Hypacrosaurus
Hypacrosaurus est un genre éteint et fossile de dinosaures ornithopodes de la famille des hadrosauridés et de la sous-famille des lambéosaurinés. Il comprend deux espèces, décrites à partir de fossiles datant du Crétacé supérieur, entre la partie supérieure du Campanien et le début du Maastrichtien, il y a environ entre 75 et 70 Ma[1] - [2]. Ses fossiles ont été retrouvés en Alberta, au Canada, ainsi que dans le Montana aux États-Unis.

Espèces de rang inférieur
- † H. altispinus Brown, 1913, espèce type
- † H. stebingeri Horner & Currie, 1994
Étymologie
Son nom de genre, Hypacrosaurus, signifie « presque le plus grand lézard » et vient du grec hypo- (ὑπό) qui signifie « moins » et de akros (ἄκρος) qui signifie « haut ». Ce nom a été donné car Hypacrosaurus était légèrement moins grand que Tyrannosaurus dont le squelette a été trouvé par le même homme, le paléontologue américain Barnum Brown[3] - [4]. Hypacrosaurus se caractérise principalement par sa crête osseuse, qui lui était vraisemblablement utile pour communiquer avec ses congénères. C'était un dinosaure herbivore, qui servait de proie aux grands tyrannosauridés.
Description
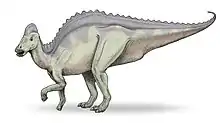
Parmi les dinosaures à bec de canard et à crête creuse (Lambeosaurinae), Hypacrosaurus est le plus facile à distinguer, notamment par la taille de ses épines neurales et la forme de sa crête. Ces épines neurales, qui font saillie en haut des vertèbres, mesurent cinq à sept fois la hauteur du corps de ces dernières[5], ce qui donne au dos d’Hypacrosaurus son allure caractéristique, tout en hauteur. Sa crête osseuse creuse est similaire à celle de son proche parent Corythosaurus mais elle est moins pointue au sommet, plus petite, plus large d'un côté à l'autre et possède un petit point osseux à l'arrière[5]. Contrairement aux autres lambéosaurinés, le passage du flux d'air dans la crête n'a pas la forme d'une courbe en « S » (du moins, pas dans le cas du H. altispinus)[6].
La taille d'un Hypacrosaurus adulte est estimée entre 9 et 10 mètres de long[5] - [7] et sa masse entre 2,5 et 4 tonnes[8] - [7].
Au niveau de son squelette, Hypacrosaurus ne se distingue des autres dinosaures à « bec de canard » que par certains détails de son bassin[9]. Comme les autres hadrosaures, Hypacrosaurus était un herbivore bipède pouvant également marcher sur quatre pattes. Les deux espèces connues, H. altispinus et H. stebingeri, n'ont pas été différenciées par la méthode dite de l'autapomorphie. H. stebingeri a de ce fait été décrit comme une transition entre le plus ancien Lambeosaurus et le plus récent Hypacrosaurus[10]. Des photographies du crâne d'un H. stebingeri adulte montrent un animal qui ressemble beaucoup à H. altispinus.
Classification
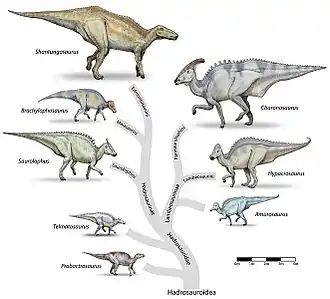
Hypacrosaurus était un hadrosaure de la sous-famille des Lambeosaurinae et a été reconnu comme tel dès la description de son crâne par Charles Whitney Gilmore en 1924[11]. Au sein des Lambeosaurinae, il est reconnu comme le plus proche parent de Lambeosaurus et de Corythosaurus[8]. Jack Horner et Phil Currie (1994) ont supposé que H. stebingeri est la transition entre Lambeosaurus et H. altispinus[10], et Michael K. Brett-Surman (1989) a lui supposé que Hypacrosaurus et Corythosaurus faisaient partie du même genre[12]. Ces genres, et plus particulièrement Corythosaurus et Hypacrosaurus, sont considérés comme la branche « casquée » ou « encapuchonnée » des lambéosaurinés et le clade qu'ils forment a été désigné un temps sous le nom informel de Corythosaurini[13] avant d'être englobé dans la tribu des Lambeosaurini[14]. Une nouvelle description de Nipponosaurus par Suzuki en 2004 trouva de proches relations entre Nipponosaurus et Hypacrosaurus stebingeri et conduisit à considérer que Hypacrosaurus était paraphylétique[15]. Cette description fut rejetée plus tard par une analyse plus complète des lambéosaurinés, montrant que les deux espèces d’Hypacrosaurus formaient un clade bien distinct des Nipponosaurus, mais incluant Corythosaurus et Olorotitan qui sont ses plus proches apparentés[16]. La validité de l'espèce H. stebingeri reste discutée (voir clagdogramme)[2].
Cladogramme
Le cladogramme ci-dessous, d'Albert Prieto-Márquez et ses collègues, réalisé en 2013, indique la position d'Hypacrosaurus dans la tribu des Lambeosaurini entre les genres Corythosaurus et Olorotitan[2]. La présence de guillemets pour l'espèce H. stebingeri montre que la position et la validité de ce taxon ne sont pas stabilisées.
| Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Un cladogramme plus large, plus complet et plus récent, établi par les mêmes auteurs en 2016, se trouve dans l'article Hadrosauridae[17].
Découverte et histoire
Origines

Les fossiles ayant été utilisés pour la description originale d’Hypacrosaurus ont été collectés en 1910 par Barnum Brown pour le compte du Musée américain d'histoire naturelle[9]. Ces restes comprennent des vertèbres cervicales et un morceau de bassin (AMNH 5204). Ils proviennent de sédiments connus sous le nom de formation de Horseshoe Canyon (Maastrichtien, Crétacé supérieur), et ont été retrouvés le long de la rivière de La Biche, en Alberta, au Canada. Brown décrivit en 1913 ces restes, avec d'autres os post-crâniens, comme ceux d'un nouveau genre qu'il nomma Saurolophus[3]. Aucun crâne n'était connu à ce moment, mais deux furent découverts et décrits peu de temps après[11].
Durant cette période, les restes de petits dinosaures à crête creuse et à bec de canards furent décrits et reliés à leur propre genre et espèce. Les premiers fossiles du genre d’Hypacrosaurus étaient, à l'époque de leur découverte, attribués au taxon Cheneosaurus tolmanensis. Ils comprenaient un crâne, des os de membres, des vertèbres ainsi que des os de bassin, tous provenant de la formation de Horseshoe Canyon[18]. Peu de temps après, Richard Swann Lull et Nelda Wright identifient un squelette (AMNH 5461) du Musée américain d'histoire naturelle provenant de la formation de Two Medicine dans le Montana comme appartenant au genre Procheneosaurus[19]. Ces taxons ainsi que d'autres furent considérés comme des genres valides jusque dans les années 1970, quand le paléontologue Peter Dodson démontra qu'il était plus probable que le genre Cheneosaurus était en fait composé de jeunes spécimens d'autres lambéosaurinés. Il a ensuite suggéré que Cheneosaurus était composé de jeunes individus d'Hypacrosaurus altispinus[20]. Bien que non formellement attestée, cette idée a été jugée acceptable[8]. Après de multiples observations, Procheneosaurus de la formation de Two Medicine se révélait pendant ce temps différent des autres spécimens de Procheneosaurus étudiés. On décida en 1994 de revoir son statut et de le considérer comme une espèce bien distincte, Hypacrosaurus stebingeri[10]. Les études phylogénétiques sur les Hadrosauroidea conduite par Prieto-Márquez et ses collègues en 2013 et 2016, montrent que H. stebingeri n'est pas si proche que cela de l'autre espèce du genre (H. altispinus)[2], voire très éloignée, tout en restant proche du genre Corythosaurus[17].
Espèces

Musée national de la nature et des sciences de Tokyo (Japon).
Hypacrosaurus altispinus, l'espèce type, est connue à partir de cinq à dix crânes articulés et quelques restes de squelettes associés issus d'individus jeunes et adultes. H. stebingeri est connu à partir d'un nombre inconnu mais important d’individus d'un âge allant de l'embryon à celui d'adulte[8]. L'hypothèse selon laquelle H. altispinus et H. stebingeri forment un groupe naturel excluant d'autres espèces d'hadrosaures peut être incorrecte. Comme indiqué dans la description de Nipponosaurus par Suzuki en 2004, leur analyse phylogénétique a révélé que Nipponosaurus était plus étroitement lié à H. altispinus que H. stebingeri l'était avec H. altispinus[15] mais cette dernière hypothèse a été rejetée par Evans et Reisz en 2007[16]. Pour Prieto-Márquez et ses collègues en 2016 elle serait proche des lambéosaurinés les plus évolués (Olorotitan, Blasisaurus et Amurosaurus)[17].
Hypacrosaurus stebingeri a été décrite à partir de divers morceaux de fossiles, incluant des nouveau-nés et des œufs ainsi que des nids trouvés dans des roches du Campanien supérieur de la formation de Two Medicine dans le Montana et près de la frontière avec l'Alberta. Ces restes représentent « la plus grande collection de squelettes de bébés hadrosaures, toutes espèces confondues »[10]. Son attribution au genre Hypacrosaurus est discutée[17].
Paléoécologie

H. altispinus a partagé l'environnement où s'est déposé la formation de Horseshoe Canyon avec d'autres hadrosauridés, Edmontosaurus et Saurolophus, l'hypsilophodontidé, Parksosaurus, l'ankylosauridé, Euoplocephalus, un nodosauridé, Edmontonia, les dinosaures à corne, Montanoceratops, Anchiceratops, Arrhinoceratops et Pachyrhinosaurus, le pachycéphalosaure Stegoceras, les ornithomimosaures Ornithomimus et Struthiomimus, une variété de théropodes mal connus incluant des troodontidés et des dromaeosauridés ainsi que les tyrannosauridés Albertosaurus et Daspletosaurus[21]. Les dinosaures issus de cette formation sont parfois appelés « Edmontoniens ». La formation de Horseshoe Canyon comprend différents milieux dont notamment des plaines inondables, des estuaires et des marécages, car durant une grande partie du Crétacé la voie maritime intérieure de l'Ouest formait une mer intérieure recouvrant toute la section centrale d'Amérique du Nord[22]. Néanmoins, H. altispinus préférait peut-être rester plus à l'intérieur des terres[8].
La formation de Two Medicine comprenait H. stebingeri, ainsi qu'un autre hadrosaure, Maiasaura, et un troodontidé Troodon. La présence de ces deux espèces a été établie à partir de la découverte de traces de nidification. Les tyrannosauridés Daspletosaurus, le caenagnathidé Chirostenotes, les dromaeosauridés Bambiraptor et Saurornitholestes, les dinosaures blindés Edmontonia et Euoplocephalus, l'hypsilophodonte Orodromeus, l'hadrosaure Prosaurolophus, les dinosaures à corne Achelousaurus, Brachyceratops, Einiosaurus et Styracosaurus ovatus étaient aussi présents[21]. Cette formation est plus éloignée de la voie maritime intérieure de l'Ouest, plus sèche et a une altitude plus élevée dans un climat plus continental[23].
Paléobiologie
Comme les autres hadrosauridés, Hypacrosaurus était certainement un herbivore bipède-quadrupède se nourrissant d'une grande variété de plantes. Son crâne lui permettait d'effectuer des mouvements de broyage analogues à la mastication, et ses dents, qui étaient renouvelées continuellement, étaient organisées en batteries dentaires contenant des centaines de dents, dont seule une poignée était en usage à tout moment, afin de les économiser. Il coupait les plantes avec son large bec puis les maintenait dans sa bouche avec des organes similaires à des joues. Grâce à son cou, Hypacrosaurus pouvait se nourrir des feuillages situés jusqu'à 4 mètres au-dessus du sol[8].
Nids et croissance

H. stebingeri pondait des œufs à peu près sphériques de 20 cm de haut pour 18,5 cm de diamètre, contenant des embryons de 60 cm de long.
Chaque portée comportait une vingtaine d'œufs. Une fois pondus dans des nids de sable ou de terre, ils étaient probablement recouverts de plantes ou de sable. Les femelles étaient en effet trop lourdes pour s'asseoir sur le nid sans risquer de briser son contenu[24]. Les jeunes et les embryons développés présentaient un crâne profond avec seulement une petite excroissance osseuse correspondant à leur future crête[10].
Selon une étude de 2017 basée sur les lignes de croissance incrémentielle de dents fossiles de certains dinosaures ornithischiens (Protoceratops andrewsi et Hypacrosaurus stebingeri) l'incubation était longue (3 à 6 mois avant l'éclosion[25]). Ce long délai pourrait d'ailleurs avoir contribué à la disparition des dinosaures non aviens face aux oiseaux et mammifères après l'extinction massive de la fin du Crétacé[26].
Les nouveau-nés mesuraient environ 1,70 mètre de long. Leur croissance était ensuite plus rapide que celles des alligators et comparable à celle des Struthioniformes. Selon Lisa Cooper et son équipe, H. stebingeri atteignait sûrement sa maturité sexuelle à l'âge de deux à trois ans, et sa taille définitive entre dix et douze ans[27].
À sa maturité sexuelle, la circonférence du fémur de ces animaux avait atteint 40 % de sa circonférence adulte. H. stebingeri avait vraisemblablement une croissance plus rapide que celle de ses principaux prédateurs, les tyrannosauridés Albertosaurus et Tyrannosaurus ; cette croissance rapide leur donnait de meilleures chances d'atteindre leur taille adulte et de pouvoir assurer une défense optimale face à ses prédateurs. Par ailleurs, une maturité sexuelle précoce assure de meilleures chances de survie à l'espèce, et correspond à une stratégie caractéristique des animaux servant de proies[28]. Du cartilage secondaire a été trouvé dans le squelette de spécimens de nouveau-nés d'H. stebingeri[29].
Fonctions de la crête

L'hypothèse principale sur la crête creuse d’Hypacrosaurus est la fonction sociale, constituant un signal visuel permettant à ses congénères d'identifier le sexe ou l'espèce. C'était peut-être aussi une sorte de caisse de résonance, amplifiant les cris émis par les animaux et facilitant donc leurs communications[8]. La crête et les passages nasaux associés ont également figuré dans le débat au sujet de l'endothermie des dinosaures, plus précisément dans les discussions portant sur les cornets nasaux.
Ces cornets sont des os ou des cartilages séparés en deux types, avec deux fonctions. Les cornets nasaux olfactifs, que l'on rencontre chez tous les tétrapodes vivants, possèdent une fonction olfactive. La fonction des cornets respiratoires est de prévenir la perte d'eau par évaporation. Seuls les oiseaux et les mammifères homéothermes modernes (animaux à sang chaud), en sont munis. Ils respirent plus fréquemment que les ectothermes (animaux à sang froid) afin de soutenir leur métabolisme plus élevé, et risquent plus de perdre de grandes quantités d'eau par la respiration, d'où cette adaptation[30]. Ruben et d'autres scientifiques ont conclu en 1996, à l'aide d'un scanner, que les cornets respiratoires n'étaient probablement pas présents chez Nanotyrannus, Ornithomimus ou Hypacrosaurus, il n'y a donc aucune preuve que ces animaux étaient à sang chaud[31].
Voir aussi
Liens externes
- (en) « Hypacrosaurus », sur http://nature.ca/ (the Canadian Museum of Nature).
- (en) « Hypacrosaurus », sur http://internt.nhm.ac.uk/ (The Natural History Museum's Dino Directory).
- Ressources relatives au vivant :
Références taxinomiques
- (en) Référence Paleobiology Database : Hypacrosaurus Brown, 1913 (éteint) (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023
Notes et références
- (en) V. M. Arbour, « A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus », Journal of Vertebrate Paleontology, vol. 29, no 4, , p. 1117–1135 (DOI 10.1671/039.029.0405)
- (en) Albert Prieto-Márquez, Fabio M. Dalla Vecchia, Rodrigo Gaete et Àngel Galobart, « Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis », PLoS ONE, vol. 8, no 7, , e69835 (DOI 10.1371/journal.pone.0069835)
- (en) Barnum Brown, « A new trachodont dinosaur, Hypacrosaurus, from the Edmonton Cretaceous of Alberta », Bulletin of the American Museum of Natural History, vol. 32, no 20, , p. 395–406 (lire en ligne)
- (en) Benjamin S. Creisler, Horns and Beaks : Ceratopsian and Ornithopod Dinosaurs, Bloomington et Indianapolis, Indiana University Press, , 185–210 p. (ISBN 978-0-253-34817-3 et 0-253-34817-X), « Deciphering duckbills »
- (en) Richard Swann Lull et Nelda E. Wright, Hadrosaurian Dinosaurs of North America, Geological Society of America, coll. « Geological Society of America Special Paper 40 », , p. 206–208.
- (en) David B. Weishampel, « The nasal cavity of lambeosaurine hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies », Journal of Paleontology, vol. 55, no 5, , p. 1046–1057
- (en) Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- (en) Jack Horner, David B. Weishampel et Catherine A. Forster, The Dinosauria, Berkeley, University of California Press, , 2e éd. (ISBN 0-520-24209-2), « Hadrosauridae », p. 438–463
- (en) Donald F. Glut, Dinosaurs : The Encyclopedia, Jefferson, Caroline du Nord, McFarland & Co, , 1076 p. (ISBN 0-89950-917-7), « Hypacrosaurus », p. 478–482.
- (en) Jack Horner et Phillip J. Currie, Dinosaur Eggs and Babies, Cambridge, Cambridge University Press, (ISBN 0-521-56723-8), « Embryonic and neonatal morphology and ontogeny of a new species of Hypacrosaurus (Ornithischia, Lambeosauridae) from Montana and Alberta », p. 312–336
- (en) Charles Whitney Gilmore, « On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Parks », Canada Department of Mines Geological Survey Bulletin (Geological Series), vol. 38, no 43, , p. 29–48
- (en) Michael K. Brett-Surman, A revision of the Hadrosauridae (Reptilia : Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation, Washington, D.C., Graduate School of Arts and Sciences of The George Washington University, , 272 p.
- (en) Cruzado-Caballero P, Pereda-Suberbiola X, Ruiz-Omeñaca JI (2010a) Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (Hadrosauridae) from the Latest Cretaceous of Arén (Huesca, Spain). Can J Earth Sci 47: 1507–1517
- (en) R. M. Sullivan, S. E. Jasinski, M. Guenther and S. G. Lucas. 2011. The first lambeosaurin (Dinosauria, Hadrosauridae, Lambeosaurinae) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico. Fossil Record 3. New Mexico Museum of Natural History and Science Bulletin 405-417
- (en) Daisuke Suzuki, « Nipponosaurus sachaliensis (Dinosauria; Ornithopoda): anatomy and systematic position within Hadrosauridae », Journal of Vertebrate Paleontology, vol. 24, no 1, , p. 145–164 (DOI 10.1671/A1034-11)
- (en) David C. Evans et Robert R. Reisz, « Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta », Journal of Vertebrate Paleontology, vol. 27, no 2, , p. 373–393 (DOI 10.1671/0272-4634(2007)27[373:AAROLM]2.0.CO;2)
- (en) A. Prieto-Marquez, G.M. Erickson et J.A. Ebersole, « A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs », Journal of Vertebrate Paleontology, vol. 36, no 2, , e1054495 (DOI 10.1080/02724634.2015.1054495)
- (en) Lawrence M. Lambe, « On Cheneosaurus tolmanensis, a new genus and species of trachodont dinosaur from the Edmonton Cretaceous of Alberta », The Ottawa Naturalist, vol. 30, no 10, , p. 117–123
- (en) William Diller Matthew, « Canadian dinosaurs », Natural History, vol. 20, no 5, , p. 1–162
- (en) Peter Dodson, « Taxonomic implications of relative growth in lambeosaurine dinosaurs », Systematic Zoology, vol. 24, no 1, , p. 37–54 (DOI 10.2307/2412696, JSTOR 2412696)
- (en) David B. Weishampel, Paul M. Barrett, Rodolfo A. Coria, Le Jean Loueff, Xu Xing, Zhao Xijin, Ashok Sahni, Elizabeth M. P. Gomani et Christopher N. Noto, The Dinosauria, Berkeley, University of California Press, , 2e éd. (ISBN 0-520-24209-2), « Dinosaur distribution », p. 517–606
- (en) Peter Dodson, The Horned Dinosaurs : A Natural History, Princeton, Princeton University Press, , 14–15 p. (ISBN 0-691-05900-4, lire en ligne).
- (en) Raymond R. Rogers, « Taphonomy of three dinosaur bone beds in the Upper Cretaceous Two Medicine Formation of northwestern Montana: evidence for drought-related mortality », PALAIOS, vol. 5, no 5, , p. 394–413 (DOI 10.2307/3514834, JSTOR 3514834)
- (en) « Meet the Hypacrosaur Family », sur Children's museum Dinosphere (consulté le )
- (en) Carolyn Gramling, « Dinosaur babies took a long time to break out of their shells », sur Science,
- (en) Gregory M. Erickson, Darta K. Zelenitski, David Ian Kay et Mark A. Norell, « Dinosaur incubation periods directly determined from growth-line counts in embryonic teeth show reptilian-grade development », PNAS, (DOI 10.1073/pnas.1613716114, résumé).
- (en) Lisa N. Cooper, « Growth rate of Hypacrosaurus stebingeri as hypothesized from lines of arrested growth and whole femur circumference », Journal of Vertebrate Paleontology, vol. 19, no 3, Suppl., , p. 35
- (en) Lisa N. Cooper, « Relative growth rates of predator and prey dinosaurs reflect effects of predation », Proceedings of the Royal Society B, vol. 275, no 1651, , p. 2609–2615 (PMID 18682367, PMCID 2605812, DOI 10.1098/rspb.2008.0912)
- (en) Alida M. Bailleul, Brian K. Hall et John R. Horner, « First Evidence of Dinosaurian Secondary Cartilage in the Post-Hatching Skull of Hypacrosaurus stebingeri (Dinosauria, Ornithischia) », PLoS ONE, vol. 7, no 4, , e36112 (DOI 10.1371/journal.pone.0036112).
- (en) Anusuya Chinsamy et Willem J. Hillenius, The Dinosauria, , 2e éd., « Physiology of nonavian dinosaurs », p. 643-659
- (en) J. A. Ruben, W. J. Hillenius, N. R. Geist, A. Leitch, T. D. Jones, P. J. Currie et J. R. Horner, « The metabolic status of some Late Cretaceous dinosaurs », Science, vol. 273, no 5279, , p. 1204–1207 (DOI 10.1126/science.273.5279.1204)