l'on appelle « art mobilier ») dans de l'ivoire, du bois, de la pierre et des os. Les peintures rupestres de Mammouth laineux existent dans plusieurs styles et tailles différents. La grotte de Rouffignac en France présente le plus grand nombre de représentations (159) et certains des dessins mesurent plus de 2 m de long. D'autres grottes notables présentent des représentations de mammouths comme la grotte Chauvet, la grotte des Combarelles et Font-de-Gaume[82]. Une représentation dans la grotte du Castillo pourrait en fait représenter un Palaeoloxodon, l'« éléphant à défenses droites »[83].
L'art mobilier est plus facile à dater que les peintures dans des grottes car on les trouve dans les mêmes dépôts que des outils et d'autres objets datant de l'ère glaciaire. La plus grande collection d'objet d'art représentant des mammouths, comportant 62 représentations sur 47 plaques, a été découverte dans les années 1960 lors de fouilles sur le site de Gönnersdorf en Allemagne. Il ne semble pas y avoir de corrélation entre le nombre des mammouths dépeints et l'espèce qui était le plus fréquemment chassée, car les os de rennes sont les restes d'animaux les plus souvent retrouvés sur les sites de fouille. Deux propulseurs de lances en forme de Mammouths laineux ont également été retrouvés en France[82]. Certains objets n'ont pas été confectionnés là où ils ont été retrouvés, mais ont circulé par le biais du commerce de l'époque[83].
Utilisation par l'Homme
.JPG.webp)
Les os de Mammouths laineux ont été utilisés comme matériau de construction pour fabriquer des habitations par les Hommes de Néandertal comme les Hommes modernes durant l'ère glaciaire. Plus de 70 habitations de la sorte ont été retrouvées, principalement dans la plaine d'Europe orientale. La base de la hutte était circulaire, et avait une surface au sol de 8 à 24 m2. De gros os étaient utilisés pour constituer les fondations de la hutte, les défenses formaient l'entrée et le toit était certainement fait de peaux, tenues en place par des os et des défenses. Certaines huttes avaient un plancher qui allait 40 cm en dessous de la surface du sol. Certaines incluaient une place pour faire le feu, qui était alimenté avec des os, probablement du fait du manque de bois à cette période. Il est possible que certains des os utilisés comme matériaux provenaient de mammouths tués par l'Homme, mais l'état des os, et le fait que les os utilisés pour construire une même habitation pouvait avoir des milliers d'années de différence suggèrent qu'ils étaient souvent collectés dans les restes d'animaux morts depuis parfois longtemps. Les os de Mammouth laineux étaient également utilisés pour fabriquer divers outils, objets et instruments de musique. Les gros os, comme les omoplates, étaient également utilisés pour recouvrir les corps humain durant les funérailles[84].

L'ivoire du Mammouth laineux était utilisé pour créer des objets d'art. Plusieurs Vénus paléolithiques, dont la Dame de Brassempouy et la Vénus de Lespugue, ont été réalisées avec ce matériau. On a également retrouvé des armes faites en ivoire, comme des poignards, des lances et un boomerang. Pour pouvoir en travailler l'ivoire, les grandes défenses devaient être hachées, ciselées et divisées en morceaux plus petits, plus faciles à travailler. Certains objets en ivoire ont permis de voir que les défenses avaient été redressées, mais on ne sait pas comment[85].
Plusieurs spécimens de Mammouth laineux présentent des traces de dépouille par les hommes, par des cassures, des marques d'outils coupants et des marques d'outils en pierre. On ne sait pas à quel point la viande de mammouth était importante pour l'Homme préhistorique, car il y avait d'autres grands herbivores à disposition. Les Hommes pouvaient consommer des carcasses de mammouths retrouvés morts plutôt que les chasser. Certaines peintures rupestres montrent des mammouths dans des structures que l'on pourrait assimiler à des pièges à fosse. Quelques spécimens ont par contre montré des traces directes et claires de chasse par l'Homme. Un spécimen sibérien avec une pointe de lance restée coincée dans son omoplate prouve d'ailleurs que les lances pouvaient être lancées avec une grande force[86]. Un spécimen de la période du moustérien en Italie montre des traces de chasse à la lance par des Hommes de Néandertal[87]. Le jeune spécimen surnommé « Yuka » est le premier mammouth congelé pour lequel on a pu observer une interaction avec l'Homme. Il montre des traces montrant qu'il a été tué par un grand prédateur, et que des humains se sont nourris sur sa carcasse peu de temps après. Certains os ont été enlevés, et ont été trouvés non loin[88]. Près de la rivière Iana en Sibérie, on a découvert plusieurs spécimens avec des traces de chasse par l'Homme, mais il semble que ces animaux n'étaient pas chassés intensivement, mais plutôt lorsque les Hommes avaient des besoins d'ivoire[89]. Deux mammouths laineux du Wisconsin, les « Mammouths de Schaefer » et d'« Hebior », présentent des traces de découpe par des Paléoaméricains[90] - [91].
Place dans la culture
2.jpg.webp)
Le Mammouth laineux a gardé une place dans la culture importante longtemps après sa disparition. Les habitants indigènes de la Sibérie ont longtemps trouvé ce que l'on sait être aujourd'hui des fossiles de mammouths, et collecté les défenses pour le marché de l'ivoire. Les Sibériens pensaient que les fossiles de Mammouth laineux étaient ceux d'animaux ressemblant à des taupes géantes qui vivaient sous la surface du sol et mouraient quand on les mettait à la surface[92] - [93]. Les défenses de Mammouths laineux ont fait l'objet d'un commerce en Asie bien avant que les Européens n'en fassent connaissance. Güyük, le Khan des Mongols au XIIIe siècle, trônait selon la légende sur un fauteuil fait en ivoire de mammouth[94]. S'inspirant des croyances des indigènes sibériens, le mammouth est enregistré dans l'encyclopédie pharmaceutique chinoise, le Bencao gangmu, comme yin shu, le « rongeur caché »[95].
Les Amérindiens utilisaient également l'ivoire des Mammouths laineux ainsi que leurs os pour réaliser des outils et des objets d'art[96]. Comme en Sibérie, les indigènes nord-américains avaient des mythes concernant les fossiles de Mammouths laineux et autres éléphants ; les Iñupiat du détroit de Béring croyaient également que ces os venaient de créatures vivant sous le sol, tandis que d'autres peuples les associaient à des géants ou « grandes bêtes »[97] - [98] - [99]. Les légendes de plusieurs peuples natifs américains sont parfois interprétées comme comprenant des traditions orales sur les mammouths, mais cette hypothèse est décriée car il est très peu probable que les traditions orales traversent le temps de cette manière[97] - [99] - [100].

L'ivoire de mammouths sibériens est exporté vers la Russie et l'Europe dès le Xe siècle. Mais ce n'est qu'en 1611 que l'ivoire sibérien arrive en Europe de l'Ouest, lorsqu'une pièce est amenée à Londres. Quand la Russie occupa la Sibérie, le marché de l'ivoire augmenta et ce matériau devint largement exporté, de grandes quantités étant extraites du sous-sol sibérien. À partir du XIXe siècle, l'ivoire de mammouth devient un produit de luxe très onéreux, utilisé pour la confection de divers objets. Aujourd'hui il y a encore une demande importante en ivoire de mammouth pour remplacer celui des éléphants qu'il est désormais interdit de commercialiser, et on l'appelle parfois « or blanc ». Des commerçants locaux estiment qu'il y a encore 10 millions de mammouths congelés en Sibérie, et certains écologistes espèrent que cela va permettre de sauver les éléphants de l'extinction. Ces derniers sont chassés par des braconniers pour leur ivoire, mais si le marché peut être approvisionné avec de l'ivoire de mammouth, cela peut limiter le braconnage. Le marché de l'ivoire d'éléphant est interdit dans la majeure partie du monde à la suite de la conférence de Lausanne en 1989, mais de l'ivoire issu du braconnage rentre sur le marché sous le nom d'ivoire de mammouth. L'ivoire de mammouth ressemble très fortement à celui des éléphants, mais le premier est plus brun et les lignes de Schreger ont une texture plus rugueuse[101]. Au XXIe siècle, le réchauffement climatique rend accès plus facilement aux défenses en Sibérie, car le pergélisol fond plus rapidement, exposant les mammouths pris dedans[102]. Un trafic d'ivoire de mammouth a ainsi vu le jour, en particulier en Chine, légalement puisqu'il s'agit d'une espèce éteinte[103].
Le Mammouth laineux, symbole de puissance, est un élément important de la culture populaire moderne. Des millénaires après les Hommes préhistoriques, les artistes l'ont représenté dans leurs œuvres, d'abord de manière caricaturale et peu réaliste au XIXe siècle, comme dans les dessins de Boltunov, Golyschew et Édouard Riou, ou dans « La fuite devant le mammouth », une toile de Paul Jamin[104]. Au siècle suivant des représentations plus réalistes réalisées par des artistes spécialistes de la reconstitution d'animaux disparus comme Zdeněk Burian, Charles R. Knight ou Mauricio Antón permettent de vulgariser une image plus crédible de l'animal[104]. Dans la littérature, le clonage moderne du mammouth est évoqué dans le roman d'Henri Vernes Les Géants de la taïga, une aventure de Bob Morane[105]. Dans le roman La Guerre du feu, J.-H. Rosny aîné décrit « L’alliance entre l’Homme et le Mammouth » (titre d'un des chapitres)[106]. Au cinéma, un court métrage de Jacques Mitsch intitulé Le mammouth Pobalski relate l'histoire d'un explorateur se rendant dans une région imaginaire de la Sibérie orientale, l'Oubalski, à la recherche d'un Mammouth laineux. Il fera la connaissance de la population locale et découvrira ses coutumes[107]. Les mammouths apparaissent par ailleurs dans différents films, dont La Guerre du feu de Jean-Jacques Annaud, d'après le roman de J.-H. Rosny aîné, ou 10 000 de Roland Emmerich[104]. Le mammouth est un élément central d'un tableau du film de docufiction Homo sapiens de Jacques Malaterre. Le mammouth Manfred, dit Manny, est l'un des trois héros principaux de la série de films d'animation L'Âge de glace[104]. Dans la bande dessinée, cet animal apparaît par exemple dans un numéro de Rahan, fils des âges farouches, où il est chevauché par le héros[104]. Les Mammouths laineux constituent l'un des éléments centraux de l'histoire des jeux vidéo Syberia et Syberia 2[108], même s'ils n'apparaissent que très peu. Ils sont également présents dans les jeux Skyrim ou World of Warcraft. Ils sont également à l'origine du nom d'une chaîne de supermarché, Mammouth[104].
Histoire évolutive
| ||||||||||||||||||||||||||||||||||

Paléontologie
Spécimens fossilisés

Des fossiles de mammouths laineux ont été trouvés dans différents types de dépôts, comme d'anciennes rivières et lacs ou le « Doggerland » en mer du Nord, qui était à sec durant l'ère glaciaire. De tels fossiles sont généralement incomplets et ne comprennent pas de tissus. Les accumulations de squelettes d'éléphants modernes sont connues sous le nom de « cimetière des éléphants », car on pensait que les vieux éléphants se rendaient dans ces lieux précis pour mourir. Des accumulations similaires d'os de mammouths ont été retrouvées, et on pense qu'il s'agissait d'animaux morts près ou dans des rivières sur des milliers d'années, et dont les os étaient transportés par le courant. Certaines accumulations pourraient également être les restes de troupeaux morts au même moment, peut-être à cause de crues. Les pièges naturels, comme les kettles, les dolines ou la boue, ont également piégé des mammouths sur des périodes de temps étalées[109].

Mis à part le cas des animaux congelés, les seuls restes de tissus connus sont ceux d'un spécimen qui a été préservé dans un suintement de pétrole à Starunia, en Pologne. Les Mammouths laineux congelés ont été trouvés dans les parties les plus septentrionales de la Sibérie et de l'Alaska, avec beaucoup moins de découvertes dans ce dernier endroit. De telles découvertes sont généralement réalisées sous les terres du cercle Arctique, dans le pergélisol. Il apparaît que les tissus mous sont moins bien préservés sur des carcasses datant d'entre 30 000 et 15 000 ans avant notre ère, peut-être car le climat a été plus doux pendant cette période. La plupart des spécimens ont été partiellement dégradés avant leur découverte, du fait de leur exposition au soleil ou d'avoir été en partie consommés. Cette « momification naturelle » nécessite que l'animal soit très rapidement enseveli dans du liquide ou un matériel semi-solide comme de la cendre, de la boue ou de l'eau en partie gelée, et dans lequel il gèle[110].
La présence de nourriture non digérée dans l'estomac et de gousses de plantes encore dans la bouche de certains spécimens laisse à croire qu'ils ne sont probablement pas morts de faim ni de froid. La maturité de la végétation ingérée permet d'estimer la période de l'année à laquelle les animaux sont morts, généralement plutôt à l'automne qu'au printemps, quand on pourrait trouver des fleurs[111]. Ces animaux ont pu tomber à travers la glace dans de petits étangs et y rester. Certains sont certainement morts dans des rivières, peut-être emportés par des crues. Près de la rivière Berelekh en Yakutie en Sibérie, plus de 8 000 os provenant d'au moins 140 mammouths ont été trouvés en un seul endroit, apparemment emportés par le courant[112].
Mammouths conservés dans la glace

Entre 1692 et 1806, seulement quatre descriptions de mammouths congelés ont été recensées en Europe. Aucun élément de ces animaux n'a été préservé, et aucun squelette complet n'a été retrouvé durant cette période[94]. Tandis que les carcasses de Mammouths laineux congelées ont été extraites par les Européens dès 1728, le premier spécimen décrit en détail a été découvert dans le delta de la rivière Léna en 1799 par Ossip Schumachov, un chasseur sibérien[113]. Schumachov l'a laissé fondre jusqu'à ce qu'il puisse en retirer les défenses pour les vendre sur le marché de l'ivoire. En 1806, à Yakoutsk, Michael Friedrich Adams entend parler de cette découverte et se rend sur les lieux où il découvre que les carnivores sauvages avaient consommé la plus grande partie des organes et de la chair, dont la trompe. Il examine le mammouth et réalise que ce qu'il en reste constitue tout de même le plus complet découvert jusqu'alors. Mises à part les défenses déjà vendues, Adams récupère l'ensemble du squelette, une patte avant, la majeure partie de la peau et presque 18 kg de poils. Pendant son retour, il achète une paire de défenses qu'il pense être celles vendues par Shumachov. Adams apporte ces restes au musée zoologique de Saint-Pétersbourg où la tâche de reconstituer le squelette est confiée à Wilhelm Gottlieb Tilesius von Tilenau[57]. Le Kunstkamera, le musée établi par Pierre le Grand, contenait un squelette d'éléphant d'Asie qui pouvait être utilisé comme référence[114]. C'est l'une des premières tentatives de reconstitution de squelette d'un animal disparu. La reconstitution se révèlera correcte dans son ensemble, mais Tilesius inverse les défenses gauche et droite, qui se retrouvent ainsi incurvées vers l'extérieur et non vers l'intérieur. Cette erreur sera corrigée en 1899, et la place correcte des défenses de mammouths est encore sujet à débat au début du XXe siècle[115] - [116].

La découverte en 1901 du « mammouth de Berezovka » est la trouvaille récente la mieux documentée. Il a été exhumé des rives de la rivière Berezovka, affluent de la Kolyma, et les autorités russes ont financé les fouilles. Sa tête avait été exposée à l'air libre : la trompe et la peau avaient été mangées par les carnivores. L'animal avait encore de l'herbe entre les dents et sur la langue, montrant qu'il était mort subitement. L'expédition pour le ramener a duré 10 mois, et le spécimen a dû être coupé en morceaux avant d'être transporté vers Saint-Pétersbourg. L'animal a été identifié comme étant un mâle de 35 à 40 ans, mort il y a 35 000 ans. Une de ses omoplates était brisée, ce qui aurait pu arriver en tombant dans une crevasse[111].
En 1929, les restes de 34 mammouths avec des tissus mous congelés (peau, chair ou organes) avaient été recensés. Seulement quatre étaient relativement complets. Depuis, autant d'autres ont été découverts. Dans la plupart des cas, la chair présente des signes de putréfaction avant sa congélation et la dessiccation qui a suivi[117]. Depuis 1860, les autorités russes ont offert des récompenses pouvant atteindre 1 000 roubles pour la découverte de carcasses de mammouths laineux, car de telles découvertes étaient souvent gardées secrètes soit par superstition, soit par convoitise pour l'ivoire. Plusieurs carcasses ont ainsi été perdues car non signalées, et une a été mangée par des chiens. Plus récemment, des expéditions scientifiques ont été spécifiquement dédiées à la recherche de tels spécimens congelés, dont le plus célèbre est un jeune surnommé « Effie », qui a été trouvé en 1948. Il est composé d'une tête, d'une trompe et d'une patte avant, et date d'il y a 25 000 ans[109].
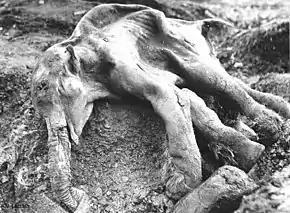
En 1977, la carcasse très bien préservée d'un mammouth laineux de sept à huit mois surnommé « Dima » a été extraite près d'un autre affluent de la Kolyma dans la région homonyme du nord-est de la Sibérie. « Dima » pesait approximativement 110 kg lors de sa mort et mesurait 104 cm de haut pour 115 cm de long. La datation par le carbone 14 situe sa mort il y a 40 000 ans. Ses organes internes sont similaires à ceux des éléphants modernes, mais ses oreilles sont plus petites que celles des éléphanteaux asiatiques du même âge. Un autre jeune moins complet, baptisé Macha, a été découvert dans la péninsule de Yamal en 1988. Il avait 3 à 4 mois, et une lacération sur sa patte droite qui pourrait avoir causé sa mort. C'est le mammouth congelé retrouvé le plus à l'ouest[118].
En 1997, le morceau d'une défense de mammouth est découvert dépassant de la toundra de la péninsule de Taïmyr en Sibérie. En 1999, cette carcasse de 20 380 ans et 25 tonnes de sédiments environnant sont transportées par un hélicoptère Mil Mi-26 vers une grotte glacée de Khatanga. Le spécimen est surnommé « mammouth de Jarkov ». En octobre 2000, les opérations minutieuses de décongélation dans cette grotte commencent par l'utilisation de sèche-cheveux pour garder intacts les poils et autres tissus[119] - [120].

En 2007, la carcasse d'une jeune femelle baptisée Liouba (« Aimée ») est découverte près de la rivière Youribeï, où elle avait été ensevelie durant 41 800 ans[26] - [121]. En coupant une section de molaire et étudiant ses anneaux de croissance, l'âge de l'animal a pu être estimé à un mois[33]. Le petit momifié pesait 50 kg, faisait 85 cm de haut et 130 cm de long[122] - [123]. Au moment de la découverte, ses yeux et sa trompe étaient intacts et une partie de la fourrure recouvrait encore son corps. Ses organes et sa peau étaient très bien conservés[124]. Liouba est certainement mort enseveli dans la vase d'une rivière que le troupeau traversait[26] - [125]. Après sa mort, son corps a pu être colonisé par des bactéries qui ont produit de l'acide lactique, qui a aidé à préserver le mammouth dans son état originel[26].

En 2012, un jeune a été retrouvé en Sibérie, avec des marques de découpe faites par l'Homme. Les scientifiques ont évalué son âge à environ 2,5 ans, et l'ont baptisé « Youka ». Son crâne et son pelvis avaint été retirés avant sa découverte, mais trouvés non loin[88] - [126]. Après sa découverte, la peau de « Youka » a été naturalisée[6].
En 2013, une carcasse bien préservée est découverte sur la Petite Liakhov, une des îles de l'archipel de Nouvelle-Sibérie. Il s'agissait d'une femelle âgée de 50 à 60 ans au moment de sa mort. La carcasse contenait des tissus musculaires bien préservés. Quand elle fut extraite de la glace, du sang liquide s'échappa de sa cavité abdominale. Ses découvreurs en conclurent que le sang du Mammouth laineux avait des propriétés anti-gel[127].
Recréation de l'espèce

L'existence de tissus conservés et d'ADN de mammouth laineux a permis d'imaginer de pouvoir récréer l'espèce avec l'aide de la science. Plusieurs méthodes ont été proposées pour arriver à cette fin. Le clonage pourrait par exemple être imaginé, en retirant le noyau d'un ovule d'une femelle éléphant, et la remplaçant par le noyau d'une cellule de tissu de mammouth laineux. La cellule devra ensuite être stimulée pour qu'elle se divise, et réinsérée dans l'appareil génital de la femelle éléphant. Le nouveau-né en résultant aurait ainsi le patrimoine génétique du mammouth laineux, même s'il se serait développé dans un environnement fœtal différent. Cependant les mammouths les mieux conservés avaient très peu d'ADN utilisable du fait de leur état de conservation. Il n'y en a pas suffisamment pour permettre de produire un embryon[128].
Une seconde méthode impliquerait d'inséminer artificiellement un ovule d'éléphant avec des cellules spermatiques d'une carcasse de mammouth laineux congelée. Le produit serait un hybride entre un éléphant et un mammouth, et le processus pourrait être répété afin d'obtenir plusieurs hybrides et les accoupler entre eux pour reproduire un animal proche du mammouth laineux originel. Cela est possible après sept générations de croisements entre ces hybrides. Le fait que les cellules spermatiques des mammifères modernes sont actives seulement 15 ans tout au plus après leur congélation est un obstacle à cette méthode[128]. Par ailleurs les croisements entre éléphants ne sont pas concluants, et dans un seul cas un éléphant d'Asie s'est accouplé avec un éléphant d'Afrique pour engendrer un éléphanteau baptisé « Motty », mais qui n'était pas viable et est mort avant d'atteindre l'âge de deux semaines[129].

En 2008, une équipe japonaise trouve de l'ADN utilisable dans des cerveaux de souris congelés durant 16 ans. Ils espèrent utiliser les mêmes méthodes pour trouver de l'ADN de mammouth utilisable[130]. En 2009, le bouquetin des Pyrénées (une sous-espèce du bouquetin ibérique) est le premier animal éteint à être cloné et revenir ainsi à la vie ; le clone ne vécut que 11 minutes avant de mourir d'une tare aux poumons[131]. Comme le génome du mammouth laineux a été complètement séquencé[132] - [133], il sera peut-être possible de recréer l'intégralité des chromosomes du mammouth laineux en ajoutant les séquences particulières aux mammouths dans les chromosomes d'un éléphant d'Asie, ce qui constitue une solution plus faisable[133] - [134], mais demeurant extrêmement difficile et onéreuse, pour recréer l'espèce. Si le processus se révèle fructueux, il y a un projet de réintroduire les mammouths laineux dans le parc du Pléistocène, une réserve naturelle de Sibérie[135].
En mars 2015, des gènes de mammouth laineux ont été copiés dans le génome d'un éléphant d'Asie, par le biais de la technique de la Clustered Regularly Interspaced Short Palindromic Repeats. Des segments génétiques issus de spécimens congelés, comprenant des gènes codant certaines caractéristiques des oreilles, de la graisse sous-cutanée et des poils, ont été introduits dans l'ADN de cellules de la peau d'un éléphant moderne. C'est la première fois que des gènes de mammouths laineux se retrouvent actifs depuis la disparition de l'espèce[136].
La recréation du mammouth laineux pourrait contribuer à lutter contre le réchauffement climatique : selon une expérience menée par Sergei et Nikita Zimov depuis les années 1980, sa réintroduction sur le pergélisol, ainsi que celle des animaux de l'époque du Pléistocène, pourrait faire baisser la température du pergélisol jusqu'à 9,4 °C, les grands herbivores encourageant la croissance des graminées des steppes, qui à leur tour ont un effet albédo élevé[103].
Les chercheurs s'interrogent néanmoins sur les questions d'éthique qui entourent la recréation d'une espèce. En plus des problèmes pratiques, il n'existe plus d'habitat convenable pour les mammouths laineux. Par ailleurs, pour une espèce sociale et grégaire, la création de quelques spécimens n'est pas idéale. Le temps et les ressources nécessaires à une telle entreprise seraient gigantesques, pour des bénéfices scientifiques incertains, et certains pensent que de telles ressources seraient plus utiles pour préserver les espèces d'éléphants existantes menacées[128] - [137]. L'aspect éthique de l'utilisation de femelle éléphant comme mère de clones de mammouths est également discuté, car la plupart des embryons ne survivront certainement pas, et on ne connaît pas réellement les besoins d'un nouveau-né mammouth[138].
Extinction
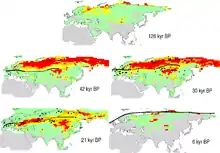
La plupart des populations de Mammouths laineux ont disparu durant la fin du Pléistocène et le début de l'Holocène, comme la plupart de la mégafaune du Pléistocène (dont le Mammouth de Colomb). Cette extinction fait partie d'une vague d'extinction massive connue comme l'extinction du quaternaire, qui a commencé 40 000 années avant et a culminé il y a 14 000 et 11 500 ans. Les avis scientifiques divergent quant à ses explications, attribuée à la chasse et/ou au changement climatique qui a entraîné une réduction de son habitat, sans que l'on sache lequel de ces facteurs a le plus pesé, ou si les deux ont été combinés. Quelle qu'en soit la cause, les grands mammifères sont généralement plus vulnérables que les petits car leurs populations sont moins importantes et leur taux de reproduction plus faible. Les populations de Mammouths laineux n'ont pas disparu simultanément, mais progressivement. La plupart a disparu il y a entre 14 000 et 10 000 ans. La dernière population continentale connue vivait dans la péninsule de Kyttyk en Sibérie il y a 9 650 ans[139] - [140]. Une petite population de Mammouths laineux a survécu sur l'île Saint-Paul, en Alaska, jusqu'à il y a 6 400 ans[141] - [142] - [143]. La toute dernière population connue a vécu isolée sur l'île Wrangel dans l'océan Arctique, jusqu'à il y a 4 000 ans, alors que la civilisation humaine était déjà avancée, et au moment de la construction de la pyramide de Khéops en Égypte antique[144] - [145] - [146] - [147].
_-_Skull_and_Lower_Jaw_A.JPG.webp)
L'ADN de restes de deux mammouths a été séquencé. L'un trouvé en Sibérie et daté d'il y a 44 800 ans et l'autre provenant de l'île Wrangel 4 300 ans avant notre ère. Ce travail a mis en évidence deux principaux temps de déclins de population : le premier remonte à 280 000 ans, auquel l'espèce a survécu, et un second il y a 12 000 ans, près du terme de l'ère glaciaire, qui a conduit à sa disparition[132].
Les mammouths de Wrangel se sont retrouvés isolés (durant 5 000 ans) par la montée du niveau de la mer. Du fait de la faible taille de la population en place quand l'île a été isolée, il y avait environ 300 à 1 000 individus sur l'île[148], impliquant une baisse de son hétérozygotie de l'ordre de 20[132] à 30 %[147] et une perte de diversité de l'ADN mitochondrial de l'ordre de 65 %[147]. La population semble pourtant être restée stable dans un premier temps, sans souffrir d'une baisse de diversité génétique supplémentaire[147] - [149].
Les études génétiques plaident pour une extinction ensuite soudaine, plutôt que résultant d'un déclin continu[147] ; par ailleurs la date de disparition correspond à peu près à l'arrivée de l'Homme sur l'île[150]. Les Mammouths laineux de l'est de la Béringie (Alaska et Yukon actuels) ont disparu plus tôt, il y a 13 300 ans, peu de temps après (environ 1 000 ans) la première apparition de l'Homme dans la région, et au même moment que le reste des Proboscidiens de la fin du Pléistocène (mammouths, gomphothères et mastodontes), ainsi que la plupart de la mégafaune de l'Amérique[151]. Par contre, les mammouths de l'île Saint-Paul ont apparemment disparu avant l'arrivée de l'Homme du fait de la réduction de leur habitat à la suite de la montée du niveau de la mer[151].
Une étude de 2008 a estimé que le changement climatique avait nettement réduit l'habitat convenant aux mammouths, passé de 7 700 000 km2 il y a 42 000 ans à 800 000 km2 il y a 6 000 ans[152] - [153]. Les Mammouths laineux avaient survécu à une diminution de leur habitat encore plus marquée à la fin de la glaciation du saalien il y a 125 000 ans, et il semble que l'Homme a chassé et conduit à leur extinction les populations persistantes à la fin de la dernière période glaciaire[154] - [155]. L'étude d'une piste datant d'il y a 11 300 à 11 000 ans dans le sud-ouest du Canada a montré que M. primigenius était en déclin tandis qu'il coexistait avec l'Homme, car il y avait moins de traces de jeunes que dans un troupeau normal[19].
Une étude récente (mars 2017, publiée par PLOS Genetics)[156] a comparé la qualité du génome insulaire à celui de mammouths continentaux vivant il y a 45 000 ans[157]. Le génome des individus insulaires contenait selon les généticiens un grand nombre de « mauvais gènes » (mutations délétères) et de codons anormaux susceptibles notamment d'avoir fait perdre à ces mammouths leur odorat, d’inhiber leur comportement sexuel et social[157]. Une mutation dans une partie du génome (FOXQ1) s’est probablement traduite par une fourrure de même couleur, mais brillante (satinée) et translucide[157].
Ces mutations apparues après que les mammouths aient déjà disparu du continent ne semblent pas uniquement dues à la consanguinité mais surtout à une moindre sélection naturelle au sein d’une population réduite[157]. Elles seront mieux comprises avec un plus grand nombre d’analyses génétiques, mais selon le paléontologue MacPhee[158], cette étude nous apprend peut-être quelque chose d'important sur ce qui se passe actuellement au sein des populations de mammifères menacées par ce type d’isolement et de raréfaction[157]. Selon lui ni la chasse humaine, ni le changement climatique ou tout autre facteur externe ne suffisait à lui seul à effacer le mammouth laineux de la planète, et il est important de comprendre comment le génome contribue ou non au risque d'extinction[157]. Mieux comprendre ces phénomènes pourrait avoir des implications pour la survie des éléphants (cousins des mammouths) et d’autres mammifères en danger de disparition[157]. Rogers[159] ajoutant qu'il vaut mieux empêcher une espèce d’entrer dans un processus de disparition que d'essayer d’ensuite retrouver sa diversité génétique car même s’il est possible de réaugmenter le nombre d'individus d’une population menacée, son génome pourra longtemps conserver les mutations délétères[157].
Les mammouths consommaient les bouleaux en entretenant un habitat herbeux. Le recul du mammouth a favorisé les forêts de bouleaux, qui absorbent plus la chaleur du soleil que les prairies, conduisant à un petit réchauffement de 0,2 C à de hautes latitudes de l'hémisphère nord selon une étude de 2010[160].
Cryptozoologie

On affirme occasionnellement que les mammouths n'auraient pas disparu. Ils pourraient survivre en de petits troupeaux isolés dans les toundras peu peuplées de l'hémisphère Nord. Au XIXe siècle, il y eut plusieurs témoignages de tribus sibériennes décrivant des créatures géantes et velues, qui n'ont pas été formellement confirmés. Un chargé d'affaires français travaillant à Vladivostok, M. Gallon, a dit en 1946 qu'il avait rencontré un trappeur russe en 1920 ayant affirmé avoir vu des éléphants géants et velus dans la taïga. Gallon a signalé aussi que ce trappeur ignorait auparavant l'existence des mammouths. En raison de la taille gigantesque de la Sibérie, on ne peut exclure totalement la possibilité de l´existence de ces créatures de nos jours, même si toutes les preuves tendent à indiquer qu'ils ont disparu il y a des milliers d'années. Il est probable que des cadavres de mammouths aient, seuls, permis aux tribus d'acquérir leurs connaissances sur ces animaux[161].
Il y a aussi des légendes autour de mammouths dans la culture amérindienne et esquimaude en Amérique du Nord et, au XIXe siècle, le bruit courait que des mammouths survivaient en Alaska[162]. En octobre 1899, Henry Tukeman a affirmé avoir tué un mammouth en Alaska, mais ce cas s'est révélé être un canular[163]. Bengt Sjögren pense que le mythe est apparu dans cette région lorsque le biologiste américain Charles Haskins Townsend a voyagé en Alaska, et rencontré des Eskimos qui vendaient des défenses de mammouths, et leur demanda s'il y avait encore des mammouths vivants en Alaska, leur montrant des représentations de l'animal[162].
Notes et références
Notes
- (en)/(de) Cet article est partiellement ou en totalité issu des articles intitulés en anglais « Woolly_mammoth » (voir la liste des auteurs) et en allemand « Wollhaarmammut » (voir la liste des auteurs).
Références
- Lister 2007, p. 82–83
- Lister 2007, p. 174–175
- (en) S. L. Vartanyan, K. A. Arslanov, J. A. Karhu, G. R. Possnert et L. D. Sulerzhitsky, « Collection of radiocarbon dates on the mammoths (Mammuthus primigenius) and other genera of Wrangel Island, northeast Siberia, Russia », Quaternary Research, vol. 70, no 1, , p. 51–59 (DOI 10.1016/j.yqres.2008.03.005)
- (en) N. Den Ouden, J. W. F. Reumer et L. W. Van Den Hoek Ostende, « Did mammoth end up a lilliput? Temporal body size trends in Late Pleistocene Mammoths, Mammuthus primigenius (Blumenbach, 1799) inferred from dental data », Quaternary International, vol. 255, , p. 53–58 (DOI 10.1016/j.quaint.2011.07.038, Bibcode 2012QuInt.255...53D)
- Lister 2007, p. 82–87
- (en) V. V. Plotnikov, E. N. Maschenko, I. S. Pavlov, A. V. Protopopov, G. G. Boeskorov et E. A. Petrova, « New data on trunk morphology in the woolly mammoth, Mammuthus primigenius (Blumenbach) », Paleontological Journal, vol. 49, no 2, , p. 200–210 (DOI 10.1134/S0031030115020070)
- Lister 2007, p. 83–84
- (en) A. Valente, « Hair structure of the Woolly mammoth, Mammuthus primigenius and the modern elephants, Elephas maximus and Loxodonta africana », Journal of Zoology, vol. 199, no 2, , p. 271–274 (DOI 10.1111/j.1469-7998.1983.tb02095.x)
- (en) V. E. Repin, O. S. Taranov, E. I. Ryabchikova, A. N. Tikhonov et V. G. Pugachev, « Sebaceous Glands of the Woolly Mammoth, Mammothus primigenius Blum.: Histological Evidence », Doklady Biological Sciences, vol. 398, , p. 382–384 (PMID 15587793, DOI 10.1023/B:DOBS.0000046662.43270.66)
- (en) H. Rompler, N. Rohland, C. Lalueza-Fox, E. Willerslev, T. Kuznetsova, G. Rabeder, J. Bertranpetit, T. Schöneberg et M. Hofreiter, « Nuclear Gene Indicates Coat-Color Polymorphism in Mammoths », Science, vol. 313, no 5783, , p. 62 (PMID 16825562, DOI 10.1126/science.1128994)
- (en) C. Workman, L. Dalen, S. Vartanyan, B. Shapiro, P. Kosintsev, A. Sher, A. Gotherstrom et I. Barnes, « Population-level genotyping of coat colour polymorphism in woolly mammoth (Mammuthus primigenius) », Quaternary Science Reviews, vol. 30, nos 17–18, , p. 2304–2308 (DOI 10.1016/j.quascirev.2010.08.020, Bibcode 2011QSRv...30.2304W)
- (en) Silvana R. Tridico, Paul Rigby, K. Paul Kirkbride, James Haile et Michael Bunce, « Megafaunal split ends: microscopical characterisation of hair structure and function in extinct woolly mammoth and woolly rhino », Quaternary Science Reviews, vol. 83, , p. 68–75 (DOI 10.1016/j.quascirev.2013.10.032, lire en ligne)
- Lister 2007, p. 94–95
- (en) B. Kurten et E. Anderson, Pleistocene Mammals of North America, New York, Columbia University Press, , 348–354 p. (ISBN 978-0-231-03733-4)
- Lister 2007, p. 92–93
- Lister 2007, p. 108–109
- Lister 2007, p. 95–105
- Lister 2007, p. 62–63
- (en) P. McNeil, L. Hills, B. Kooyman et S. Tolman, « Mammoth tracks indicate a declining Late Pleistocene population in southwestern Alberta, Canada », Quaternary Science Reviews, vol. 24, nos 10–11, , p. 1253–1259 (DOI 10.1016/j.quascirev.2004.08.019, Bibcode 2005QSRv...24.1253M)
- (en) K. L. Campbell, J. E. E. Roberts, L. N. Watson, J. R. Stetefeld, A. M. Sloan, A. V. Signore, J. W. Howatt, J. R. H. Tame, N. Rohland, T. J. Shen, J. J. Austin, M. Hofreiter, C. Ho, R. E. Weber et A. Cooper, « Substitutions in woolly mammoth hemoglobin confer biochemical properties adaptive for cold tolerance », Nature Genetics, vol. 42, no 6, , p. 536–540 (PMID 20436470, DOI 10.1038/ng.574)
- (en) V. Lynch, O. C. Bedoya-Reina, A. Ratan, M. Sulak, D. I. Drautz-Moses, G. H. Perry, W. Miller et S. C. Schuster, « Elephantid genomes reveal the molecular bases of Woolly Mammoth adaptations to the arctic », Cell Reports, (DOI 10.1101/018366)
- (en) Ewen Callaway, Nature magazine, « Mammoth Genomes Provide Recipe for Creating Arctic Elephants », scientificamerican.com,
- Lister 2007, p. 88–91
- (en) H. Bocherens, M. Fizet, A. Mariotti, R. A. Gangloff et J. A. Burns, « Contribution of isotopic biogeochemistry (13C,15N,18O) to the paleoecology of mammoths (Mammuthus primigenius) », Historical Biology, vol. 7, no 3, , p. 187–202 (DOI 10.1080/10292389409380453)
- (en) B. Van Geel, D. C. Fisher, A. N. Rountrey, J. Van Arkel, J. F. Duivenvoorden, A. M. Nieman, G. B. A. Van Reenen, A. N. Tikhonov, B. Buigues et B. Gravendeel, « Palaeo-environmental and dietary analysis of intestinal contents of a mammoth calf (Yamal Peninsula, northwest Siberia) », Quaternary Science Reviews, vol. 30, nos 27–28, , p. 3935–3946 (DOI 10.1016/j.quascirev.2011.10.009, Bibcode 2011QSRv...30.3935V)
- (en) D. C. Fisher, A. N. Tikhonov, P. A. Kosintsev, A. N. Rountrey, B. Buigues et J. Van Der Plicht, « Anatomy, death, and preservation of a woolly mammoth (Mammuthus primigenius) calf, Yamal Peninsula, northwest Siberia », Quaternary International, vol. 255, , p. 94–105 (DOI 10.1016/j.quaint.2011.05.040, Bibcode 2012QuInt.255...94F)
- (en) J. Z. Metcalfe, F. J. Longstaffe et G. D. Zazula, « Nursing, weaning, and tooth development in woolly mammoths from Old Crow, Yukon, Canada: Implications for Pleistocene extinctions », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 298, nos 3–4, , p. 257–270 (DOI 10.1016/j.palaeo.2010.09.032)
- Lister 2007, p. 92–95
- Lister 2007, p. 83–107
- (en) D. Mol, J. Shoshani, A. Tikhonov, B. van Geel, S. Sano, P. Lasarev et L. Agenbroad, « The Yukagir mammoth: brief history, 14c dates, individual age, gender, size, physical and environmental conditions and storage », Scientific Annals, School of Geology Aristotle University of Thessaloniki, vol. 98, , p. 299–314
- Lister 2007, p. 102–103
- Lister 2007, p. 104–105
- (en) A. N. Rountrey, D. C. Fisher, A. N. Tikhonov, P. A. Kosintsev, P. A. Lazarev, G. Boeskorov et B. Buigues, « Early tooth development, gestation, and season of birth in mammoths », Quaternary International, vol. 255, , p. 196–205 (DOI 10.1016/j.quaint.2011.06.006, Bibcode 2012QuInt.255..196R)
- Lister 2007, p. 108–111
- (en) J. W. F. Reumer, C. M. A. Ten Broek et F. Galis, « Extraordinary incidence of cervical ribs indicates vulnerable condition in Late Pleistocene mammoths », PeerJ, vol. 2, , e318 (PMID 24711969, PMCID 3970796, DOI 10.7717/peerj.318)
- Lister 2007, p. 87
- (en) Ariana Remmel, « Mammoth’s epic travels preserved in tusk », Nature, vol. 596, , p. 329- (DOI 10.1038/d41586-021-02206-1).
- (en) Matthew J. Wooller, Clement Bataille, Patrick Druckenmiller, Gregory M. Erickson, Pamela Groves et al., « Lifetime mobility of an Arctic woolly mammoth », Science, vol. 373, no 6556, , p. 806-808 (DOI 10.1126/science.abg1134).
- Lister 2007, p. 88–89
- (en) E. Willerslev, J. Davison, M. Moora, M. Zobel, E. Coissac, M. E. Edwards, E. D. Lorenzen, M. Vestergård, G. Gussarova, J. Haile, J. Craine, L. Gielly, S. Boessenkool, L. S. Epp, P. B. Pearman, R. Cheddadi, D. Murray, K. A. Bråthen, N. Yoccoz, H. Binney, C. Cruaud, P. Wincker, T. Goslar, I. G. Alsos, E. Bellemain, A. K. Brysting, R. Elven, J. R. H. Sønstebø, J. Murton et A. Sher, « Fifty thousand years of Arctic vegetation and megafaunal diet », Nature, vol. 506, no 7486, , p. 47–51 (PMID 24499916, DOI 10.1038/nature12921)
- Patou-Mathis 2015, Chapitre 12
- (en) K. Takahashi, G. Wei, H. Uno, M. Yoneda, C. Jin, C. Sun, S. Zhang et B. Zhong, « AMS 14C chronology of the world's southernmost woolly mammoth (Mammuthus primigenius Blum.) », Quaternary Science Reviews, vol. 26, nos 7–8, , p. 954–957 (DOI 10.1016/j.quascirev.2006.12.001, Bibcode 2007QSRv...26..954T)
- (en) D. J. Álvarez-Lao et N. García, « Comparative revision of the Iberian woolly mammoth (Mammuthus primigenius) record into a European context », Quaternary Science Reviews, vol. 32, , p. 64–74 (DOI 10.1016/j.quascirev.2011.11.004)
- (en) A. J. Stuart, « The extinction of woolly mammoth (Mammuthus primigenius) and straight-tusked elephant (Palaeoloxodon antiquus) in Europe », Quaternary International, vol. 126–128, , p. 171–177 (DOI 10.1016/j.quaint.2004.04.021, Bibcode 2005QuInt.126..171S)
- (en) P. Szpak, D. R. Gröcke, R. Debruyne, R. D. E. MacPhee, R. D. Guthrie, D. Froese, G. D. Zazula, W. P. Patterson et H. N. Poinar, « Regional differences in bone collagen δ13C and δ15N of Pleistocene mammoths: Implications for paleoecology of the mammoth steppe », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 286, , p. 88–96 (DOI 10.1016/j.palaeo.2009.12.009)
- (en) R. Debruyne, G. Chu, C. E. King, K. Bos, M. Kuch, C. Schwarz, P. Szpak, D. R. Gröcke, P. Matheus, G. Zazula, D. Guthrie, D. Froese, B. Buigues, C. De Marliave, C. Flemming, D. Poinar, D. Fisher, J. Southon, A. N. Tikhonov, R. D. E. MacPhee et H. N. Poinar, « Out of America: Ancient DNA Evidence for a New World Origin of Late Quaternary Woolly Mammoths », Current Biology, vol. 18, no 17, , p. 1320–1326 (PMID 18771918, DOI 10.1016/j.cub.2008.07.061)
- (en) M. T. P. Gilbert, D. I. Drautz, A. M. Lesk, S. Y. W. Ho, J. Qi, A. Ratan, C. -H. Hsu, A. Sher, L. Dalen, A. Gotherstrom, L. P. Tomsho, S. Rendulic, M. Packard, P. F. Campos, T. V. Kuznetsova, F. Shidlovskiy, A. Tikhonov, E. Willerslev, P. Iacumin, B. Buigues, P. G. P. Ericson, M. Germonpre, P. Kosintsev, V. Nikolaev, M. Nowak-Kemp, J. R. Knight, G. P. Irzyk, C. S. Perbost, K. M. Fredrikson et T. T. Harkins, « Intraspecific phylogenetic analysis of Siberian woolly mammoths using complete mitochondrial genomes », Proceedings of the National Academy of Sciences, vol. 105, no 24, , p. 8327–8332 (PMID 18541911, PMCID 2423413, DOI 10.1073/pnas.0802315105)
- (en) M. Reich, A. Gehler, D. Mohl, H. van der Plicht et A. M. Lister, « The rediscovery of type material of Mammuthus primigenius (Mammalia: Proboscidea) », International Mammoth Conference IV (poster), , p. 295.
- (en) J. Brookes, A catalogue of the anatomical & zoological museum of Joshua Brookes, vol. 1, Londres, Richard Taylor, (lire en ligne), p. 73
- (en) « Mammoth entry in Oxford English Dictionary »,
- Patou-Mathis 2015, p. 13-14
- Lister 2007, p. 49
- (en) J. Simpson, « Word Stories: Mammoth », sur Oxford English Dictionary Online, Oxford University Press, (consulté le )
- (en) B. Switek, Written in Stone : Evolution, the Fossil Record, and Our Place in Nature, Bellevue Literary Press, , 174–180 p. (ISBN 978-1-934137-29-1).
- (en) H. Sloane, « An Account of Elephants Teeth and Bones Found under Ground », Philosophical Transactions, vol. 35, nos 399–406, 1727–1728, p. 457–471 (DOI 10.1098/rstl.1727.0042, Bibcode 1727RSPT...35..457S).
- (en) H. Sloane, « Of Fossile Teeth and Bones of Elephants. Part the Second », Philosophical Transactions, vol. 35, nos 399–406, 1727–1728, p. 497–514 (DOI 10.1098/rstl.1727.0048, Bibcode 1727RSPT...35..497S).
- (en) The Academy of Natural Sciences, « Woolly Mammoth (Mammuthus primigenius) » [archive du ], The Academy of Natural Sciences, (consulté le ).
- (en) J. P. Breyne, T. s. et M. Wolochowicz, « A Letter from John Phil. Breyne, M. D. F. R. S. To Sir Hans Sloane, Bart. Pres. R. S. With Observations, and a Description of Some Mammoth's Bones Dug up in Siberia, Proving Them to Have Belonged to Elephants », Philosophical Transactions of the Royal Society of London, vol. 40, nos 445–451, , p. 124–138 (DOI 10.1098/rstl.1737.0026).
- G. Cuvier, « Mémoire sur les épèces d'elephans tant vivantes que fossils, lu à la séance publique de l'Institut National le 15 germinal, an IV », Magasin encyclopédique, 2e année, , p. 440–445.
- (en) H. F. Osborn, Proboscidea : A monograph of the discovery, evolution, migration and extinction of the mastodonts and elephants of the world, vol. 2, New York, J. Pierpont Morgan Fund, , 1116–1169 p. (lire en ligne).
- (en) V. J. Maglio, « Origin and evolution of the Elephantidae », Transactions of the American Philosophical Society, vol. 63, no 3, , p. 1–149 (DOI 10.2307/1379357, JSTOR 1379357).
- (en) W. E. Garutt, A. Gentry et A. M. Lister, « Mammuthus Brookes, 1828 (Mammalia, Proboscidea): proposed conservation, and Elephas primigenius Blumenbach, 1799 (currently Mammuthus primigenius): proposed designation as the type species of Mammuthus, and designation of a neotype », Bulletin of Zoological Nomenclature, vol. 47, , p. 38–44 (lire en ligne).
- (en) M. Reich et A. Gehler, « Giants' Bones and Unicorn Horns Ice Age Elephants Offer 21st Century Insights », Collections - Wisdom, Insight, Innovation, vol. 8, , p. 44–50.
- Lister 2007, p. 18–21
- [Shoshani et al. 2007] (en) Jeheskel Shoshani, Marco P. Ferretti, Adrian M. Lister, Larry D. Agenbroad, Haruo Saegusa, Dick Mol et Keiichi Takahashi, « Relationships within the Elephantinae using hyoid characters », Quaternary International, vol. 169-170, , p. 174 (DOI 10.1016/j.quaint.2007.02.003, Bibcode 2007QuInt.169..174S, lire en ligne [sur researchgate.net]).
- (en) L. Gross, « Reading the Evolutionary History of the Woolly Mammoth in Its Mitochondrial Genome », PLoS Biology, vol. 4, no 3, , e74 (PMID 20076539, PMCID 1360100, DOI 10.1371/journal.pbio.0040074)
- (en) Alfred L. Roca, Yasuko Ishida, Adam L. Brandt, Neal R. Benjamin, Kai Zhao et Nicholas J. Georgiadis, « Elephant Natural History: A Genomic Perspective », Annual Review of Animal Biosciences, vol. 3, no 1, , p. 139–167 (PMID 25493538, DOI 10.1146/annurev-animal-022114-110838, lire en ligne)
- (en) J. Krause, P. H. Dear, J. L. Pollack, M. Slatkin, H. Spriggs, I. Barnes, A. M. Lister, I. Ebersberger, S. Pääbo et M. Hofreiter, « Multiplex amplification of the mammoth mitochondrial genome and the evolution of Elephantidae », Nature, vol. 439, no 7077, , p. 724–727 (PMID 16362058, DOI 10.1038/nature04432)
- (en) N. Rohland, D. Reich, S. Mallick, M. Meyer, R. E. Green, N. J. Georgiadis, A. L. Roca et M. Hofreiter, « Genomic DNA Sequences from Mastodon and Woolly Mammoth Reveal Deep Speciation of Forest and Savanna Elephants », PLoS Biology, vol. 8, no 12, , e1000564 (PMID 21203580, PMCID 3006346, DOI 10.1371/journal.pbio.1000564)
- (en)Will findings recreate the woolly mammoth?, Pittsburgh Post-Gazette, 20 novembre 2008
- (en) « Woolly-Mammoth Genome Sequenced », Science Daily, (consulté le )
- (en) E. Cappellini, L. J. Jensen, D. Szklarczyk, A. L. Ginolhac, R. A. R. Da Fonseca, T. W. Stafford, S. R. Holen, M. J. Collins, L. Orlando, E. Willerslev, M. T. P. Gilbert et J. V. Olsen, « Proteomic analysis of a Pleistocene mammoth femur reveals more than one hundred ancient bone proteins », Journal of Proteome Research, vol. 11, no 2, , p. 917–926 (PMID 22103443, DOI 10.1021/pr200721u)
- (en) A. M. Lister, A. V. Sher, H. Van Essen et G. Wei, « The pattern and process of mammoth evolution in Eurasia », Quaternary International, vol. 126–128, , p. 49–64 (DOI 10.1016/j.quaint.2004.04.014, Bibcode 2005QuInt.126...49L)
- (en) M. P. Ferretti, « Structure and evolution of mammoth molar enamel », Acta Palaeontologica Polonica, vol. 48, , p. 383–396
- Lister 2007, p. 12–43
- (en) I. V. Foronova et A. N. Zudin, « Discreteness of evolution and variability in mammoth lineage: method for group study », The World of Elephants - Proceedings of the 1st International Congress, Rome, 2001, Rome, , p. 540–543 (lire en ligne)
- (en) I.V. Foronova, « Mammuthus intermedius (Proboscidea, Elephantidae) from the late Middle Pleistocene of the southern Western and Central Siberia, Russia: the problem of intermediate elements in the mammoth lineage », Russian Journal of Theriology, vol. 13, , p. 71–82 (lire en ligne)
- (en) J. Enk, A. Devault, R. Debruyne, C. E. King, T. Treangen, D. O'Rourke, S. L. Salzberg, D. Fisher, R. MacPhee et H. Poinar, « Complete Columbian mammoth mitogenome suggests interbreeding with woolly mammoths », Genome Biology, vol. 12, no 5, , R51 (PMID 21627792, PMCID 3219973, DOI 10.1186/gb-2011-12-5-r51)
- (en) A. M. Lister et A. V. Sher, « Evolution and dispersal of mammoths across the Northern Hemisphere », Science, vol. 350, no 6262, , p. 805–809 (PMID 26564853, DOI 10.1126/science.aac5660)
- « Le mammouth » (consulté le )
- Lister 2007, p. 116–117
- Lister 2007, p. 118–125
- (en) I. M. Braun et M. R. Palombo, « Mammuthus primigenius in the cave and portable art: An overview with a short account on the elephant fossil record in Southern Europe during the last glacial », Quaternary International, vol. 276-277, , p. 61–76 (DOI 10.1016/j.quaint.2012.07.010)
- Lister 2007, p. 128–132
- Lister 2007, p. 131–137
- Lister 2007, p. 151–155
- (en) M. Mussi et P. Villa, « Single carcass of Mammuthus primigenius with lithic artifacts in the Upper Pleistocene of northern Italy », Journal of Archaeological Science, vol. 35, no 9, , p. 2606–2613 (DOI 10.1016/j.jas.2008.04.014)
- (en) B. Aviss, « Woolly mammoth carcass may have been cut into by humans », BBC, (lire en ligne, consulté le )
- (en) P. Nikolskiy et V. Pitulko, « Evidence from the Yana Palaeolithic site, Arctic Siberia, yields clues to the riddle of mammoth hunting », Journal of Archaeological Science, vol. 40, no 12, , p. 4189–4197 (DOI 10.1016/j.jas.2013.05.020)
- (en) D. F. Overstreet et M. F. Kolb, « Geoarchaeological contexts for Late Pleistocene archaeological sites with human-modified woolly mammoth remains in southeastern Wisconsin, U.S.A », Geoarchaeology, vol. 18, , p. 91–114 (DOI 10.1002/gea.10052)
- (en) D. J. Joyce, « Chronology and new research on the Schaefer mammoth (?Mammuthus primigenius) site, Kenosha County, Wisconsin, USA », Quaternary International, vol. 142-143, , p. 44–57 (DOI 10.1016/j.quaint.2005.03.004)
- (en) Raymond Lee Newcomb, « Our lost explorers : the narrative of the Jeannette Arctic Expedition as related by the survivors, and in the records and last journals of Lieutenant De Long »,
- (en) S. Patkanov, « Die lrtysch-Ostjaken und ihre Volkspoesie », St. Petersburg, , p. 123–124
- (en) I. P. Tolmachoff, « The carcasses of the mammoth and rhinoceros found in the frozen ground of Siberia », Transactions of the American Philosophical Society, vol. 23, no 1, , p. 11–23 (DOI 10.2307/1005437, JSTOR 1005437)
- (en) Berthold Laufer, « Arabic and Chinese Trade in Walrus and Narwhal Ivory », T'oung Pao, Second Series, vol. 14, no 3, , p. 329 (JSTOR 4526349). La source de Bertholds pour la croyance Khantys est Patkanov 1897, p. 123–124
- (en) C. Cohen, The Fate of the Mammoth : Fossils, Myth, and History, University of Chicago Press, , 197–198 p. (ISBN 0-226-11292-6, lire en ligne)
- (en) W. D. Strong, « North American Indian traditions suggesting a knowledge of the mammoth », American Anthropologist, vol. 36, , p. 81–88 (DOI 10.1525/aa.1934.36.1.02a00060)
- (en) G. E. Lankford, « Pleistocene Animals in Folk Memory », The Journal of American Folklore, vol. 93, no 369, , p. 294–296 (DOI 10.2307/540573, JSTOR 540573) (inscription requise)
- (en) A. Mayor, Fossil Legends of the First Americans, Princeton, Princeton University Press, , 446 p. (ISBN 0-691-11345-9), p. 97
- (en) G. E. Lankford, « Pleistocene Animals in Folk Memory », The Journal of American Folklore, vol. 93, no 369, , p. 293–304 (DOI 10.2307/540573, JSTOR 540573) (inscription requise)
- Lister 2007, p. 137–139
- (en) B. Larmer, « Mammoth Tusk Hunters », nationalgeographic.com,
- « Le mammouth pourrait être ressuscité pour lutter contre le réchauffement climatique », sur National Geographic.fr, (consulté le ).
- Patou-Mathis 2015, p. Avant-propos
- « Les Géants de la taïga » sur le site NooSFere (consulté le ).
- « La Guerre du feu » sur le site NooSFere (consulté le ).
- « Oubalski, le Mammouth Pobalski » (consulté le )
- Patou-Mathis 2015, p. Des mammouths et des Hommes
- Lister 2007, p. 45–75
- Lister 2007, p. 50–51
- (en) E. W. Pfizenmayer, Siberian Man and Mammoth, Londres, Blackie and Son, , 46–61 p.
- (en) N. K. Vereshchagin, « The mammoth "cemeteries" of north-east Siberia », Polar Record, vol. 17, no 106, , p. 3 (DOI 10.1017/S0032247400031296)
- (en) M. Adams, « Some Account of a Journey to the Frozen-Sea, and of the Discovery of the Remains of a Mammoth », The Philadelphia Medical and Physical Journal, vol. 3, , p. 120–137
- (la) W. G. Tilesio, « De skeleto mammonteo Sibirico ad maris glacialis littora anno 1807 effosso, cui praemissae Elephantini generis specierum distinctiones », Mémoires de l'Académie Impériale des Sciences de St. Pétersbourg, vol. 5, , p. 406–514
- (en) C. Cohen, The Fate of the Mammoth : Fossils, Myth, and History, Chicago, University of Chicago Press, , 297 p. (ISBN 978-0-226-11292-3, lire en ligne), p. 113
- (en) E. Pfizenmayer, « A Contribution to the Morphology of the Mammoth, Elephas Primigenius Blumenbach; With an Explanation of My Attempt at a Restoration », Annual Report of the Board of Regents of the Smithsonian Institution, , p. 326–334
- (en) W. R. Farrand, « Frozen Mammoths and Modern Geology: The death of the giants can be explained as a hazard of tundra life, without evoking catastrophic events », Science, vol. 133, no 3455, , p. 729–735 (PMID 17777646, DOI 10.1126/science.133.3455.729, Bibcode 1961Sci...133..729F)
- Lister 2007, p. 57–58
- (en)Mol, D. et al. (2001). "The Jarkov Mammoth: 20,000-Year-Old carcass of a Siberian woolly mammoth Mammuthus primigenius (Blumenbach, 1799)". The World of Elephants, Proceedings of the 1st International Congress (16–20 octobre 2001, Rome): 305–309. Full text pdf
- (en) R. G. Debruyne, V. R. Barriel et Pascal Tassy, « Mitochondrial cytochrome b of the Lyakhov mammoth (Proboscidea, Mammalia): New data and phylogenetic analyses of Elephantidae », Molecular Phylogenetics and Evolution, vol. 26, no 3, , p. 421–434 (PMID 12644401, DOI 10.1016/S1055-7903(02)00292-0)
- (en) P. A. Kosintsev, E. G. Lapteva, S. S. Trofimova, O. G. Zanina, A. N. Tikhonov et J. Van Der Plicht, « Environmental reconstruction inferred from the intestinal contents of the Yamal baby mammoth Lyuba (Mammuthus primigenius Blumenbach, 1799) », Quaternary International, vol. 255, , p. 231–238 (DOI 10.1016/j.quaint.2011.03.027, Bibcode 2012QuInt.255..231K)
- (en) P. Rincon, « Baby mammoth discovery unveiled », BBC News, (lire en ligne, consulté le )
- (en) D. Solovyov, « Baby mammoth find promises breakthrough », Reuters, (lire en ligne, consulté le )
- (en) O. Smith, « Baby mammoth Lyuba, pristinely preserved, offers scientists rare look into mysteries of Ice Age », Daily News, New York, (lire en ligne)
- (en) Daniel C. Fisher, « X-ray computed tomography of two mammoth calf mummies », Journal of Paleontology, vol. 88, no 4, , p. 664–675 (DOI 10.1666/13-092)
- (en) E. N. Mashchenko, A. V. Protopopov, V. V. Plotnikov et I. S. Pavlov, « Specific characters of the mammoth calf (Mammuthus primigenius) from the Khroma River (Yakutia) », Biology Bulletin, vol. 40, no 7, , p. 626–641 (DOI 10.1134/S1062359013070042)
- (en) K. Wong, « Can a mammoth carcass really preserve flowing blood and possibly live cells? », Nature, (DOI 10.1038/nature.2013.13103)
- Lister 2007, p. 42–43
- (en) R. Stone, « Cloning the Woolly Mammoth », Discover Magazine, (lire en ligne)
- (en) « Cloned Mammoths Made More Likely by Frozen Mice », FOX News, (lire en ligne, consulté le )
- (en) « Extinct ibex is resurrected by cloning », Telegraph Online, (lire en ligne, consulté le )
- (en) E. Palkopoulou, S. Mallick, P. Skoglund, J. Enk, N. Rohland, H. Li, A. Omrak, S. Vartanyan, H. Poinar, A. Götherström, D. Reich et L. Dalén, « Complete Genomes Reveal Signatures of Demographic and Genetic Declines in the Woolly Mammoth », Current Biology, vol. 25, , p. 1395–1400 (DOI 10.1016/j.cub.2015.04.007, lire en ligne)
- (en) P. Ghosh, « Mammoth genome sequence completed », BBC, (consulté le )
- (en) « Scientists Sequence Woolly-Mammoth Genome » [archive du ], Mammoth Genome Project, Pennsylvania State University (consulté le )
- (en) S. A. Zimov, « ESSAYS ON SCIENCE AND SOCIETY: Pleistocene Park: Return of the Mammoth's Ecosystem », Science, vol. 308, no 5723, , p. 796–798 (PMID 15879196, DOI 10.1126/science.1113442)
- (en) Sarah Fecht, « Woolly Mammoth DNA Successfully Spliced Into Elephant Cells », Popular Science,
- (en) A. Griffin, « Woolly mammoth could be revived after scientists paste DNA into elephant's genetic code », The Independent,
- (en) Pasqualino Loi, Joseph Saragusty et Grazyna Ptak, « Cloning the Mammoth: A Complicated Task or Just a Dream? », Adv Exp Med Biol, vol. 753, , p. 489–502 (PMID 25091921, DOI 10.1007/978-1-4939-0820-2_19)
- Lister 2007, p. 146–148
- (en) A. J. Stuart, L. D. Sulerzhitsky, L. A. Orlova, Y. V. Kuzmin et A. M. Lister, « The latest woolly mammoths (Mammuthus primigenius Blumenbach) in Europe and Asia: A review of the current evidence », Quaternary Science Reviews, vol. 21, nos 14–15, , p. 1559–1569 (DOI 10.1016/S0277-3791(02)00026-4)
- (en) R. Dale Guthrie, « Radiocarbon evidence of mid-Holocene mammoths stranded on an Alaskan Bering Sea island », Nature, vol. 429, no 6993, , p. 746–749 (PMID 15201907, DOI 10.1038/nature02612)
- (en) D. R. Yesner, D. W. Veltre, K. J. Crossen et R. W. Graham, « 5,700-year-old Mammoth Remains from Qagnax Cave, Pribilof Islands, Alaska », Second World of Elephants Congress, (Hot Springs: Mammoth Site), , p. 200–203
- (en) K. S. Crossen, « 5,700-Year-Old Mammoth Remains from the Pribilof Islands, Alaska: Last Outpost of North America Megafauna », Geological Society of America, vol. 37, , p. 463 (lire en ligne)
- (en) Markus Milligan, « Mammoths still walked the earth when the Great Pyramid was being built » (consulté le )
- (en) A. J. Stuart, P. A. Kosintsev, T. F. G. Higham et A. M. Lister, « Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth », Nature, vol. 431, no 7009, , p. 684–689 (PMID 15470427, DOI 10.1038/nature02890)
- (en) S. L. Vartanyan et al., « Radiocarbon Dating Evidence for Mammoths on Wrangel Island, Arctic Ocean, until 2000 BC », Radiocarbon, vol. 37, no 1, , p. 1–6 (ISSN 0033-8222, lire en ligne)
- (en) V. Nyström, J. Humphrey, P. Skoglund, N. J. McKeown, S. Vartanyan, P. W. Shaw, K. Lidén, M. Jakobsson, I. A. N. Barnes, A. Angerbjörn, A. Lister et L. Dalén, « Microsatellite genotyping reveals end-Pleistocene decline in mammoth autosomal genetic variation », Molecular Ecology, vol. 21, no 14, , p. 3391–3402 (PMID 22443459, DOI 10.1111/j.1365-294X.2012.05525.x)
- (en) W. Dunham, « Lonely end for the world's last woolly mammoths », sur ABC Science, Reuters, (consulté le )
- (en) V. Nystrom, L. Dalen, S. Vartanyan, K. Liden, N. Ryman et A. Angerbjorn, « Temporal genetic change in the last remaining population of woolly mammoth », Proceedings of the Royal Society B: Biological Sciences, vol. 277, no 1692, , p. 2331–2337 (DOI 10.1098/rspb.2010.0301)
- (en) R. E. Ackerman, « Early maritime traditions in the Bering, Chukchi, and East Siberian seas », Arctic Anthropology, vol. 35, no 1, , p. 247–262 (JSTOR 40316468)
- (en) Stuart Fiedel, American Megafaunal Extinctions at the End of the Pleistocene, Springer, , 21–37 p. (ISBN 978-1-4020-8792-9, DOI 10.1007/978-1-4020-8793-6_2, lire en ligne)
- (en) D. Nogués-Bravo, J. S. Rodríguez, J. N. Hortal, P. Batra et M. B. Araújo, « Climate Change, Humans, and the Extinction of the Woolly Mammoth », PLoS Biology, vol. 6, no 4, , e79 (PMID 18384234, PMCID 2276529, DOI 10.1371/journal.pbio.0060079)
- (en) C. Sedwick, « What Killed the Woolly Mammoth? », PLoS Biology, vol. 6, no 4, , e99 (PMID 20076709, PMCID 2276526, DOI 10.1371/journal.pbio.0060099)
- (en) P. S Martin, Twilight of the mammoths : ice age extinctions and the rewilding of America, Berkeley, University of California Press, , 165–173 p. (ISBN 0-520-23141-4)
- (en) D. Burney et T. Flannery, « Fifty millennia of catastrophic extinctions after human contact », Trends in Ecology & Evolution, vol. 20, no 7, , p. 395–401 (PMID 16701402, DOI 10.1016/j.tree.2005.04.022)
- Rebekah L. Rogers & Montgomery Slatkin (2017), Excess of genomic defects in a woolly mammoth on Wrangel island ; PLOS le 2 mars 2017 ; https://dx.doi.org/10.1371/journal.pgen.1006601
- Brian Switek (2017), Dying woolly mammoths were in ‘genetic meltdown’ Genomic study of extinction in progress may offer insights for modern conservation ; 02 March 2017, doi:10.1038/nature.2017.21575 (résumé)
- Ross MacPhee, paléotologue à l'American Museum of Natural History de New York City
- Rebekah Rogers (University of California, Berkeley), co-auteur avec Montgomery Slatkin de l'étude,
- (en) C. E. Doughty, A. Wolf et C. B. Field, « Biophysical feedbacks between the Pleistocene megafauna extinction and climate: the first human-induced global warming? », Geophysical Research Letters, vol. 37, no 15, (DOI 10.1029/2010GL043985, Bibcode 2010GeoRL..3715703D)
- Lister 2007, p. 55
- DOI 10.1525/aa.1934.36.1.02a00060
- (en) M. Murray, « Henry Tukeman: Mammoth's Roar was Heard All The Way to the Smithsonian » [archive du ], Tacoma Public Library, (consulté le )
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- Claude Guérin, « Superordre des Proboscidiens », dans Les grands mammifères plio-pléistocènes d'Europe, Masson, coll. « Préhistoire », Paris, 1996, (ISBN 2-225-84951-X)
- (en) A. Lister et P. Bahn, Mammoths : Giants of the Ice Age, Londres, Frances Lincoln, , 192 p. (ISBN 978-0-520-26160-0).

- Marylène Patou-Mathis, Histoires de mammouth, Fayard, coll. « Divers Histoire », , 392 p. (ISBN 978-2-213-68336-2 et 2-213-68336-0, lire en ligne).
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :
Références taxonomiques
- (en) Référence NCBI : Mammuthus primigenius (taxons inclus) (consulté le )
- (en) Référence Tree of Life Web Project : Mammuthus primigenius (consulté le )
- (en) Référence Paleobiology Database : Mammuthus primigenius Blumenbach 1799 (éteint) (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023