Edmontosaurus
Edmontosaurus regalis


- † Claosaurus annectens
Marsh, 1892 - † Thespesius saskatchewanensis
Sternberg, 1926 - † Anatotitian copei
Lull & Wright, 1942
Edmontosaurus est un genre éteint de grands dinosaures herbivores « à bec de canard » du sous-ordre des ornithopodes et de la famille des hadrosauridés. Ses fossiles ont été retrouvés dans des sédiments de l'ouest de l'Amérique du Nord datant de la fin du Campanien jusqu'à la fin du Maastrichtien au Crétacé supérieur, il y a entre 73 et 66 millions d'années. C'est l'un des derniers dinosaures non-aviens à avoir vécu aux côtés de Triceratops et de Tyrannosaurus peu de temps avant l'extinction Crétacé-Tertiaire.
Edmontosaurus est l'un des plus grands hadrosauridés. Il mesurait 13 mètres de long et pesait environ 4 tonnes. Il est connu par plusieurs spécimens bien conservés qui comprennent non seulement des os mais aussi, dans certains cas, des empreintes cutanées étendues et un possible reste de bol digestif.
Edmontosaurus a une histoire taxonomique longue et compliquée remontant à la fin du XIXe siècle. Diverses espèces classées autrefois dans des genres tels que Claosaurus, Thespesius, Trachodon, Anatotitan et Anatosaurus sont maintenant considérés comme appartenant au genre Edmontosaurus. Les premiers fossiles de ce genre ont été découverts dans le sud de l'Alberta, au Canada, dans ce qu'on appelait autrefois la formation d'Edmonton. L'espèce type, E. regalis, a été nommée par Lawrence Lambe en 1917[1], bien que plusieurs autres espèces qui sont maintenant classées dans le genre Edmontosaurus aient été nommées plus tôt. La plus connue d'entre elles est E. annectens, initialement nommée par Othniel Charles Marsh en 1892, Claosaurus annectens puis connue pendant de nombreuses années sous le nom de Anatosaurus annectens. On connait aussi une troisième espèce plus petite, E. saskatchewanensis. Le nom Edmontosaurus qui signifie « lézard d'Edmonton », a été créé d'après la formation d'Edmonton, aujourd'hui connue sous le nom de formation de Horseshoe Canyon.
Edmontosaurus avait une large répartition dans l'Ouest de l'Amérique du Nord. La répartition des fossiles suggère qu'il préférait vivre sur les côtes et les plaines côtières. C'était un herbivore qui pouvait se déplacer soit sur deux soit sur quatre pattes. Retrouvé dans plusieurs gisements, Edmontosaurus semble avoir vécu en groupe et pourrait avoir été migrateur. La richesse de ses fossiles a permis aux paléontologues d'étudier en détail sa paléobiologie, notamment son cerveau, son mode d'alimentation, ses pathologies et blessures, dont une attaque de tyrannosaure sur un edmontosaure.
Description
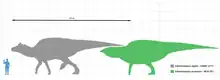
Edmontosaurus a été décrit en détail à partir de plusieurs spécimens[2] - [3] - [4] - [5]. Comme d'autres hadrosauridés, c'était un animal massif avec une longue queue aplatie transversalement et une tête terminée par une sorte de « bec de canard ». La tête n'avait pas de crêtes creuses ou pleines, contrairement à beaucoup d'autres hadrosauridés. Les pattes avant n'étaient pas aussi fortes que les pattes arrière, mais étaient assez longues pour être utilisées pour l'appui ou la marche. Edmontosaurus figurait parmi les plus grands hadrosauridés : un adulte atteignait en moyenne environ 9 mètres de long et quelques-uns des plus grands spécimens ont même atteint 12[6] à 13 mètres de long[7]. Son poids est estimé avoir été de l'ordre de 4,0 tonnes[8]. E. regalis était la plus grande espèce connue mais son rang a été contesté car le grand hadrosauridé Anatotitan copei est considéré comme un Edmontosaurus annectens, comme l'ont proposé Jack Horner et ses collègues dès 2004[8] - [9] - [10]. On estime que le spécimen type de E. regalis, (NMC 2288), mesurait 9 à 12 mètres de long[11]. E. annectens était légèrement plus court. Deux squelettes montés, USNM 2414 et YPM 2182, mesurent respectivement 8,00 et 8,92 mètres de longueur[11] - [12]. Cependant, il y a au moins une publication parlant d'un possible E. annectens beaucoup plus grand, de près de 12 mètres de long[13]. E. saskatchewanensis était plus petit avec une longueur totale estimée entre 7,0 et 7,3 mètres[14].
Tête

La tête d'un Edmontosaurus adulte mesurait environ un mètre de long, E. regalis ayant la plus longue du genre et E. annectens la plus courte[11]. Elle était de profil à peu près triangulaire[2], sans crête osseuse à son sommet[14]. Vue de dessus, l'avant et l'arrière étaient plus larges que la partie centrale, avec la partie antérieure constituée d'un large bec en forme de bec de canard ou de cuillère. Ce bec était édenté, et ses extrémités supérieure et inférieure terminées par une partie kératinitisée[8]. On a retrouvé d'importants restes du bec supérieur en kératine de Mummy, le fossile conservé au muséum Senckenberg à Francfort-sur-le-Main[6]. Pour ce fossile, la partie non-osseuse du bec dépassait l'os d'au moins 8 centimètres de long[15]. Les narines d’Edmontosaurus étaient allongées et logées dans de profondes dépressions entourées de fortes bordures osseuses en dessus, en arrière et en dessous[16]. Dans un cas au moins (le spécimen de Senckenberg), les anneaux sclérotiques ont été retrouvés dans les cavités orbitaires[17]. Un autre os, rarement retrouvé, l'étrier (un os de l'oreille des reptiliens), a également été retrouvé dans un fossile d’Edmontosaurus[8].
Reste du corps

Edmontosaurus possédait des membres antérieurs courts, une queue longue et pointue, des pieds pourvus de trois orteils. Sa tête était plate et se terminait par un bec dépourvu de dents. Il mastiquait sa nourriture grâce à près d'un millier de petites dents jugales. Sa taille pouvait atteindre 13 mètres pour un poids de 3,5 tonnes. Ses vertèbres possédaient des petites protubérances courtes et plates, qui devaient dépasser de sa peau telles des épines, allant de la queue à la tête, en passant par tout le dos.
On pense que ce dinosaure était quadrupède et bipède en même temps, comme tous les ornithopodes. Il devait passer la plupart du temps à quatre pattes, se nourrissant de plantes basses, mais il pouvait se relever et courir à deux pattes, pour éviter un prédateur par exemple. Si les représentations anciennes montraient un animal lourd, pataud et lent, des récentes études biomécaniques ont démontré qu'il pouvait courir à plus de 50 km/h, soit plus vite qu'un Tyrannosaurus rex. On pense aujourd'hui qu'il s'agissait d'un animal svelte et agile.
Bien qu'il fût quasiment dépourvu de défense, il possédait des sens très développés (vue, ouïe et odorat), lui permettant d'éviter de rencontrer trop souvent ses prédateurs. Sa peau était constituée de cuir. Il est possible qu'il ait été pourvu d'une poche cutanée à proximité du nez. Certains paléontologues pensent qu'il pouvait la gonfler comme un ballon, peut-être comme un rituel destiné à affirmer la suprématie d'un mâle à l'intérieur de la horde, ou pour communiquer.
On ne trouvait des dents que sur les maxillaires et les dentaires (os principal de la mâchoire inférieure). Les dents étaient sans cesse remplacées, demandant environ six mois pour se développer entièrement[18]. Elles poussaient en colonnes, avec un maximum de six dents en formation par colonne, le nombre de colonnes variant selon la taille de l'animal[5]. Le nombre de colonnes pour les différentes espèces sont les suivants: 51 à 53 colonnes pour le maxillaire et 48 à 49 pour le dentaire (les dents de la mâchoire supérieure étant légèrement plus étroites que celles de la mâchoire inférieure) pour E. regalis, 43 colonnes par maxillaire et 36 par dentaire pour E. annectens et respectivement 52 et 44 pour E. saskatchewanensis[14].
Le nombre de vertèbres diffère selon les spécimens. E. regalis avait treize vertèbres cervicales, dix-huit vertèbres thoraciques, neuf vertèbres lombaires et un nombre inconnu de vertèbres caudales[14]. Un spécimen autrefois considéré comme appartenant à Anatosaurus edmontoni (maintenant attribué à E. annectens) est signalé comme ayant une vertèbre surnuméraire au niveau du dos et 85 vertèbres caudales, mais avec taux de restauration non divulgué[14]. On considérait que les autres hadrosauridés avaient 50 à 70 vertèbres caudales[8], mais cela semble avoir été surestimé. Le début du dos remontait d'avant en arrière, relié en avant au cou plié vers le haut alors que le sommet du reste du dos et de la queue étaient horizontaux[8]. La plus grande partie du dos et de la queue était renforcée par des tendons ossifiés disposés en treillis le long des apophyses épineuses des vertèbres. Cela permettait que le dos et au moins une partie de la queue restent droits[19] - [20]. On interprète le rôle de ces tendons ossifiés comme servant à renforcer la colonne vertébrale contre les effets de la pesanteur, sachant que ce gigantesque animal avait une colonne vertébrale horizontale supportée essentiellement par les pattes arrière et les hanches[19].

Les omoplates étaient de longues lames plates, à peu près parallèles à la colonne vertébrale. Les hanches étaient composées de trois éléments : un ilion allongé au-dessus de l'articulation de la hanche, un ischion en dessous se prolongeant en arrière par une longue tige mince, et un pubis orienté en avant qui s'étalait en une structure en forme de plaque. Cette structure de la hanche empêchait l'animal de se redresser, car dans une telle position le fémur aurait poussé conjointement sur l'ilion et le pubis, au lieu de pousser seulement sur l'ilion plus solide. Les neuf vertèbres lombaires étaient fusionnées renforçant la solidité de la hanche[5].
Les pattes avant étaient plus courtes et moins lourdement construites que les pattes arrière. L'humérus avait une grande crête delto-pectorale servant à la fixation des muscles, tandis que le cubitus et le radius étaient relativement minces. Le bras et l'avant-bras étaient sensiblement de la même longueur. Le poignet était simple, avec seulement deux petits os. Chaque main avait quatre doigts, sans pouce. Les deuxième, troisième et quatrième doigts avaient à peu près la même longueur et étaient réunis dans une enveloppe charnue. Les deuxième et troisième doigts se terminaient par des sortes d'ongles qui étaient également recouverts de peau et n'étaient pas apparents. Le petit doigt, écarté des trois autres, était beaucoup plus court. Les fémurs étaient robustes et droits, avec une importante crête vers le milieu de la face postérieure[5]. Cette crête servait à la fixation des muscles puissants allant des hanches à la queue et qui tiraient les cuisses (et donc les pattes arrière) vers l'arrière et contribuaient à faire de la queue un important organe d'équilibre[21]. Chaque pied avaient trois doigts, sans gros orteil ou petit orteil. Les orteils se terminaient par des sortes de sabots[5].
Revêtement cutané

On a retrouvé de multiples spécimens d’Edmontosaurus avec leurs empreintes cutanées conservées. Plusieurs de ces découvertes ont été très médiatisées, comme celle de la « momie Trachodon » au début du xxe siècle[22] - [23] et du spécimen « Dakota »[24] - [25] - [26] celui-ci possédant encore semble-t-il des composés organiques cutanés[26]. C'est grâce à ces découvertes, que le dessin des écailles d’Edmontosaurus est connu pour la plupart des parties du corps.
On admet maintenant qu’AMNH 5060, la « momie Trachodon » (appelée ainsi parce qu'elle ressemble à une momie), est un spécimen d’E. annectens. On a retrouvé des empreintes cutanées du museau, d'une grande partie du cou et du thorax, et de certaines parties des pattes avant et arrière[15]. La queue et une partie des pattes arrière avaient été emportées par l'érosion avant la découverte, de sorte que ces zones ne sont pas connues chez ce spécimen[23]. En outre, certaines zones avec des reliefs cutanés, tels que les articles composant la crête du cou (voir ci-dessous) et les doigts, ont été accidentellement détruits lors de la préparation de l'animal[22]. On pense que l'animal s'est desséché après être mort de faim et de soif dans un lit de rivière à sec[23], probablement au niveau ou à proximité d'un point d'eau à sec[27] - [28]. La carcasse desséchée aurait ensuite été enterrée lors d'une inondation soudaine, recouverte de particules suffisamment fines pour en faire un moulage des structures épidermiques[23].

L'épiderme était mince et composé de petites écailles ne se chevauchant pas[22], comme on peut le voir de nos jours chez le monstre de Gila[15]. Deux types d'écailles étaient présents sur la plus grande partie du corps : de petits tubercules pointus ou convexes, de 1 à 3 millimètres de diamètre, désordonnés (écailles de fond), et des tubercules plus grands, plats, polygonaux (écailles structurelles) faisant généralement moins de 5 millimètres de diamètre, mais atteignant jusqu'à 10 millimètres au niveau de la partie inférieure des pattes avant. Les écailles structurelles étaient regroupées en plaques séparées par des écailles de fond, avec des écailles transitionnelles entre les deux. Sur la plus grande partie du corps, les écailles structurelles étaient disposées en plaques circulaires ou ovales, tandis que sur le haut des pattes avant, elles formaient des bandes à peu près parallèles entre elles et à l'omoplate. En règle générale, les plaques étaient plus grandes sur la partie supérieure du corps et plus petites sur la face inférieure. On a retrouvé des plaques allant jusqu'à 50 centimètres de longueur au-dessus des hanches[23].

Les seules impressions cutanées de la tête viennent de la région des narines. Au lieu de traces d'écailles, on trouve des marques de tissus mous pliés, avec une zone plus épaisse située à l'extrémité antérieure de l'ouverture de ce qui doit avoir été l'emplacement approximatif des narines elles-mêmes[15]. Le cou et le dos portaient une crête souple courant le long de la ligne médiane, avec une rangée de plaques ovales posées au-dessus des apophyses épineuses des vertèbres. La hauteur totale de cette crête n'est pas connue chez AMNH 5060, ni le dessin de son bord supérieur, car l'extrémité supérieure a été perdue. La crête mesurait au moins 8 centimètres de hauteur, et était plissée pour permettre les mouvements. Osborn a supposé qu'elle était assez grande pour disposer d'une autre rangée de plaques[23].
La partie inférieure des pattes avant portait les plus grandes écailles, disposées en une seule grande plaque couvrant les principales régions. Les extrémités des pattes antérieures étaient couvertes de petites écailles structurelles et des tissus mous dans les espaces interdigitaux des trois doigts centraux, ne libérant même pas les extrémités des doigts. Osborn a interprété cela comme un système de pagaie utilisé par l'animal lors de ses baignades[23]. Robert T. Bakker plus tard, leur a donné un autre rôle celui de tampon en tissus mous pour amortir la marche, analogue à celui des chameaux[29]. Comme au niveau des pattes avant, la partie inférieure des pattes arrière avait de grosses écailles. On ne connait rien actuellement du revêtement cutané du reste de la jambe, bien que des impressions sur un spécimen de Lambeosaurus, un lambeosauriné, suggèrent que la peau des cuisses avaient la même que le corps, comme les oiseaux modernes[15].
La queue de l’AMNH 5060 est absente, mais d'autres spécimens ont fourni des détails relatifs à cette zone. Des impressions cutanées d'une partie de queue appartenant à l'un de ses proches parents, un Anatotitan, retrouvé dans la formation de Hell Creek au Montana, montrent une crête au-dessus des vertèbres. La crête mesurait environ 8,0 centimètres de hauteur, faite de segments d'environ 5,0 centimètres de longueur et 4,5 centimètres de hauteur, espacés de 1,0 cm, chaque segment correspondant à une vertèbre[30]. Une autre queue, cette fois appartenant à un jeune E. annectens possédait des impressions cutanées fossilisées avec des écailles ainsi que des textures cutanées originales. Ces impressions comprenaient des écailles imbriquées elliptiques, des écailles rainurées et une espèce de corne trapézoïdale de 9 sur 10 cm2[31].
Classification
Cladogrammes successifs des Hadrosauridae (ou Hadrosaurinae) avec la position d'Edmontosaurus.
D’après Horner et al. (2004)[8] :
D’après Gates et Sampson (2007)[32] :
D’après Paul Godefroit et al. (2012)[33] :
-
- Bactrosaurus
- Hadrosauridae
- Hadrosaurinae
- Saurolophidae
- Lambeosaurinae
- Saurolophinae
- Wulagasaurus
-
- Brachylophosaurini
-
- Kritosaurus
-
-
- Saurolophini
- Edmontosaurini
-
- Edmontosaurus annectens
- Edmontosaurus regalis
D’après Albert Prieto-Márquez et al. (2016)[9] :
-
- Telmatosaurus
-
-
- Claosaurus
-
- Tethyshadros
- Hadrosauridae
- Hadrosaurus
-
- Eotrachodon
- Saurolophidae
- Lambeosaurinae
- Saurolophinae
-
-
- Naashoibitosaurus
-
- Kritosaurus
-
- Gryposaurus
-
- Big Bend UTEP 37.7
-
- « Sabinosaure » PASAC-1
-
Edmontosaurus est un hadrosauridé, une famille de dinosaures qui à ce jour n'est connue que pour avoir vécu au Crétacé supérieur. Il est classé dans les Saurolophinae (ex-Hadrosaurinae), un clade des hadrosauridés qui n'avait pas de crêtes creuses. Les autres membres du groupe comprennent Brachylophosaurus, Gryposaurus, Lophorhothon, Maiasaura, Naashoibitosaurus, Prosaurolophus et Saurolophus[8]. Un autre grand hadrosauridé des dernières formations du Crétacé de l'ouest de l'Amérique du Nord, Anatotitan copei, est aujourd'hui considéré comme un synonyme junior d'E. annectens[34] - [8] - [35] - [33] - [9] - [10]. Le saurolophiné chinois géant Shantungosaurus est le plus proche (groupe frère) d'Edmontosaurus[36] - [9].
Bien que le statut d’Edmontosaurus comme saurolophiné ne soit pas contesté, son emplacement exact dans le clade a beaucoup évolué (voir les cladogrammes successifs). Les premières classifications, telle que celle présentée dans l'influente monographie de 1942 de R.S. Lull et Nelda Wright plaçait Edmontosaurus et diverses espèces d’Anatosaurus (dont la plupart seront ultérieurement reclassées dans d'autres espèces ou incorporées dans le genre Edmontosaurus) comme une lignée parmi d'autres lignées d' « hadrosaures à tête plate »[34]. L'une des premières études faites en utilisant des méthodes cladistiques en 1990 a estimé qu'il était lié au genre Anatotitan et Shantungosaurus, dans un clade (alors informel, aujourd'hui une tribu), les Edmontosaurini qui était proche du clade des Saurolophini[34]. Une étude réalisée en 2007 par Terry Gates et Scott Sampson a trouvé des résultats globalement similaires, Edmontosaurus est resté apparenté à Saurolophus et Prosaurolophus et éloigné de Gryposaurus, Brachylophosaurus et Maiasaura[32]. Cependant, la révision de la classification des Hadrosauridae, par Jack Horner et ses collègues (2004), était arrivé à un résultat sensiblement différent : Edmontosaurus serait niché entre Gryposaurus et les brachylophosaures et éloigné de Saurolophus[8]. Les divergences sont compliquées par le manque de travail sur les relations évolutives des hadrosaurinés.
Depuis les années 2010, l'apport de nouvelles données et d'études phylogénétiques à l'échelle des Hadrosauroidea a permis de préciser la taxonomie d'Edmontosaurus. Il est classé comme un Saurolophinae, de la tribu des Edmontosaurini, à laquelle il a donné son nom[33] - [9] - [10].
Découvertes et historique
Claosaurus annectens et autres premières découvertes
Edmontosaurus a une histoire longue et compliquée en paléontologie, ayant passé des décennies avec différentes espèces classées dans d'autres genres. Son histoire taxonomique se confond en divers points avec les genres Agathaumas, Anatosaurus, Anatotitan, Claosaurus, Hadrosaurus, Thespesius et Trachodon[8] - [37] et les articles de référence antérieurs à 1980 utilisent généralement les noms d’Anatosaurus, Claosaurus, Thespesius ou Trachodon pour des fossiles d'edmontosaure (à l'exclusion de ceux affectés à E. regalis), selon les auteurs et les dates. Bien que le nom d’Edmontosaurus ait été créé seulement en 1917, la plus ancienne découverte certaine de son espèce (E. annectens) a été nommée en 1892 et classée comme une espèce du genre Claosaurus et des fragments de fossiles qui peuvent appartenir à ce genre ont été décrits dès 1871[8].
Les premiers vestiges décrits pouvant appartenir au genre Edmontosaurus ont été nommés Trachodon atavus en 1871 par Edward Drinker Cope[38]. Cette espèce a été considérée sans aucune justification comme un synonyme d’Edmontosaurus regalis dans deux revues[8] - [34], bien avant d'être identifié sous le nom d’Atavus regalis pendant plusieurs décennies. En 1874, Cope nomme, sans les décrire, Agathaumas milo une vertèbre sacrée et des fragments de tibias datant de la fin du Maastrichtien (Crétacé supérieur) découverts dans la formation de Laramie au Colorado[6] - [39] - [40]. Plus tard, la même année, il décrit ces os sous le nom d’Hadrosaurus occidentalis[6] - [40] - [41]. Ces os sont aujourd'hui perdus[40]. Comme pour Trachodon atavus, le nom d’Agathaumas milo est attribué sans justification aux restes d’Edmontosaurus regalis dans deux revues[8] - [34] alors qu'ils portaient celui de regalis depuis plusieurs décennies. Aucune de ces deux espèces n'a suscité beaucoup d’intérêt ; c'est ainsi que les deux sont absentes de la monographie de Lulle et Wright en 1942. Une troisième espèce découverte depuis longtemps, Trachodon selwyni, décrite par Lawrence Lambe en 1902 à partir d'une mâchoire inférieure découverte dans ce qui est maintenant connu comme la formation de Dinosaur Park de l'Alberta[42], a été décrite à tort par Glut en 1997 comme ayant été affectée à Edmontosaurus regalis par Lull et Wright[6]. C'était faux et elle aurait dû être classée « de validité douteuse »[43]. C'est ce qui a été confirmé par la suite par plusieurs articles sur les hadrosauridés[8] - [34].
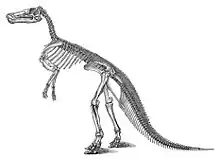

La première espèce qui a été attribuée au genre Edmontosaurus a été appelée en 1892 Claosaurus annectens par Othniel Charles Marsh. Cette espèce a été décrite à partir des restes des squelette USNM 2414 et YPM 2182, ce second étant le paratype. Les deux squelettes ont été découverts en 1891 par John Bell Hatcher dans l'étage de la fin du Maastrichtien (Crétacé supérieur) de la formation de Lance dans le comté de Niobrara (Comté de Converse à l'époque), dans le Wyoming[44]. Cette espèce a quelques particularités historiques : elle est parmi les premiers dinosaures à avoir bénéficié d'une restauration du squelette, et est le premier hadrosauridé ainsi reconstitué[37] - [45] et YPM 2182 et USM 2414 sont, respectivement, les premier et deuxième squelettes de dinosaures pratiquement complets montés aux États-Unis[12] YPM 2182 a été exposé en 1901[37] et USNM 2414 en 1904[12].
En raison de la connaissance incomplète des hadrosauridés à l'époque, après la mort de Marsh, Claosaurus annectens est classé tantôt comme une espèce de Claosaurus, tantôt de Thespesius, tantôt de Trachodon. Les avis varient considérablement suivant les auteurs ; des manuels et encyclopédies faisaient une distinction entre Claosaurus annectens proche des Iguanodon et le genre Hadrosaurus (défini sur les restes maintenant connus sous le nom d’Anatotitan copei), tandis que Hatcher identifiait explicitement C. annectens comme un hadrosauridé[37]. La révision de Hatcher, publiée en 1902, a été lourde de conséquences: il considérait presque tous les genres d'hadrosauridés alors connus comme des synonymes de Trachodon. Cela comprenait les genres Cionodon, Anatotitan, Hadrosaurus, Ornithotarsus, Pteropelyx et Thespesius ainsi que Claorhynchus et Polyonax, genres fragmentaire qu'on pense maintenant appartenir à l'infra-ordre Ceratopsia[46]. Les travaux de Hatcher avaient mené à un bref consensus, jusqu'à ce que, après 1910, de nouveaux fossiles en provenance du Canada et du Montana aient montré qu'il existait une plus grande diversité des hadrosauridés qu'on ne le pensait. [33] Charles W. Gilmore en 1915 a réévalué la famille des hadrosauridés et a recommandé que Thespesius soit réintroduit dans les hadrosauridés de la formation de Lance et les unités rocheuses du même âge, tandis que le genre Trachodon, défini sur des éléments insuffisants, devrait être limité aux hadrosauridés de la formation de Judith River, plus ancienne, et à ses équivalents. En ce qui concerne Claosaurus annectens, Gilmore a recommandé qu'il soit considéré comme un Thespesius occidentalis[47]. La réintégration par Sternberg de Thespesius dans les hadrosauridés[4], dont il était le seul représentant provenant de l'ex-formation de Lance, aujourd'hui la formation de Frenchman[8], aura d'autres conséquences pour la taxonomie des Edmontosaurus dans les décennies suivantes.
Au cours de cette période (1902-1915), deux autres échantillons importants de C. annectens sont récupérés. Le premier, la « momie Trachodon » (AMNH 5060), a été découvert en 1908 par Charles Hazelius Sternberg et ses fils dans les roches de la formation de Lance près de Lusk, dans le Wyoming. Sternberg travaillait pour le Musée d'histoire naturelle de Londres mais Henry Fairfield Osborn du Musée américain d'histoire naturelle a pu acheter l'exemplaire 2 000 $[48]. Les Sternberg ont récupéré un deuxième exemplaire dans la même région en 1910[49], pas aussi bien conservé mais possédant des empreintes de peau. Ils ont vendu ce spécimen (SM 4036) au Muséum Senckenberg en Allemagne[48].
Découvertes canadiennes

La première découverte appelée Edmontosaurus dès l'origine a été faite en 1917 par Lawrence Lambe. Il s'agissait de deux squelettes partiels trouvés dans la formation d'Edmonton le long de la rivière Red Deer dans le sud de l'Alberta au Canada[50]. La formation d'Edmonton a donné son nom au genre[37]. Les fossiles ont été trouvés dans la partie inférieure de la formation maintenant connue sous le nom de formation de Horseshoe Canyon[6] qui est légèrement plus ancienne que les roches dans lesquelles ont été trouvés les restes de Claosaurus annectens[14], renommée Edmontosuarus annectens en 2004[8]. L'espèce type, E. regalis (Edmontosaurus « royal » ou plus modestement « de taille royale »)[37] est basée sur NMC 2288, composé d'un crâne, de vertèbres articulées entre elles jusqu'à la sixième vertèbre caudale, de côtes, d'une partie des os du bassin, d'un humérus (os du bras) et de la plus grande partie d'un membre postérieur. Il a été découvert en 1912 par Levi Sternberg. Le second fossile, le paratype NMC 2289, se compose d'un squelette presque complet où il ne manque que le bec, la plus grande partie de la queue et une partie des pattes a été découvert en 1916 par George F. Sternberg. Lambe a constaté que ce nouveau dinosaure était très proche de Diclonius mirabilis (spécimens maintenant dénommés Anatotitan copei) et a attiré l'attention sur la taille et la robustesse du genre Edmontosaurus[50]. Au départ, Lambe n'a décrit que les crânes des deux squelettes puis il est revenu à l'étude de ce genre en 1920 pour décrire le reste du squelette de NMC 2289[2]. Le reste du squelette de ce spécimen type reste à ce jour non décrit, encore dans ses revêtements en plâtre[6].
Deux espèces supplémentaires qui seront par la suite incorporées dans le genre Edmontosaurus ont été découvertes dans les années 1920, mais elles ont toutes les deux été initialement classées dans le genre Thespesius. Gilmore a nommé la première Thespesius edmontoni en 1924. T. edmontoni a été découvert également dans la formation d'Edmonton. Il est basé sur NMC 8399, un squelette presque complet où il ne manque que la plus grande partie de la queue. NMC 8399 a été découvert près de la rivière Red Deer en 1912 par une équipe de Sternberg[3]. Ses membres antérieurs, ses tendons ossifiés, et des empreintes de peau ont été brièvement décrits en 1913 et 1914 par Lambe, qui avait d'abord pensé qu'il s'agissait d'un exemplaire de Trachodon marginatus[51] avant de se raviser[52]. Ce fossile est devenu le premier squelette de dinosaure à être monté pour être exposé dans un musée canadien. Gilmore a constaté que cette nouvelles espèce était très proche de celle qu'il appelait Thespesius annectens mais a gardé les deux espèces séparées en raison de différences de détail sur les membres antérieurs. Il a également noté que cette espèce avait plus de vertèbres cervicales et dorsales que l'espèce de Marsh mais il a pensé que Marsh s'était trompé en supposant que les spécimens d’annectens étaient complets dans ces régions anatomiques[3].
En 1926, Charles Mortram Sternberg nomma Thespesius saskatchewanensis le fossile NMC 8509, composé d'un crâne et d'un squelette partiel découvert sur le plateau de Wood Mountain, au sud de la Saskatchewan. Il avait recueilli ce spécimen en 1921 dans des roches qui ont été affectées à la formation de Lance[4] maintenant la formation de Frenchman[8]. NMC 8509 comprenait un crâne presque complet, de nombreuses vertèbres, une épaule et une hanche partielles et une partie des membres postérieurs, représentant le premier spécimen important de dinosaure récupéré dans la Saskatchewan. Sternberg choisit de le classer dans le genre Thespesius parce que c'était le seul genre d'hadrosauridés connu dans la formation de Lance à l'époque[4]. À l'époque, T. saskatchewanensis était un Thespesius inhabituel en raison de sa petite taille, estimée entre 7 et 7,3 mètres de longueur[5].
D’Anatosaurus à aujourd'hui
En 1942, Lull et Wright ont essayé de résoudre la taxonomie complexe des hadrosaurinés en créant un nouveau genre, Anatosaurus, regroupant plusieurs espèces qui ne cadraient pas bien dans leurs genres précédents. Le nouveau genre, qui signifie « lézard canard », en raison du large bec rappelant celui des canards, avait comme espèce type l'ancien Claosaurus annectens. En outre ce genre récupérait Thespesius edmontoni, T. saskatchewanensis, une grande mâchoire inférieure que Marsh avait nommée Trachodon longiceps en 1890 et une nouvelle espèce, Anatosaurus copei, dont deux squelettes exposés au Musée américain d'histoire naturelle étaient connus depuis longtemps sous le nom de Diclonius mirabilis (ou ses variantes). Ainsi, ces différentes espèces sont devenues Anatosaurus annectens, A. copei, A. edmontoni, A. longiceps et A. saskatchewanensis[14].

Cet état de choses a perduré pendant plusieurs décennies, jusqu'à ce qu'un étudiant, Michael K. Brett-Surman, réexamine les documents lors de ses études supérieures dans les années 1970 et 1980. Il conclut que l'espèce type d’Anatosaurus, A. annectens, était en fait une espèce d’Edmontosaurus et qu’A. copei était assez différent des autres espèces pour être classé dans son propre genre[36] - [53] - [54]. Bien que les thèses et les mémoires ne soient pas considérés comme des publications officielles par la Commission internationale de nomenclature zoologique, qui réglemente la dénomination des animaux, ses conclusions, connues d'autres paléontologues, ont été adoptées par plusieurs ouvrages de vulgarisation de l'époque[55] - [56]. Brett-Surman et Ralph Chapman ont créé le genre Anatotitan pour classer A. copei en 1990[57]. Parmi les autres espèces, A. saskatchewanensis et A. edmontoni ont été aussi classées dans le genre Edmontosaurus[34] tandis que A. longiceps a été classé dans le genre Anatotitan, soit comme seconde espèce[58] soit comme synonyme d’A. copei[34]. Comme l'espèce type d’Anatosaurus (A. annectens) a été transférée dans le genre Edmontosaurus, le nom Anatosaurus a été abandonné comme synonyme junior d’Edmontosaurus.
Le nouveau genre comprenait trois espèces valides : l'espèce type E. regalis, E. annectens (dont l'ancien Anatosaurus edmontoni qui avait été modifié en edmontonensis) et E. saskatchewanensis[34] Le débat sur la taxonomie appropriée pour les spécimens d’A. copei continue encore aujourd'hui : reprenant les arguments de Hatcher publiés en 1902, Jack Horner, David B. Weishampel et Catherine Forster considèrent ces spécimens comme des Edmontosaurus annectens avec des crânes aplatis[8]. En 2007, la description d'une autre "momie", « Dakota », a été publiée; elle a été découverte en 1999 par Tyler Lyson dans la formation de Hell Creek au Dakota du Nord[24] - [25].
Inventaire des fossiles retrouvés
En 1908, Charles Hazelius Sternberg, un disciple de Cope, découvrit avec ses fils (Lévi, Charles et George) une empreinte de peau de l'Anatosaurus. Ce fut la première empreinte jamais découverte.
En 1999, un spécimen, nommé Dakota (en), âgé entre -67 et -65 millions d'années, dans un état de conservation idéal puisque momifié avant de s'être pétrifié, a été retrouvé dans la formation de Hell Creek (dans l'est du Montana et l'ouest des Dakota) par Tyler Lyson (en), un lycéen américain passionné de paléontologie. Il a fini par être déterré totalement et étudié en 2006. Grâce à ce fossile, on a pu démontrer que l'edmontosaure mesurait jusqu'à un mètre de plus que ce que l'on pensait avant (l'écart entre chaque vertèbre par rapport à l'autre serait de 1 centimètre, et ces dernières ne seraient pas accolées les unes aux autres, comme l'on pensait) lui conférant une plus grande agilité. Sa partie postérieure aurait été 25 pour cent plus massive qu'estimé lors des reconstitutions antérieures, sa peau était constituée de cuir, et il pouvait courir jusqu'à plus de 50 km/h. La peau était si bien conservée que l'on y a retrouvé des traces sur certaines parties du corps de l'animal suggérant qu'il possédait des bandes et des rayures tel le zèbre[59] - [60].
Le Muséum national d'histoire naturelle, à Paris, possède un squelette d'edmontosaure découvert en 1910 aux États-Unis et envoyé en France en 1911[61]. Ce spécimen est resté dans les réserves du Muséum de 1911 à 2018, année où il a été décidé de monter le squelette sous la direction du paléontologue Ronan Allain et de la lithopréparatrice Hélène Bourget. Le dévoilement au public de cette pièce paléontologique de grande valeur est la conséquence d'une exposition itinérante organisée par le musée Naturalis des Pays-Bas, qui possède un squelette authentique de tyrannosaure, le spécimen « Trix », ainsi surnommé en honneur de la reine néerlandaise Beatrix. La vedette de cette exposition (ayant déjà parcouru les Pays-Bas, l'Autriche et l'Espagne avant d'arriver à Paris en juin 2018) était tout naturellement le tyrannosaure « Trix », mais le Muséum français entendit que du point de vue de la pédagogie il était utile de montrer aussi au public son propre squelette d'edmontosaure puisqu'effectivement les edmontosaures comptaient parmi les proies les plus habituelles des tyrannosaures[62].
Espèces et répartition
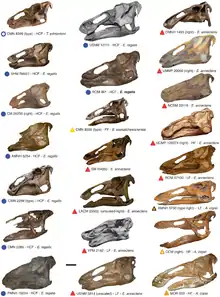
On considère actuellement qu’Edmontosaurus possède au moins trois espèces valides : l'espèce type E. regalis, E. annectens et E. saskatchewanensis[8] - [34].
E. regalis a été retrouvé dans la formation de Horseshoe Canyon, les formations de la rivière Sainte Marie et de Scollard dans l'Alberta, la formation de Hell Creek au Montana, au Dakota du Nord et au Dakota du Sud, la formation de Lance au Dakota du Sud et au Wyoming et la formation de Laramie au Colorado, qui datent toutes du Maastrichtien, à la fin du Crétacé supérieur. Au moins une douzaine d'individus ont été retrouvés, dont sept crânes avec des restes de squelette associés et cinq à sept crânes seuls, Trachodon atavus et Agathaumas milo étant des synonymes possibles[8] - [34].
E. annectens a été retrouvé dans la formation de Scollard dans l'Alberta, la formation de Hell Creek au Montana et au Dakota du Sud, la formation Lance au Dakota du Sud et au Wyoming, et la formation de Laramie au Colorado. Il n'aurait vécu qu'à la fin du Maastrichtien et est connue par au moins cinq crânes avec restes de squelette[8]. Un chercheur, Kraig Derstler a écrit qu’E. annectens était « peut-être le dinosaure le mieux connu à ce jour » (1994)[63]. Selon une étude de 2004, Thespesius edmontoni (ou edmontonensis), Anatosaurus copei et Trachodon longiceps devraient être considérés comme de possibles synonymes d’E. annectens[8]. On admettait généralement que T. edmontoni pouvait être assimilé à E. annectens depuis 1990[34] mais les cas d’A . copei et T. longiceps n'ont pas encore été validés par d'autres auteurs. S'ils s'avèrent appartenir à E. annectens, le total augmenterait de cinq individus affectés précédemment à A. copei[57] et de la mâchoire inférieure qui sert d'holotype à T. longiceps (le nombre donné pour E. annectens par J. Horner en 2004 est incorrect car il n'est pas modifié par rapport à une publication antérieure[34] qui ne considère pas A. copei et T. longiceps comme synonymes d’E. annectens). La liste des formations pourrait être aussi erronée, pour la formation dont vient T. edmontoni (il n'est pas précisé de quelle partie de l'ancienne formation Edmonton il provient). T. edmontoni pourrait avoir été mal affecté ; James Hopson, en 1975, pensait que cela pouvait être un jeune E. regalis[16] et Nicolas Campione a également constaté que son crâne ne pouvait être différencié d’E. regalis contemporain dans une pré-étude de 2009[64]. E. annectens diffère d’E. regalis par son crâne plus long, plus aplati, moins robuste[6]. Mais Brett-Surman considérait E. regalis et E. annectens comme les possibles représentants mâles et femelles de la même espèce[36]. Tous les spécimens d’E. regalis proviennent toutefois de formations plus anciennes que ceux d’E. annectens[64].
E. saskatchewanensis est connu à partir de quatre ou cinq individus, la plupart n'étant représentés que par des crânes. Il n'a été retrouvé que dans la formation de Frenchman de Saskatchewan datant de la fin du Maastrichtien[8]. Il se distingue des deux autres espèces par sa taille plus petite[4] - [6]. Il n'a été publié que peu de choses sur cette espèce depuis sa description initiale, en comparaison avec les deux autres espèces. Les travaux préliminaires de Nicolas Campione ont montré qu'il s'agissait probablement d'un synonyme d’E. annectens mais une étude plus approfondie est nécessaire pour le confirmer ou l'infirmer[64].
En outre, il existe de nombreux fossiles d’Edmontosaurus qui n'ont pas été identifiés par espèce. Les restes qui n'ont pas été attribués à une espèce particulière (identifiés comme E. sp.) peuvent étendre le territoire occupé autrefois par le genre jusqu'à la formation Prince Creek en Alaska[65] et la formation de Javelina au Texas[66].
Paléoécologie
Le genre Edmontosaurus est un genre de vaste répartition temporelle et spatiale. Les gisements où on l'a trouvé peuvent être divisés en deux groupes selon leur âge : les formations de Horseshoe Canyon et de la rivière Ste Marie d'une part, et les formations de Frenchman, Hell Creek, Lance, Laramie et Scollard de l'autre. La première période, la plus ancienne, est également connue sous le nom d'« Edmontonien » alors que la seconde, correspondant à des gisements plus jeunes est connue sous le nom de « Lancien ». Ces deux périodes informelles avaient des faunes de dinosaures différentes[67].
La paléoécologie de l'Edmontonien
Le début de l'Edmontonien est défini par la première apparition d’Edmontosaurus regalis dans les gisements[68]. Bien qu'on ne la considère parfois comme ne datant que du début du Maastrichtien[66], la formation du canyon Horseshoe couvre une période un peu plus longue. Les premiers dépôts datant d'environ 73 millions d'années, donc de la fin du Campanien et les derniers d'entre il y a 68,0 à 67,6 millions d'années[69]. Edmontosaurus regalis a été retrouvé dans la plus ancienne des cinq couches de la formation de Horseshoe Canyon mais il est absent des autres[70]. Près des trois quarts des spécimens de dinosaures trouvés dans les badlands près de Drumheller, dans l'Alberta, semblent appartenir au genre Edmontosaurus[71]. La formation de Horseshoe Canyon semble avoir connu une forte influence marine, en raison d'un empiétement de la Voie maritime intérieure de l'Ouest, la mer peu profonde qui a recouvert la partie centrale de l'Amérique du Nord pendant la plus grande partie du Crétacé[67]. E. regalis partageait les lieux avec ses compatriotes hadrosauridés Hypacrosaurus et Saurolophus, l'hypsilophodonte Parksosaurus, les dinosaures à cornes Montanoceratops, Anchiceratops, Arrhinoceratops et Pachyrhinosaurus, le pachycéphalosauridé Stegoceras, l'ankylosauridé Euoplocephalus, le nodosauridé Edmontonia, les ornithomimosauridés Ornithomimus et Struthiomimus, divers petits théropodes mal connus, comme des troodontidés et des dromæosauridés et les tyrannosauridés Albertosaurus et Daspletosaurus[66]. Edmontosaurus vivait dans les régions côtières tandis que Hypacrosaurus et Saurolophus vivaient plus à l'intérieur des terres[72]. Edmontosaurus et Saurolophus vivaient pas habituellement ensemble[72]. L'habitat typique des edmontosaures de cette formation a été décrit comme les régions à proximité des marécages de cyprès chauves et des tourbières le long des deltas côtiers. Pachyrhinosaurus préférait lui aussi cet habitat aux plaines inondables où vivaient Hypacrosaurus, Saurolophus, Anchiceratops et Arrhinoceratops[71]. L'association Pachyrhinosaurus-Edmontosaurus dans les régions côtières de l'Edmontonien est retrouvée jusqu'en Alaska[73].
La paléoécologie du Lancien
Le Lancien est la dernière période avant l'extinction ses dinosaures non-aviens à la fin du Crétacé. Edmontosaurus est l'un des dinosaures les plus communs de cette période. Robert Bakker rapporte qu'il représente jusqu'à un septième du nombre de spécimens de grands dinosaures de cette époque dans la région ; le plus grand nombre (les cinq sixièmes) étant composé de Triceratops[74]. Les plaines où vivaient l'association Triceratops-Edmontosaurus, dominée par le premier, s'étendaient du Colorado à la Saskatchewan[73]. La faune typique de dinosaures des formations où vivaient Edmontosaurus au Lancien comprenaient également l'hypsilophodonte Thescelosaurus, quelques rares hadrosauridés Anatotitan, quelques rares cératopsidés Nedoceratops (= "Diceratops") et Torosaurus, les pachycephalosauridés Pachycephalosaurus et Stygimoloch, les ankylosauridés Ankylosaurus, les théropodes Ornithomimus, Troodon et Tyrannosaurus[66] - [75].

La formation de Hell Creek, telle qu'on peut l'interpréter par l'affleurement de la région de Fort Peck au Montana, est considérée comme une ancienne plaine inondable couverte de forêts, qui avait un climat subtropical relativement sec et qui possédait une grande variété de végétaux, comme des angiospermes arborescents, des conifères comme le cyprès chauve, des fougères et ginkgos. Le littoral était à des centaines de kilomètres à l'est. On y trouvait des tortues d'eau et divers mammifères multituberculés arboricoles et des varans aussi grands que l'actuel dragon de Komodo y chassaient. Le Triceratops était le plus abondant des grands dinosaures et le Thescelosaurus le plus abondant des petits dinosaures herbivores. Les restes d'edmontosaures ont été rassemblés ici dans les sables des canaux, dont des fossiles de jeunes individus d'un mètre de long. Les accumulations de fossiles d'edmontosaures sont probablement dues à des groupes en mouvement[76].
La formation de Lance qui affleure à environ 100 km au nord de Fort Laramie dans l'est du Wyoming, est considérée comme un dépôt de bayou semblable à ceux des plaines côtières de la Louisiane. Il était plus proche d'un grand delta que la formation de Hell Creek située plus au nord et a reçu beaucoup plus de sédiments. Des Araucarias et des palmiers, des plantes tropicales, poussaient dans les forêts de feuillus, une végétation différente de la flore de la plaine côtière plus au nord[77]. Le climat était humide et subtropical, avec des conifères, des palmiers nains et des fougères dans les marais, des conifères, des frênes, des Cyclobalanopsis et des arbustes dans les forêts[63]. Des poissons, des salamandres, des tortues, des lézards, des serpents, des oiseaux et de petits mammifères vivaient aux côtés des dinosaures. Les petits dinosaures ne sont pas trouvés ici en aussi grande quantité que dans les sédiments de Hell Creek mais Thescelosaurus semble une fois de plus y avoir été relativement abondant. Triceratops y est connu par de nombreux crânes, qui ont tendance à être un peu plus petits que ceux des individus trouvés plus au Nord. La formation de Lance a fourni deux « momies » d'edmontosaures[77].
Paléobiologie
Cerveau et système nerveux
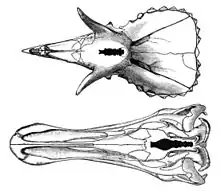
Le cerveau d’Edmontosaurus a été décrit dans plusieurs articles à partir d'études utilisant des moulages de la cavité crânienne. On a étudié ainsi les cerveaux d’E. annectens[78] - [79] et d’E. regalis[2] ainsi que de spécimens dont l'espèce n'est pas encore connue[80] - [81] - [82]. Le cerveau n'était pas particulièrement important pour un animal de cette taille. La longueur disponible pour le cerveau n'étant d'environ que le quart de la longueur du crâne[2] et divers moulages ont trouvé un volume endocrânien de 374 à 450 cm3[81]. Or il faut tenir compte du fait que le cerveau pouvait n'avoir occupé que 50 % de cet espace, le reste étant occupé par la dure-mère qui entoure le cerveau[81] - [82]. Par exemple, le cerveau du spécimen dont l'endocrâne mesurait 374 cm3 est estimé n'avoir eu qu'un volume de 268 cm3[82]. Le cerveau avait une structure allongée[81] et comme chez les animaux autres que les mammifères, il n'avait aucun néocortex[82]. Comme chez Stegosaurus, le tube neural était élargi au niveau des hanches, mais pas au même degré : l'espace endosacral du Stegosaurus était 20 fois le volume de son endocrâne tandis que l'espace endosacral de l’Edmontosaurus était seulement 2,59 fois plus important[81].
Adaptations alimentaires
Comme tous les hadrosauridés, Edmontosaurus était un grand herbivore terrestre. Ses dents poussaient constamment et étaient remplacées par des séries de dents nouvelles se développant en permanence, seulement une poignée d'entre elles étant en usage à un moment donné[8]. Il utilisait son large bec pour couper les végétaux tout venants, peut-être en broutant[8], peut-être en fermant les mâchoires sur les rameaux et les branches pour, en tirant la tête récupérer les feuilles et les pousses les plus nutritives[19]. Comme les dents étaient implantées à l'extérieur de la mâchoire, et en s'appuyant sur d'autres détails anatomiques, on en a déduit qu’Edmontosaurus et la plupart des autres ornithischiens avaient des structures analogues à nos joues, dotées de muscles ou non, dont le rôle était de conserver la nourriture dans la bouche[83] - [84]. La zone d'alimentation de l'animal allait du niveau du sol à environ 4 mètres de hauteur[8].
Avant les années 1960 et 1970, beaucoup pensaient que les hadrosauridés comme Edmontosaurus vivaient dans l'eau et se nourrissaient de plantes aquatiques[85]. Un exemple de ce type d'interprétation est celui de William Morris dans les années concevant l'animal avec un bec non ossifié. Il imaginait alors que l'animal avait un régime alimentaire semblable à celui de certains canards modernes, avec un système de filtration des mollusques et crustacés aquatiques et en rejetant l'eau par des sillons en forme de V le long de la face interne de la partie supérieure du bec[13]. Cette interprétation du bec a été rejetée, car les sillons et les crêtes des becs d'edmontosaures ressemblent davantage à celles des becs de tortues herbivores qu'aux structures flexibles observées chez les oiseaux qui se nourrissent par filtration[85].

Le principal modèle actuel sur la façon dont les hadrosauridés se nourrissaient a été avancé en 1984 par le paléontologue David B. Weishampel. Il a proposé que la structure du crâne permettait des mouvements d'avant en arrière de la mâchoire inférieure, et un possible écartement des os de la mâchoire supérieure portant les dents lorsque la bouche était fermée. Les dents de la mâchoire supérieure broyaient les végétaux contre les dents de la mâchoire inférieure comme des râpes[8] - [86]. Un tel type de mouvement est analogue à celui de la mastication chez les mammifères, bien que s'accomplissant dans un environnement complètement différent[87]. Un élément de preuve important en faveur du modèle imaginé par Weishampel est l'orientation des rayures sur les dents, traduisant des mouvements d'avant en arrière de la mâchoire. D'autres mouvements auraient pu produire des rayures similaires, comme des mouvements entre eux des os des deux moitiés de la mâchoire inférieure. Tous les mouvements possibles n'ont pas été examinés au moyen des techniques modernes actuelles[88].
Weishampel a développé son modèle à l'aide d'une simulation par ordinateur. Natalia Rybczynski et ses collègues ont mis à jour ce travail avec un modèle d'animation en trois dimensions beaucoup plus sophistiqué, la numérisation d'un crâne d’E. regalis à l'aide de lasers. Ils ont pu reproduire les mouvements proposés par Weishampel avec leur système d'études, mais ils ont pu constater que des mouvements secondaires entre d'autres os étaient nécessaires, avec des séparations au maximum de 1,3 à 1,4 centimètre entre certains os au cours du cycle de la mastication. Rybczynski et ses collègues n'étaient pas convaincus que le modèle Weishampel soit viable, mais ont trouvé plusieurs améliorations à apporter à leur animation. Les améliorations prévues comprennent l'incorporation de tissus mous et des marques d'usure et de rayures des dents, ce qui devrait mieux limiter les mouvements. Ils notent qu'il existe plusieurs autres hypothèses à tester de la même manière[87]. De nouveaux travaux de Casey Holliday et Lawrence Witmer ont constaté que les ornithopodes comme Edmontosaurus ne possédaient pas les types d'articulation des os du crâne rencontrés chez les animaux modernes connus pour avoir des crânes dits cinétiques (crânes permettant des mouvements entre les os le constituant), tels que les squamates et les oiseaux. Ils pensent que ce qui avait été interprété comme des articulations permettant des mouvements des os du crâne des dinosaures était en fait des zones de croissance cartilagineuse des os[88].
La conception d'un crâne aux os soudés entre eux a été contestée en 2009 par Vincent Williams et ses collègues. Se référant aux microtraces retrouvées sur les dents, ils ont trouvé quatre types de rayures différents. La type le plus courant a été interprété comme résultant d'un mouvement oblique et non d'un simple mouvement de haut en bas ou avant-arrière, ce qui est plus cohérent avec le modèle de Weishampel. On pense que ce mouvement était le principal mouvement pour le broyage des aliments. Deux autres types de rayures ont été interprétés comme résultant de mouvements vers l'avant ou vers l'arrière de la mâchoire. Le dernier type est variable et résulte probablement de l'ouverture des mâchoires. La combinaison de ces mouvements est plus complexe qu'on ne l'avait prédit. Parce que les rayures l'emportent sur les microtraces, Williams et collaborateurs ont suggéré qu’Edmontosaurus se nourrissait en hauteur (grazer) et non au niveau du sol (browser), ce qu'on peut savoir par le faible nombre d'éraflures dues à la consommation de matières abrasives que sont les plantes riches en silice comme les prêles et la terre qui est accidentellement ingérée en broutant au niveau du sol[89].
La description de la présence de gastrolithes dans l'estomac d'un Claosaurus semble basée sur une probable double erreur. Tout d'abord, ce spécimen est maintenant considéré comme un Edmontosaurus annectens. Barnum Brown, qui a découvert le spécimen en 1900, l'avait classé comme un Claosaurus parce qu’E. annectens était considéré comme une espèce de Claosaurus à l'époque. En outre, il est probable que les gastrolithes soient en fait des graviers entraînés là pendant l'enfouissement du cadavre[37].
Contenu digestif
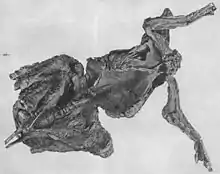
Les deux « momies » recueillies par l'équipe de Sternberg avaient peut-être encore le contenu de leur tube digestif. Charles H. Sternberg a signalé la présence d'un tel contenu carbonisé dans le spécimen du Musée américain d'histoire naturelle[90] mais ce matériel n'a jamais été décrit[91]. Les végétaux découverts dans le specimen du Musée Senckenberg ont été décrits, mais leur description s'est avérée difficile à interpréter. Les plantes trouvées comprennent des aiguilles du conifère Cunninghamites elegans, des brindilles de conifères et de feuillus et de nombreuses petites graines et fruits[92]. Leur description en 1922 a fait l'objet d'un débat dans le journal de langue allemande Paläontologische Zeitschrift. Kräusel, qui avait décrit les matériaux, les a considérés comme le contenu de l'intestin de l'animal[92] tandis qu'Abel ne pouvait pas exclure que les plantes n'aient été entraînées dans la carcasse après la mort de l'animal[93].

À l'époque, les hadrosauridés étaient considérés comme des animaux aquatiques, et Kräusel affirme que les résultats trouvés pour ce spécimen n'excluent pas que les hadrosauridés mangeaient des plantes d'eau[19] - [92]. Ces découvertes n'ont eu que peu d'impact sur les auteurs de langue anglaise, en dehors peut-être d'une brève mention de la possible double vie aquatique-terrestre de ces animaux[94] et ce jusqu'à ce que John Ostrom remette en cause dans un article l'ancienne conception des hadrosauridés comme animaux aquatiques. Au lieu d'essayer d'adapter la découverte faite au modèle aquatique, il a utilisé cette dernière comme un élément de preuve que les hadrosauridés étaient des herbivores terrestres[19]. Même si sa conception des hadrosauridés comme des animaux terrestres a été généralement acceptée[8], les plantes fossiles de Senckenberg laissent des doutes. Kenneth Carpenter a laissé entendre qu'elles pouvaient en fait représenter le contenu des intestins d'un animal affamé et non un régime alimentaire typique[27] - [28]. D'autres auteurs ont noté que, parce que les fossiles de plantes ont été retirés de leur contexte d'origine dans l'échantillon et ont été fortement préparés, il n'est plus possible d'assurer le suivi sur le travail original, en laissant ouverte la possibilité que les plantes aient été emmenées avec les débris[91] - [95].
Les études isotopiques
L'alimentation et la physiologie d’Edmontosaurus ont été étudiées au moyen d'isotopes stables de carbone et d'oxygène conservé dans l'émail des dents. Lorsqu'ils mangent, boivent ou respirent, les animaux absorbent du carbone et de l'oxygène, qui sont incorporés dans les os. Les proportions des isotopes de ces deux éléments sont déterminées par divers facteurs internes et externes, tels que le type de plantes consommé, la physiologie de l'animal, la salinité du milieu et le climat. Si les rapports isotopiques des fossiles ne sont pas altérés par la fossilisation et les changements survenus plus tard, ils peuvent être étudiés pour obtenir des renseignements sur les conditions de vie à l'origine; les animaux à sang chaud auront des compositions isotopiques différentes de celles de leur environnement, les animaux qui se nourrissent de certains types de plantes ou utilisent certains procédés digestifs auront des compositions isotopiques distinctes, et ainsi de suite. On utilise en général l'émail des dents pour ces études car la structure du minéral qui le compose est la plus résistante aux changements chimiques du squelette[18].
Une étude réalisée en 2004 par Kathryn Thomas et Sandra Carlson a utilisé les dents de la mâchoire supérieure de trois individus considérés comme un jeune, un subadulte et un adulte, provenant de la formation de Hell Creek dans le comté de Corson, au Dakota du Sud. Dans cette étude, on a échantillonné différentes dents d'une colonne dentaire à l'aide de multiples microperçages. Cette méthode a l'avantage de déterminer les variations des isotopes contenus dans les dents sur des périodes de temps courtes. De ce travail, il semble qu'on puisse conclure que les dents d'edmontosaures demandaient environ huit mois pour se former, la formation étant un peu plus rapide chez les jeunes edmontosaures. Les dents de ces trois individus semblent montrer des variations de ratios isotopiques de l'oxygène qui pourraient correspondre à des changements de climat chaud et sec ou frais et humide; Thomas et Carlson ont envisagé la possibilité pour ces animaux d'avoir migré mais ils ont préféré favoriser l'hypothèse de variations saisonnières locales car la migration aurait plus probablement conduit à une homogénéisation des ratios, beaucoup d'animaux migrant pour rester dans des plages spécifiques de température ou à proximité de sources de nourriture particulière[18].
On a également trouvé chez ces edmontosaures des concentrations isotopiques supérieures à celles de l'environnement pour le carbone, ce qui, chez les mammifères modernes serait interprété comme des animaux ayant un régime mixte à base de plantes en C3 (la plupart des plantes) et de plantes en C4 (graminées), mais les plantes en C4 étaient extrêmement rares au Crétacé supérieur peut-être même complètement absentes. Thomas et Carlson ont avancé plusieurs hypothèses pour expliquer cette situation, les plus probables incluant un régime riche en gymnospermes, une consommation élevée en sel -les plantes poussant dans des zones côtières adjacentes à la voie maritime intérieure de l'Ouest- et une différence physiologique entre dinosaures et mammifères pour former les tissus. Une combinaison de ces facteurs est également possible[18].
Pathologies et santé
En 2003, on a apporté la preuve de l'existence de tumeurs, comme des hémangiomes, des fibromes desmoplastiques, des cancers métastasés et des ostéoblastomes sur des os d’Edmontosaurus. Rothschild et les a mises en évidence sur des vertèbres de dinosaures en utilisant la tomographie par ordinateur et la fluoroscopie. Plusieurs autres hadrosauridés, comme Brachylophosaurus, Gilmoreosaurus et Bactrosaurus se sont révélés également porteurs de telles maladies. Alors que ces études ont porté sur plus de 10 000 fossiles, ces tumeurs n'ont été retrouvées que chez Edmontosaurus et des genres très proches. On ne sait pas si ces tumeurs ont été causées par des facteurs environnementaux ou une prédisposition génétique[96].
On a retrouvé aussi des cas d'ostéochondrose chez Edmontosaurus. Cette pathologie, qui résulte du remplacement anormal de cartilages articulaires par du tissu osseux durant la croissance, a été retrouvée sur 2,2 % des 224 os des orteils. La cause de cette situation est inconnue. On a proposé plusieurs origines possibles : une prédisposition génétique, des traumatismes, le régime alimentaire, des troubles circulatoires, un excès d'hormones thyroïdiennes, des carences en facteurs de croissance...
Chez les dinosaures, l'ostéochondrose (comme les tumeurs) ont été le plus souvent trouvées chez les hadrosauridés[97].
Locomotion
Comme pour d'autres hadrosauridés, on pense qu’Edmontosaurus était un bipède facultatif qui, s'il passait la plupart du temps à quatre pattes, pouvait se dresser sur ses pattes arrière en cas de besoin. Il est probable qu'il restait à quatre pattes lorsqu'il était immobile ou se déplaçait lentement et se mettait sur ses pattes postérieures seulement pour se déplacer plus rapidement[8]. Les recherches menées par modélisation informatique en 2007 suggèrent qu’Edmontosaurus pouvait courir vite, peut-être jusqu'à 45 km/h[24]. D'autres simulations faites en utilisant un spécimen subadulte dont le poids est estimé à 715 kg de son vivant a produit un modèle qui pouvait marcher, courir à quatre pattes ou sur ses pattes arrière, sauter à pieds joints, trotter ou galoper. Les chercheurs ont découvert à leur grande surprise que la démarche la plus rapide était le saut à pieds joints comme chez les kangourous avec une vitesse maximale de 62 km/h ce qui semble peu probable en raison de la taille de l'animal et du manque de traces fossiles retrouvées. Ce résultat a donc été interprété comme le signe d'une inexactitude dans la simulation. Les plus rapides des allures en dehors du saut étaient le galop (57 km/h) et la course sur ses pattes postérieures (maximum de vitesse simulée de 50 km/h). Ils ont eu une faible préférence pour la course en position bipède comme option la plus probable pour ses déplacements à grande vitesse, mais ils n'ont pas exclu un déplacement quadrupède rapide[98].
On a longtemps pensé qu'ils étaient des animaux aquatiques ou semi-aquatiques, mais les hadrosauridés n'étaient pas mieux adaptés à l'eau que d'autres dinosaures (en particulier les théropodes qu'on considérait autrefois comme incapables de poursuivre les hadrosauridés dans l'eau). Les hadrosauridés avaient des mains fines aux doigts courts, ce qui rendait leurs membres antérieurs peu efficaces pour se propulser dans l'eau et la queue n'était pas beaucoup plus utile en raison des tendons ossifiés qui augmentaient sa rigidité et du peu de développement des points d'attache des muscles qui auraient été nécessaires pour remuer la queue d'un côté à l'autre[29] - [99].
Relations avec les théropodes

Edmontosaurus vivait à la même époque et dans les mêmes lieux que Tyrannosaurus et un spécimen adulte d’ E. annectens exposé au Musée de la Nature et des Sciences de Denver présente des signes d'une morsure de théropode au niveau de la queue. Les apophyses épineuses des treizième à la dix-septième vertèbres caudales sont endommagées, lésions compatibles avec une attaque par l'arrière droit de l'animal. Une apophyse a été en partie cisaillée et les autres sont déformées; trois d'entre elles portent des marques apparentes de morsure. Le dessus de la queue était au moins à 2,90 mètres de hauteur et la seule espèce connue de théropode de ce niveau stratigraphique assez grand pour porter une telle attaque est T. rex. Les os ont partiellement guéri, mais l'animal est décédé avant que les traces de l'attaque n'aient complètement disparu. Les os montrent également des signes d'infection. Kenneth Carpenter, qui a étudié le squelette, a noté qu'il semble également avoir eu une fracture guérie au niveau de la hanche gauche avant cette attaque, car elle est mieux guérie. Il a suggéré que cet edmontosaure était une proie facile car il aurait pu boiter après la première blessure. Comme il a survécu à l'attaque, Carpenter a suggéré qu'il aurait pu avoir manœuvré ou couru plus vite que son attaquant, ou que les dommages à sa queue sont dues au fait qu'il avait utilisé sa queue comme arme contre le tyrannosaure[100].
Un autre spécimen d’E. annectens mesurant 7,60 mètres de long et trouvé dans le Dakota du Sud, montre des signes de morsures par de petits théropodes au niveau de sa mâchoire inférieure. Certaines morsures ont partiellement guéries. Michael Triebold, discutant de manière informelle de ce spécimen, a proposé un scénario où de petits théropodes attaquaient l'edmontosaure à la gorge ; l'animal aurait survécu à l'attaque initiale, mais succombé à ses blessures peu de temps après[101]. Certains gisements d'edmontosaures correspondent à des charniers. Des marques de morsure par Albertosaurus et Saurornitholestes sont communes dans un site de l'Alberta[102]. Daspletosaurus se nourrissait également d’Edmontosaurus et de Saurolophus, un autre hadrosauridé, dans un autre site de l'Alberta[103].
Comportement social
On connait de vastes gisements d'edmontosaures et la taille de ces gisements suggère que ces animaux étaient grégaires[8].Quatre sites contenant des restes d'edmontosaures sont identifiés dans un répertoire sur les gisements de fossiles : en Alaska (formation de Prince Creek), Alberta (formation de Horseshoe Canyon), Dakota du Sud (formation de Hell Creek) et Wyoming (formation de Lance)[65]. Un lit d'edmontosaures de la formation de Lance dans l'est du Wyoming, couvre plus d'un kilomètre carré, les os d’Edmontosaurus sont plus abondants sur 40 hectares de ce site. On estime entre 10 000 et 25 000 le nombre d'edmontosaures présents là[104].
Contrairement à beaucoup d'autres hadrosauridés, Edmontosaurus n'avait pas de crête osseuse. Il pouvait avoir cependant des structures molles bien apparentes au niveau de la tête : l'os entourant les ouvertures nasales avait de profondes encoches et ces cavités semblent avoir servi de sacs gonflables, servant peut-être à la fois de signal visuel et auditif[16]. Edmontosaurus pourrait avoir présenté un dimorphisme sexuel, car on a retrouvé des formes osseuses plus ou moins robustes mais on n'a pas établi que cela soit lié à un dimorphisme sexuel[105].
En raison de sa large répartition, de l'Alaska au Colorado, inclut des zones polaires qui avaient peu de lumière durant une grande partie de l'année, Edmontosaurus est considéré comme ayant pu être un animal migrateur. Une étude sur la migration des dinosaures faite par Phil R. Bell et Eric Snively envisage qu’E. regalis était capable de parcourir 2 600 km aller-retour par an à condition d'avoir des quantités de nourriture et de graisse suffisantes. Un tel voyage imposait une vitesse moyenne de marche d'environ 2 à 10 kilomètres à l'heure et aurait pu se faire entre l'Alaska à l'Alberta. Cette possible migration des edmontosaures contraste avec de nombreux autres dinosaures, comme les théropodes, les sauropodes, et les ankylosaures, que Bell et Snively pensent susceptibles d'avoir été capables d'hiverner[106] - [107].
Dans la culture populaire
Documentaires, Films et Séries
- Dans le documentaire "La marche des dinosaures", qui raconte le déroulement de la première migration d'un edmontosaure.
- Dans le documentaire de la BBC Planète Dinosaure, on y voit un troupeau qui se fait attaquer par un groupe de Troodon, un juvénile séparé des autres edmontosaures se fait assaillir par les prédateurs, la mère arrive à son secours mais le lendemain celui-ci meurt.
- Dans le documentaire Jurassic Fight Club, un edmontosaure se fait attaquer par des Dromaeosaurus.
- Dans le documentaire Planète préhistorique, une femelle edmontosaure sauve ces enfants des flammes
Jeux vidéos
- Edmontosaurus est présent dans le jeu de gestion Jurassic World Evolution (2018).
- Dans l'univers Dinosaur King, il apparait seulement dans les jeux vidéos
Galerie
- Edmontosaurus annectens, National Museum of Natural History, Washington D.C.
- "Momie" d'Edmontosaurus, découverte en 1912. Cette dernière est bien moins complète que celle découverte en 1999. Edmontosaurus est l'un des rares dinosaures dont on a trouvé de nombreux spécimens très bien conservés et momifiés. Des empreintes de peau ont notamment été trouvées.
- Peau fossile d'Edmontosaurus. Cette partie provient de l'abdomen. On y distingue des milliers de petites "écailles" rondes et des plis. Edmontosaurus est l'un des rares dinosaures dont on connait la peau. Des découvertes récentes tendent à prouver qu'Edmontosaurus était zébré.
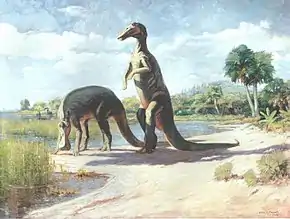 Reconstitution sans doute erronée, datant de 1905. L'espèce de Trachodon après avoir été renommée Anatotitan est aujourd'hui attribuée à Edmontosaurus annectens. Notons qu'ici l'animal traîne sa queue au sol, ce que l'on a prouvé aujourd'hui comme impossible chez les dinosaures.
Reconstitution sans doute erronée, datant de 1905. L'espèce de Trachodon après avoir été renommée Anatotitan est aujourd'hui attribuée à Edmontosaurus annectens. Notons qu'ici l'animal traîne sa queue au sol, ce que l'on a prouvé aujourd'hui comme impossible chez les dinosaures. Représentation ancienne de Trachodon.
Représentation ancienne de Trachodon.
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
Articles
- (en) Lawrence M. Lambe, « A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta », The Ottawa Naturalist, vol. 31, no 7, , p. 65–73 (lire en ligne [PDF] (entire volume, 18 mb), consulté le ).

- (en) B.M. Rothschild et Tanke, D. H.; Helbling II, M.; and Martin, L.D., « Epidemiologic study of tumors in dinosaurs », Naturwissenschaften, vol. 90, no 11, , p. 495–500 (PMID 14610645, DOI 10.1007/s00114-003-0473-9, lire en ligne, consulté le ).
Ouvrages
- (en) Lawrence M. Lambe, The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta, 120, Department of Mines, Geological Survey of Canada, (ISBN 0-659-96553-4, lire en ligne [PDF]), p. 1–79.
- (en) Charles W. Gilmore, A new species of hadrosaurian dinosaur from the Edmonton Formation (Cretaceous) of Alberta, 38, Department of Mines, Geological Survey of Canada, , p. 13–26.
- (en) Charles M. Sternberg, A new species of Thespesius from the Lance Formation of Saskatchewan, 44, Department of Mines, Geological Survey of Canada, , p. 77–84.
- (en) Richard Swann Lull et Nelda E. Wright, Hadrosaurian Dinosaurs of North America, vol. 40, GSA : Geological Society of America, (ISBN 9780813720401, DOI 10.1130/SPE40), p. 50–93.
- (en) Donald F. Glut, Dinosaurs : The Encyclopedia, Jefferson, North Carolina, McFarland & Co, , 1076 p., relié (ISBN 978-0-89950-917-4, LCCN 95047668), « Edmontosaurus », p. 389–396.
- (en) David Lambert et the Diagram Group, The Dinosaur Data Book, New York, Avon Books, (ISBN 978-0-380-75896-8), p. 60
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :
Références taxinomiques
- (en) Référence Paleobiology Database : Edmontosaurus Lambe, 1917
- (en) Référence Paleobiology Database : Edmontosaurus regalis Lambe, 1917
Notes et références
Références
- Lambe 1917, p. 65–73.
- Lambe 1920, p. 1–79.
- Gilmore 1924, p. 13–26.
- Sternberg 1926, p. 77–84.
- Lull et Wright 1942, p. 50–93.
- Glut 1997, p. 389–396.
- Lambert et the Diagram Group 1990, p. 60.
- (en) John R. Horner, Weishampel, David B.; et Forster, Catherine A, The Dinosauria, Berkeley, University of California Press, , 2nd éd. (ISBN 0-520-24209-2), « Hadrosauridae », p. 438–463
- (en) A. Prieto-Marquez, G.M. Erickson et J.A. Ebersole, « A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs », Journal of Vertebrate Paleontology, vol. 36, no 2, , e1054495 (DOI 10.1080/02724634.2015.1054495)
- (en) P. Cruzado-Caballero and J. E. Powell. 2017. Bonapartesaurus rionegrensis, a new hadrosaurine dinosaur from South America: implications for phylogenetic and biogeographic relations with North America. Journal of Vertebrate Paleontology 37(2):e1289381:1-16
- (en) Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 225
- (en) Frederic A. Lucas, « The dinosaur Trachodon annectens », Smithsonian Miscellaneous Collections, vol. 45, , p. 317–320
- (en) William J. Morris, « Hadrosaurian dinosaur bills — morphology and function », Contributions in Science (Los Angeles County Museum of Natural History), vol. 193, , p. 1–14
- (en) Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 151–164
- Lull et Wright 1942, p. 110–117.
- (en) James A. Hopson, « The evolution of cranial display structures in hadrosaurian dinosaurs », Paleobiology, vol. 1, no 1, , p. 21–43
- (en) Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 128–130
- (en) Kathryn J. Stanton Thomas et and Carlson, Sandra J., « Microscale δ18O and δ13C isotopic analysis of an ontogenetic series of the hadrosaurid dinosaur Edmontosaurus: implications for physiology and ecology », Palaeogeography, Palaeoclimatology, and Palaeoecology, vol. 206, no 2004, , p. 257–287 (DOI 10.1016/j.palaeo.2004.01.007)
- (en) John H. Ostrom, « A reconsideration of the paleoecology of the hadrosaurian dinosaurs », American Journal of Science, vol. 262, , p. 975–997 (DOI 10.2475/ajs.262.8.975)
- (en) Peter M. Galton, « The posture of hadrosaurian dinosaurs », Journal of Paleontology, vol. 44, no 3, , p. 464–473
- (en) Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 98–110
- (en) Henry Fairfield Osborn, « The epidermis of an iguanodont dinosaur », Science, vol. 29, no 750, , p. 793–795 (PMID 17787819, DOI 10.1126/science.29.750.793)
- (en) Henry Fairfield Osborn, « Integument of the iguanodont dinosaur Trachodon », Memoirs of the American Museum of Natural History, vol. 1, , p. 33–54 (lire en ligne [PDF] (very large; 76,048 kb), consulté le )
- (en) « Mummified Dinosaur Unveiled », National Geographic News, (lire en ligne, consulté le )
- (en) Christopher Lee, « Scientists Get Rare Look at Dinosaur Soft Tissue », Washington Post, (lire en ligne, consulté le )
- (en) Phillip L. Manning et Morris, Peter M.; McMahon, Adam; Jones, Emrys; Gize, Andy; Macquaker, Joe H. S.; Wolff, G.; Thompson, Anu; Marshall, Jim; Taylor, Kevin G.; Lyson, Tyler; Gaskell, Simon; Reamtong, Onrapak; Sellers, William I.; van Dongen, Bart E.; Buckley, Mike; and Wogelius, Roy A., « Mineralized soft–tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA) », Proceedings of the Royal Society B, vol. 276, no 1672, , p. 3429–3437 (PMID 19570788, PMCID 2817188, DOI 10.1098/rspb.2009.0812)
- (en) Kenneth Carpenter, Fourth Symposium on Mesozoic Terrestrial Ecosystems, Drumheller, August 10–14, 1987, 3, Drumheller, Alberta, Royal Tyrrell Museum of Palaeontology, (ISBN 0-7732-0047-9), « Paleoecological significance of droughts during the Late Cretaceous of the Western Interior », p. 42–47
- (en) Kenneth Carpenter, « How to make a fossil: part 2 – Dinosaur mummies and other soft tissue », The Journal of Paleontological Sciences, vol. online, (lire en ligne [PDF], consulté le )
- (en) Robert T. Bakker, The Dinosaur Heresies : New Theories Unlocking the Mystery of the Dinosaurs and their Extinction, New York, William Morrow, , poche (ISBN 978-0-8217-2859-8, LCCN 88050951), « The case of the duckbill's hand », p. 146–159
- (en) John R. Horner, « A "segmented" epidermal frill in a species of hadrosaurian dinosaur », Journal of Paleontology, vol. 58, no 1, , p. 270–271
- (en) Tyler R. Lyson et Hanks, H. Douglas; and Tremain, Emily S., « New skin structures from a juvenile Edmontosaurus from the Late Cretaceous of North Dakota », Abstracts with Programs — Geological Society of America, vol. 35, no 2, , p. 13 (lire en ligne, consulté le )
- (en) Terry A. Gates et Sampson, Scott D., « A new species of Gryposaurus (Dinosauria: Hadrosauridae) from the late Campanian Kaiparowits Formation, southern Utah, USA », Zoological Journal of the Linnean Society, vol. 151, no 2, , p. 351–376 (DOI 10.1111/j.1096-3642.2007.00349.x)
- (en) Godefroit, P.; Bolotsky, Y. L.; Lauters, P. (2012). Joger, Ulrich, ed. "A New Saurolophine Dinosaur from the Latest Cretaceous of Far Eastern Russia". PLoS ONE 7 (5): e36849. DOI 10.1371/journal.pone.0036849. PMC 3364265.
- (en) David B. Weishampel, and Horner, Jack R., The Dinosauria, Berkeley, University of California Press, , 1st éd. (ISBN 0-520-06727-4), « Hadrosauridae », p. 534–561
- (en) Campione, N. & Evans, D. C., « Cranial growth and variation in Edmontosaurs (Dinosauria: Hadrosauridae): implications for latest Cretaceous megaherbivore diversity in North America », PLoS ONE, vol. 6, no 9, (DOI 10.1371/journal.pone.0025186, lire en ligne, consulté le )
- (en) Michael K. Brett-Surman, A revision of the Hadrosauridae (Reptilia : Ornithischia) and their evolution during the Campanian and Maastrichtian, Washington, D.C., George Washington University,
- (en) Benjamin S. Creisler, Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, Bloomington and Indianapolis, Indiana University Press, (ISBN 0-253-34817-X), « Deciphering duckbills: a history in nomenclature », p. 185–210
- (en) Edward Drinker Cope, « Supplement to the synopsis of the extinct Batrachia and Reptilia of North America », American Philosophical Society, Proceedings, vol. 12, no 86, , p. 41–52
- (en) Edward Drinker Cope, « Report on the stratigraphy and Pliocene vertebrate paleontology of northern Colorado », U.S. Geological and Geographical Survey of the Territories Annual Report, vol. 1, , p. 9–28
- (en) Kenneth Carpenter et and Young, D. Bruce, « Late Cretaceous dinosaurs from the Denver Basin, Colorado », Rocky Mountain Geology, vol. 37, no 2, , p. 237–254
- (en) Edward Drinker Cope, « Report on the vertebrate paleontology of Colorado », U.S. Geological and Geographical Survey of the Territories Annual Report, vol. 2, , p. 429–454
- (en) Lawrence M. Lambe, « On Vertebrata of the mid-Cretaceous of the Northwest Territory. 2. New genera and species from the Belly River Series (mid-Cretaceous). », Contributions to Canadian Paleontology, vol. 3, , p. 25–81
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 220–221.
- (en) Othniel Charles Marsh, « Notice of new reptiles from the Laramie Formation », American Journal of Science, vol. 43, , p. 449–453
- (en) Othniel Charles Marsh, « Restorations of Claosaurus and Ceratosaurus », American Journal of Science, vol. 44, , p. 343–349
- (en) John B. Hatcher, « The genus and species of the Trachodontidae (Hadrosauridae, Claosauridae) Marsh », Annals of the Carnegie Museum, part 14, vol. 1, , p. 377–386
- (en) Charles W. Gilmore, « On the genus Trachodon », Science, vol. 41, no 1061, , p. 658–660 (PMID 17747979, DOI 10.1126/science.41.1061.658)
- (en) M. A. Norell, Discovering Dinosaurs in the American Museum of Natural History, New York, Knopf, , 1re éd. (ISBN 978-0-679-43386-6, LCCN 94046225), p. 154–155
- (en) Cristiano Dal Sasso, Dinosaurs of Italy, Bloomington and Indianapolis, Indiana University Press, , 213 p. (ISBN 978-0-253-34514-1, LCCN 2004014815, lire en ligne), p. 112
- (en) Lawrence M. Lambe, « A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta », The Ottawa Naturalist, vol. 31, no 7, , p. 65–73 (lire en ligne [PDF] (entire volume, 18 mb), consulté le )
- (en) Lawrence M. Lambe, « The manus in a specimen of Trachodon from the Edmonton Formation of Alberta », The Ottawa Naturalist, vol. 27, , p. 21–25
- (en) Lawrence M. Lambe, « On the fore-limb of a carnivorous dinosaur from the Belly River Formation of Alberta, and a new genus of Ceratopsia from the same horizon, with remarks on the integument of some Cretaceous herbivorous dinosaurs », The Ottawa Naturalist, vol. 27, , p. 129–135
- (en) Michael K. Brett-Surman, The appendicular anatomy of hadrosaurian dinosaurs, Berkeley, University of California,
- (en) Michael K. Brett-Surman, « Phylogeny and paleobiogeography of hadrosaurian dinosaurs », Nature, vol. 277, no 5697, , p. 560–562 (DOI 10.1038/277560a0)
- (en) Donald F. Glut, The New Dinosaur Dictionary, Secaucus, NJ, Citadel Press, (ISBN 978-0-8065-0782-8), p. 49, 53
- (en) David Lambert, A Field Guide to Dinosaurs, New York, Avon Books, , 1re éd., 256 p., poche (ISBN 978-0-380-83519-5, LCCN 83002820), p. 156–161
- (en) Ralph E. Chapman, Dinosaur Systematics : Perspectives and Approaches, Cambridge, Cambridge University Press, , 356 p. (ISBN 978-0-521-43810-0, lire en ligne), « Morphometric observations on hadrosaurid ornithopods », p. 163–177
- (en) George Olshevsky, A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia, San Diego, Publications Requiring Research,
- Futura-Planète, « Un dinosaure momifié ! »
- « Edmontosaurus Dakota. Le dinosaure momifié »
- AFP, La Croix, « Au Muséum de Paris, l'art et la manière de "remonter" un dinosaure », 14/03/2018
- Exposition « Un T. rex à Paris », Muséum national d'histoire naturelle, Paris
- (en) Kraig Derstler, The Dinosaurs of Wyoming, Wyoming Geological Association, , « Dinosaurs of the Lance Formation in eastern Wyoming », p. 127–146
- Campione, N.E. (2009). "Cranial variation in Edmontosaurus (Hadrosauridae) from the Late Cretaceous of North America." North American Paleontological Convention (NAPC 2009): Abstracts, p. 95a.
- (en) Anna K. Behrensmeyer, « Evolution of Terrestrial Ecosystems Bonebed Database » [PDF], Bonebeds: Genesis, Analysis, and Paleobiological Significance, University of Chicago Press, (consulté le )
- (en) David B. Weishampel, Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M.P.; and Noto, Christopher N., The Dinosauria, Berkeley, University of California Press, , 2nd éd. (ISBN 0-520-24209-2), « Dinosaur distribution », p. 517–606
- (en) Peter Dodson, The Horned Dinosaurs : A Natural History, Princeton, Princeton University Press, , 360 p. (ISBN 978-0-691-05900-6, OCLC 60183976, lire en ligne), p. 14–15
- (en) Robert M. Sullivan, Late Cretaceous Vertebrates from the Western Interior, Albuquerque, New Mexico, New Mexico Museum of Natural History and Science, (lire en ligne [PDF]), « The Kirtlandian land-vertebrate "age" – faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America », p. 7–29
- (en) X-C. Wu et Brinkman, D.B.; Eberth, D.A.; and Braman, D.R., « A new ceratopsid dinosaur (Ornithischia) from the uppermost Horseshoe Canyon Formation (upper Maastrichtian), Alberta, Canada », Canadian Journal of Earth Science, vol. 44, no 9, , p. 1243–1265 (DOI 10.1139/E07-011)
- (en) David A. Eberth, « Review and comparison of Belly River Group and Edmonton Group stratigraphy and stratigraphic architecture in the southern Alberta Plains », Canadian Society of Petroleum Geology Diamond Jubilee Convention, Programs and Abstracts, vol. 117, , (cd) (lire en ligne [PDF], consulté le )
- (en) Dale A. Russell, An Odyssey in Time : Dinosaurs of North America, Minocqua, Wisconsin, NorthWord Press, Inc., (ISBN 978-1-55971-038-1), p. 170–171
- (en) Dale A. Russell et and Chamney, T. P., « Notes on the biostratigraphy of dinosaurian and microfossil faunas in the Edmonton Formation (Cretaceous), Alberta », National Museum of Canada Natural History Papers, vol. 35, , p. 1–35
- (en) Thomas M. Lehman, Mesozoic Vertebrate Life, Bloomington and Indianapolis, Indiana University Press, , 577 p. (ISBN 978-0-253-33907-2, LCCN 00053534), « Late Cretaceous dinosaur provinciality », p. 310–328
- Bakker, Robert T. (1986). The Dinosaur Heresies. p. 438.
- (en) Phillip Bigelow, « Cretaceous "Hell Creek Faunal Facies"; Late Maastrichtian » (consulté le )
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. p. 175–180
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. p. 180–181
- (en) Othniel Charles Marsh, « The skull and brain of Claosaurus », American Journal of Science, vol. 45, , p. 83–86
- (en) Othniel Charles Marsh, Sixteenth Annual report of the United States Geological Survey to the Secretary of the Interior, 1894–1895 : Part 1, Washington, D.C., U.S. Geological Survey, (lire en ligne), « The dinosaurs of North America », p. 133–244
- (en) Barnum Brown, « Anchiceratops, a new genus of horned dinosaurs from the Edmonton Cretaceous of Alberta, with discussion of the ceratopsian crest and the brain casts of Anchiceratops and Trachodon », Bulletin of the American Museum of Natural History, vol. 33, , p. 539–548
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 122–128.
- (en) Harry J. Jerison et Horner, John R.; et Horner, Celeste C, « Dinosaur forebrains », Journal of Vertebrate Paleontology, vol. 21, no 3, Suppl., , p. 64A
- (en) Peter M. Galton, « The cheeks of ornithischian dinosaurs », Lethaia, vol. 6, , p. 67–89 (DOI 10.1111/j.1502-3931.1973.tb00873.x)
- Fastovsky, D.E., and Smith, J.B. (2004). "Dinosaur paleoecology." The Dinosauria. p. 614–626.
- (en) Paul M. Barrett, « The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria) », Palaeontology, vol. 48, no 2, , p. 347–358 (DOI 10.1111/j.1475-4983.2005.00448.x)
- (en) David B. Weishampel, Evolution in jaw mechanics in ornithopod dinosaurs, Berlin; New York, Springer-Verlag, (ISBN 0-387-13114-0, ISSN 0301-5556, PMID 6464809)
- (en) Natalia Rybczynski et Tirabasso, Alex; Bloskie, Paul; Cuthbertson, Robin; and Holliday, Casey, « A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses », Palaeontologia Electronica, vol. 11, no 2, , online publication (lire en ligne, consulté le )
- (en) Casey M. Holliday et and Witmer, Lawrence M., « Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids », Journal of Vertebrate Paleontology, vol. 28, no 4, , p. 1073–1088 (DOI 10.1671/0272-4634-28.4.1073)
- (en) Vincent S. Williams et Barrett, Paul M.; and Purnell, Mark A., « Quantitative analysis of dental microwear in hadrosaurid dinosaurs,and the implications for hypotheses of jaw mechanics and feeding », Proceedings of the National Academy of Sciences, vol. 106, no 27, , p. 11194–11199 (PMID 19564603, PMCID 2708679, DOI 10.1073/pnas.0812631106)
- (en) Charles H. Sternberg, « A new Trachodon from the Laramie Beds of Converse County, Wyoming », Science, vol. 29, no 749, , p. 753–54
- (en) Philip J. Currie, Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers, Pékin, China Ocean Press, , 250 p. (ISBN 978-7-5027-3898-3), « "Stomach" contents of a hadrosaurid from the Dinosaur Park Formation (Campanian, Upper Cretaceous) of Alberta, Canada », p. 111–114
- (de) R. Kräusel, « Die Nahrung von Trachodon », Paläontologische Zeitschrift, vol. 4, , p. 80
- (de) O. Abel, « Diskussion zu den Vorträgen R. Kräusel and F. Versluys », Paläontologische Zeitschrift, vol. 4, , p. 87
- (en) G. R. Wieland, « Dinosaur feed », Science, vol. 61, no 1589, , p. 601–603 (PMID 17792714, DOI 10.1126/science.61.1589.601)
- (en) Justin S. Tweet et Chin, Karen; Braman, Dennis R.; and Murphy, Nate L., « Probable gut contents within a specimen of Brachylophosaurus canadensis (Dinosauria: Hadrosauridae) from the Upper Cretaceous Judith River Formation of Montana », PALAIOS, vol. 23, no 9, , p. 624–635 (DOI 10.2110/palo.2007.p07-044r)
- Rothschild et Tanke, D. H.; Helbling II, M.; and Martin, L.D. 2003, p. 495–500.
- (en) Bruce Rothschild, and Tanke, Darren H., Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, Bloomington and Indianapolis, Indiana University Press, (ISBN 0-253-34817-X), « Osteochondrosis is Late Cretaceous Hadrosauria », p. 171–183
- (en) W. I. Sellers et Manning, P. L.; Lyson, T.; Stevens, K.; and Margetts, L., « Virtual palaeontology: gait reconstruction of extinct vertebrates using high performance computing », Palaeontologia Electronica, vol. 12, no 3, , unpaginated (lire en ligne, consulté le )
- (en) M. K. Brett-Surman, James O. Farlow (éditeur) et M. K. Brett-Surman (éditeur), The Complete Dinosaur, Bloomington, Indiana University Press, , 752 p. (ISBN 0-253-33349-0), « Ornithopods », p. 330–346
- (en) Kenneth Carpenter, « Evidence of predatory behavior by theropod dinosaurs. », Gaia, vol. 15, , p. 135–144 (lire en ligne, consulté le ) [not printed until 2000]
- (en) Tony Campagna, « The PT interview: Michael Triebold », Prehistoric Times, vol. 40, , p. 18–19
- (en) Aase Roland Jacobsen et and Ryan, Michael J., « Taphonomic aspects of theropod tooth-marked bones from an Edmontosaurus bone bed (Lower Maastrichtian), Alberta, Canada », Journal of Vertebrate Paleontology, vol. 19, no 3, suppl., , p. 55A
- (en) « City Site Was Dinosaur Dining Room », ScienceDaily, ScienceDaily, (consulté le )
- (en) Arthur Chadwick et Spencer, Lee; and Turner, Larry, « Preliminary depositional model for an Upper Cretaceous Edmontosaurus bonebed », Journal of Vertebrate Paleontology, vol. 26, no 3, suppl., , p. 49A
- (en) Rebecca Gould et Larson, Robb; and Nellermoe, Ron, « An allometric study comparing metatarsal IIs in Edmontosaurus from a low-diversity hadrosaur bone bed in Corson Co., SD », Journal of Vertebrate Paleontology, vol. 23, no 3, suppl., , p. 56A–57A
- (en) Phil R. Bell et and Snively, E., « Polar dinosaurs on parade: a review of dinosaur migration », Alcheringa, vol. 32, no 3, , p. 271–284 (DOI 10.1080/03115510802096101)
- (en) Robin Lloyd, « Polar Dinosaurs Endured Cold Dark Winters », LiveScience.com, Imaginova, (consulté le )