Parasaurolophus
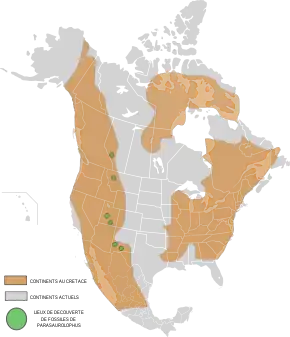
Parasaurolophus est un genre éteint et fossile de dinosaures appartenant à l'ordre des ornithischiens, au sous-ordre des ornithopodes et à la famille des hadrosauridés. Ces animaux ont vécu au Crétacé supérieur, durant une partie du Campanien, il y a entre 76,6 et 73 Ma (millions d'années), dans l'actuelle Amérique du Nord. Au moins une quinzaine de fossiles ont été découverts en 2008. Ils sont répartis en trois espèces nommées P. walkeri, P. tubicen et P. cyrtocristatus. Ce dinosaure a été décrit pour la première fois en 1922 par William Parks à partir d'un crâne et d'un squelette incomplet découverts dans la province de l'Alberta au Canada dans la formation de Dinosaur Park. Le plus au sud des fossiles découverts l'a été au Nouveau-Mexique, le plus au nord en Alberta.

Répartition géographique
Espèces de rang inférieur
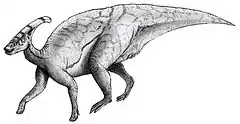
Parasaurolophus est un dinosaure de la famille des Hadrosauridae, connue pour les ornementations variées de leur tête. Parasaurolophus présente une grande crête qui ressemble à un long tube incurvé vers l'arrière partant du haut du crâne. L'hadrosauridé Charonosaurus, originaire quant à lui de la région couverte aujourd'hui par la Chine, et qui est considéré comme son proche parent connu, avait un crâne semblable et, une crête potentiellement semblable. La crête de Parasaurolophus est un sujet de débat entre scientifiques, le consensus sur sa fonction comporte : la reconnaissance visuelle à la fois des deux espèces et des deux sexes, amplificateur acoustique, ou encore système de thermorégulation. C'est un des « dinosaures à bec de canard » les plus rares, connu seulement à partir d'une poignée de bons spécimens.
Étymologie
Parasaurolophus a été formé à partir de trois termes de grec ancien, παρα [para] « près, parallèle », σαυρος [saurus] « lézard » et λοφος [lophos] « crête[2] », ce qui peut s'interpréter par « reptile à crêtes à cloisons parallèles[3] ».
William Parks a nommé le spécimen P. walkeri en honneur à Byron Edmund Walker, président du conseil d'administration des Trustees du Musée royal de l'Ontario[4].
Le nom scientifique tubicen est le nom d'une trompette romaine qui soufflait dans un long tuba[5]. Des restes d'au moins trois spécimens de P. tubicen ont été découverts au Nouveau-Mexique[6].
Le terme cyrtocristatus a été formé à partir de deux racines latines, cyrto qui signifie « courbé[7] » et de cristatus qui signifie « huppé ».
Description

Comme pour de nombreux dinosaures, le squelette de Parasaurolophus n'est pas connu dans sa totalité. Sa morphologie n'est donc pas exactement connue. La longueur du spécimen type de P. walkeri est estimée à 9,50 m pour un crâne mesurant environ 1,60 m de long (crête incluse), alors que celui de P. tubicen est estimé à plus de 2 m ce qui indique un animal plus grand[8]. Sa masse est estimée à 2,5 tonnes[1]. Le seul membre antérieur connu du spécimen type est relativement court pour un hadrosaure, avec une petite mais large omoplate. Ceci tend à montrer que les hadrosaures descendent d'animaux bipèdes. Le fémur de P. walkeri mesure 103 cm, et est très robuste par rapport à sa longueur, comparé aux autres hadrosaures[9]. L'humérus et le bassin sont aussi très robustes[10]. Parasaurolophus a la crête la plus longue de tous les hadrosauridés, puisqu'elle mesure environ 1,80 mètre de long[11].
Tout comme les autres hadrosaures, Parasaurolophus était capable de marcher sur ses deux pattes postérieures, ou à quatre pattes. Il préférait probablement rechercher sa nourriture à quatre pattes, mais courir sur deux pattes[6]. Les processus épineux des vertèbres étaient longs, chose assez commune chez les lambéosaurinés[9] ; très grands au-dessus des hanches, ils augmentaient la hauteur du dos. L'aspect de la peau est connue chez P. walkeri, avec des écailles uniformes, telles de petites protubérances, mais pas de plus grandes structures[4].
L'attribut le plus reconnaissable de Parasaurolophus, était sa crête, qui sortait de l'arrière de sa tête, et qui était un prolongement de l'os nasal et du prémaxillaire. Le spécimen type de P. walkeri avait une encoche sur les processus épineux près de l'endroit où l'extrémité de la crête aurait cogné le dos, mais c'était peut-être une pathologie spécifique de cet individu[9]. William Parks, qui a nommé le genre, a supposé qu'un ligament partait de la crête jusqu'à l'encoche et servait à soutenir la tête[4]. Bien que cette idée semble peu probable[12], il arrive que Parasaurolophus soit représenté avec un pan de peau allant de la crête au cou. Vu que cette encoche se trouve exactement à l'endroit où la crête touchait le dos quand Parasaurolophus penchait la tête en arrière, il se peut qu'elle serve à laisser la place à la crête qui autrement gênerait les mouvements de l'animal[13].
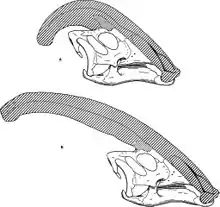
La crête était creuse, avec des conduits distincts allant de chaque narine à l'extrémité de la crête avant de faire demi-tour, redescendre la crête et se diriger vers l'intérieur du crâne. Les conduits étaient plus simples chez P. walkeri, mais beaucoup plus complexes chez P. tubicen, où certains conduits étaient bouchés à une extrémité, et d'autres se fusionnaient et s'embranchaient[14]. Alors que P. walkeri et P. tubicen avaient de longues crêtes avec une légère courbe, P. cyrtocristatus avait une courte crête avec un profil plus circulaire[15].
Le cou du Parasaurolophus était sans doute fortement incurvé, comme celui des bisons actuels, et pourvu d'énormes os. Il pouvait marcher aussi bien sur quatre pattes que sur deux pattes, mais son dos restait horizontal et non pas vertical, comme on l'a cru longtemps. Et son énorme queue ne traînait pas par terre[16].
On a longtemps pensé que Parasaurolophus se servait de sa gigantesque queue pour nager, mais on a découvert des fossiles de Parasaurolophus dans des roches dont la nature bien particulière suggère qu'il vivait dans un environnement terrestre sec, semblable à l'habitat des éléphants. De plus, la manière dont les os de la queue s'emboîtent montre que ce dinosaure n'était pas capable d'effectuer de puissants mouvements latéraux avec sa queue, nécessaires pour pouvoir nager. Les contenus d'estomacs fossilisés retrouvés contiennent aussi principalement des plantes terrestres. Il ne vivait donc pas dans les alentours des fleuves et des lacs du Crétacé supérieur, mais plutôt sur la terre ferme, se nourrissant de plantes coriaces.
Certains doigts de Parausorolophus retrouvés semblent être palmés, mais certains paléontologues estiment que ce sont des conséquences de la fossilisation[13].
Systématique
Liste des espèces
Classification


Comme son nom l'indique, les scientifiques rapprochaient Parasaurolophus de Saurolophus, en raison de leur crête semblable[4]. Dès 1924, il a été vite reconnu par Gilmore comme un membre de la sous-famille des lambéosaurinés[1], Saurolophus étant quant à lui un hadrosauriné[17]. Parasaurolophus est habituellement considéré comme d'une lignée séparée de lambéosaurinés, une lignée distincte de celle des autres dinosaures à crête tels que Corythosaurus, Hypacrosaurus, et Lambeosaurus[6] - [18] - [19]. Son parent le plus proche est Charonosaurus, avec un crâne semblable mais cependant avec une crête incomplète et découvert dans la région du fleuve Amour au nord-est de la Chine[20] et ces deux taxons ont été regroupés dans un clade, la tribu des Parasaurolophini[19] - [21] - [22]. Les fossiles de P. cyrtocristatus, avec leur petite crête un peu plus ronde, pourraient être le taxon le plus basal des trois espèces connues de Parasaurolophus[19], ou bien pourraient être des jeunes ou encore des femelles de P. tubicen[23].
Taxinomie
Un seul spécimen de l'holotype P. walkeri a été découvert[6]. Il diffère de P. tubicen par les tubes situés dans sa crête[14], et de P. cyrtocristatus par sa longue crête et un humérus plus long que son radius[15]. Trois spécimens possibles du P. cyrtocristatus ont été découverts au Nouveau-Mexique et dans l'Utah. Ces spécimens sont plus petits et leur crête est courte et ronde[14]. Leur petite taille et la forme de leur crête laissent à penser qu'ils pourraient être des jeunes ou des femelles du P. tubicen qui date à peu près de la même époque et de la même formation au Nouveau-Mexique. Comme l'a noté Thomas Williamson le matériel type de P. cyrtocristatus est environ 72 % plus petit que P. tubicen, et c'est environ la taille où les autres lambeosaurinés commencent à montrer un dimorphisme sexuel au niveau de leur crête (~70 % de la taille d'un adulte)[23]. Cette proposition a été rejetée par plusieurs études récentes sur les Lambeosaurinae[6] - [19].
P. tubicen est la plus grande espèce, avec dans sa crête des passages d'air plus complexes que ceux du P. walkeri, et une crête plus longue et plus fine que celle de P. cyrtocristatus[14].
Phylogénie
Le cladogramme d'Albert Prieto-Márquez et ses collègues, réalisé en 2013, indique la position de Parasaurolophus au sein de la tribu des Parasaurolophini, en groupe frère avec le genre chinois Charonosaurus[21] :
| Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||
Un cladogramme plus large, plus complet et plus récent, établi par les mêmes auteurs en 2016, se trouve dans l'article Hadrosauridae[22].
Découverte et datation
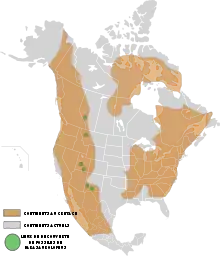

L'holotype 768 du Musée royal de l'Ontario comporte un crâne et un squelette partiel auquel manque une grande partie de la queue ainsi que les membres postérieurs en dessous des genoux. Il a été trouvé par une équipe de l'université de Toronto, en 1920, près de Sand Creek le long de la Red Deer dans la province canadienne d'Alberta[4]. Ces sédiments font partie de la formation de Dinosaur Park datée de la partie moyenne du Campanien, entre 76 et 74,8 million d'années[24] - [25]. William Parks donna le nom de P. walkeri à ce spécimen en hommage à Sir Byron Edmund Walker, Chairman of the Board of Trustees du Royal Ontario Museum[4]. Outre ce spécimen, les fossiles de Parasauropholus sont rares dans l'Alberta[26], qui compte seulement un autre crâne partiel provenant de la formation de Dinosaur Park[27] et trois autres spécimens sans crâne appartenant sans doute au genre[28].
En 1921, Charles H. Sternberg a découvert une partie d'un crâne (PMU.R1250) dans le comté de San Juan (Nouveau-Mexique), issu du membre De-na-zin de la formation géologique de Kirtland, dont l'âge est de 73,4 à 73 Ma (millions d'années), soit un peu plus récent que celui la formation précédente[29]. Le spécimen a été envoyé à Uppsala en Suède, où en 1931, Carl Wiman l'a décrit comme étant une deuxième espèce, P. tubicen[30]. Un second crâne quasi complet de P. tubicen (NMMNH P-25100) a été découvert au Nouveau-Mexique en 1995. En 1999, à la suite d'une étude utilisant la technique de la tomodensitométrie sur ce crâne, Robert Sullivan et Thomas Williamson ont écrit une monographie, couvrant les aspects de son anatomie et de sa taxinomie, ainsi que les fonctions de sa crête[14]. Williamson a publié ensuite une revue indépendante sur les restes, où il expose son désaccord sur les conclusions taxinomiques[23].
John Ostrom a décrit un autre spécimen intéressant (FMNH P27393), découvert au Nouveau-Mexique, en 1961, comme étant une nouvelle espèce : P. cyrtocristatus. C'est un crâne presque complet avec une petite crête ronde, et la quasi-totalité du reste d'un corps, à l'exception des pieds, du cou et d'une partie de la queue[15]. Ce spécimen a été enseveli soit à la fin de la période de dépôt de la formation de Fruitland, ou plus probablement au début de celle de la formation de Kirtland, soit un âge de 74,5 Ma (millions d'années) dans le Campanien[14] - [29].
La zone d'extension de ces espèces a été étendue en 1979, lorsque David B. Weishampel et James A. Jensen ont décrit un crâne partiel disposant d'une crête semblable (BYU 2467) datant également du Campanien et découvert dans la formation de Kaiparowits, située dans le comté de Garfield (Utah) et datée entre 76,6 et 74,5 Ma (millions d'années)[31] - [32]. Depuis, un autre crâne avec une morphologie de petite crête ronde comme celle de P. cyrtocristatus a été découvert dans l'Utah[14].
La formation de Kaiparowits en Utah a aussi fourni en 2013 un crâne et un squelette d'un très jeune Parasaurolophus de 2,50 mètres de long, soit le quart de sa taille adulte ; ce spécimen a été surnommé « Joe »[33]. Il constitue à la fois le squelette le plus complet et celui du plus jeune individu du genre. Les coupes histologiques réalisées dans les os n'ont révélé aucun arrêt de lignes de croissance, suggérant que l'animal pourrait être mort durant sa première année[33]. Sa crête est réduite, globalement hémisphérique, très peu développée en proportion du crâne qui lui mesure déjà 24,6 centimètres de long[33]. Le crâne de « Joe » a été étudié avec des scanners médicaux qui ont confirmé le rôle de la crête dans l'émission des sons. Si les Parasaurolophus adultes peuvent émettre des sons de type haut-parleurs graves (boomer ou woofer), la crête du jeune « Joe », courte et petite, ne pouvait émettre que des sons bien plus aigus (fréquences sonores élevées) de type tweeter[34].
Paléoécologie
Parasaurolophus walkeri, de la formation de Dinosaur Park, était un membre d'une faune préhistorique variée et bien documentée, incluant des dinosaures bien connus comme les cératopsidés Centrosaurus, Styracosaurus et Chasmosaurus, d'autres dinosaures à bec de canard comme Prosaurolophus, Gryposaurus, Corythosaurus et Lambeosaurus; le tyrannosauridé Gorgosaurus; et les dinosaures à armures comme Edmontonia et Euoplocephalus[35]. C'était un spécimen rare de cette faune[26]. La formation de Dinosaur Park est interprétée comme étant une région peu accidentée, couverte de rivières et de vallées d'inondation, qui est devenue plus marécageuse, et influencée par les conditions marines dues, avec le temps, à la transgression vers l'ouest de la voie maritime intérieure de l'Ouest, mer divisant en deux l'Amérique du Nord durant cette époque[36]. Le climat était plus chaud que celui de l'actuelle Alberta, sans gelée, mais avec des saisons plus humides et plus sèches. Les conifères étaient a priori les plantes dominantes de la canopée, avec un sous-bois de fougères, de fougères arborescentes et d'angiospermes[37]. Les Ceratopsidae tels les Chasmosaurinae y étaient a priori plus répandus que les hadrosauridés[38].
Les espèces du Nouveau-Mexique partageaient leur environnement avec le grand sauropode Alamosaurus, le dinosaure à bec de canard Kritosaurus, le ceratopsidé Pentaceratops, des dinosaures à armure comme Nodocephalosaurus, les Saurornitholestes, et des tyrannosauridés pour l'instant non identifiés[35].
Face à ses prédateurs, Parasaurolophus ne possédait pas de défenses naturelles (contrairement à Iguanodon par exemple), mais on suppose qu'il possédait une assez bonne vue et une bonne ouïe ce qui lui permettait de détecter sans peine ses prédateurs à distance[13].
Paléobiologie
Alimentation
Comme les autres hadrosauridés, Parasaurolophus était un grand herbivore bipède/quadrupède, s'alimentant de plantes et son crâne sophistiqué lui permettait un mouvement de broyage de sa nourriture analogue à la rumination. Ses dents jugales étaient constamment remplacées et formaient des batteries dentaires qui contenaient des centaines de dents, mais seulement une poignée d'entre elles étaient utilisées en même temps. Les plantes devaient être coupées par son large bec et maintenues dans la mâchoire par un organe semblable à la joue, ce qui permettait d'attaquer les plantes les plus coriaces. Contrairement aux autres reptiles qui ne mâchent généralement pas leur nourriture, puisqu'ils se contentent d'avaler, Parasaurolophus comme les autres ornithopodes, pouvait mâcher sa nourriture, comme le font les mammifères. Il pouvait s'alimenter de feuillages situés jusqu'à 4 mètres de hauteur[6]. Comme l'a noté Bob Bakker, les lambéosaurinés avaient des becs plus étroits que les hadrosaurinés, ce qui implique que Parasaurolophus pouvait mieux sélectionner son alimentation que ses cousins au bec large et dénués de crête[39].
Crête nasale
Plusieurs hypothèses sur les fonctions de la crête dite « nasale » de Parasaurolophus ont été avancées, mais elles n'ont pas été jugées crédibles pour la plupart[40] - [12]. Aujourd'hui, les hypothèses ayant cours sont l'identification du sexe ou de l'espèce, l'amplification sonore facilitant la communication et la thermorégulation. Cependant, on ne sait pas clairement parmi ces fonctions laquelle est la plus importante et à quels moments de leur croissance[41].
Différences entre les espèces et étapes de croissance

Comme pour les autres lambéosaurinés, on pense que la crête crânienne de Parasaurolophus changeait avec l'âge et qu'elle était caractéristique d'un dimorphisme sexuel chez les adultes. James Hopson, un des premiers chercheurs à avoir décrit les crêtes des lambeosaurinés pense que le P. cyrtocristatus avec sa petite crête était une femelle de P. tubicen[42]. Thomas Williamson croit plutôt que c'était un jeune P. tubicen[23]. Aucune de ces deux hypothèses n'a été unanimement reconnue. Comme seulement six bons crânes et une boîte crânienne d'un jeune ont été découverts, la mise au jour de nouveaux fossiles permettrait vraisemblablement de faire la lumière sur cette relation éventuelle. Williamson a noté que dans n'importe quel cas, le jeune Parasaurolophus possédait sûrement une crête petite et ronde comme P. cyrtocristatus, qui grandissait probablement de plus en plus vite à mesure que les individus approchaient la maturité sexuelle[23]. De nouvelles études plus récentes sur la boîte crânienne qui était au départ attribuée au Lambeosaurus, puis à présent à Parasaurolophus apportent des preuves de l'existence d'une petite crête tubulaire chez les jeunes.
Ce spécimen possède une petite dilatation vers le haut des os frontaux, qui était similaire mais plus petite que pour les spécimens adultes ; chez les adultes, les os frontaux formaient une plateforme qui supportait la base de la crête. Ce spécimen indique aussi que la croissance de la crête chez Parasaurolophus et le profil facial des jeunes individus diffèrent du modèle Corythosaurus-Hypacrosaurus-Lambeosaurus, en partie à cause du manque d'un fin cocon osseux dans la crête qui composait la partie supérieure de la crête chez les trois autres lambeosaurinés[27].
Hypothèses rejetées sur les fonctions

Des propositions se concentrent sur les adaptations à la vie aquatique, suivant l'hypothèse que les hadrosauridés étaient amphibiens. Ainsi, Alfred Sherwood Romer a suggéré en 1993 que cette crête permettait à l'animal de plonger en apnée[43], Martin Wilfarth pensait que c'était une attache pour une trompe mobile utilisée comme tuba et à attraper la nourriture[44], Charles Mortram Sternberg croyait que leur crête servait de pompe à air pour empêcher l'eau d'entrer dans les poumons[45], et Edwin Harris Colbert pensait qu'elle servait de réservoir à air pour plonger sous l'eau[46].
William Parks a proposé qu'elle était rattachée aux vertèbres par des ligaments ou des muscles, et aidait à bouger et à supporter la tête[4]. Othenio Abel a suggéré qu'elle était utilisée comme arme dans les combats entre ces espèces[47], et Angela Milner suggère qu'elle pouvait servir de déflecteur de feuilles, comme le casque des casuariidés[40].
Il a été également suggéré que la principale fonction de la crête était d'abriter des organes spéciaux. Halszka Osmólska a proposé qu'elle abritait des glandes à sel[48], et John Ostrom a proposé qu'elle abritait des zones plus larges pour l'épithélium olfactif permettant ainsi d'améliorer l'odorat des lambeosaurinés, qui n'avait pas vraiment de facultés pour se défendre[49]. Duane Gish a formulé, quant à lui, une proposition peu courante : la crête aurait abrité des glandes chimiques permettant à Parasaurolophus de cracher des jets de feu chimique à ses ennemis, comme l'actuel coléoptère bombardier[50].
La plupart de ces hypothèses ont été discréditées ou rejetées[12]. Par exemple, il n'y a aucun trou au bout de la crête permettant à l'animal de plonger en apnée. Il n'y aucune cicatrice de muscle pour une trompe et il serait même étrange qu'un animal avec un bec en ait besoin. Si elle avait eu comme fonction de servir de pompe à air elle n'aurait à ce moment-là pas laissé s'infiltrer l'eau. Un réservoir d'air n'aurait pas pu être suffisant pour un animal de la taille d'un Parasaurolophus. D'autres hadrosauridés possédaient de grande tête sans avoir besoin d'une large crête creuse pour servir de point d'attache à des ligaments[49]. Ainsi, aucune des propositions expliquent pourquoi la crête possède une telle forme, pourquoi les autres lambeosaurinés avaient des crêtes si différentes mais possédant des fonctions similaires, comment les hadrosauridés sans crête ou avec des crêtes solides pouvaient vivre sans de telles capacités, ou pourquoi certains hadrosauridés possédaient des crêtes solides.
Ces considérations ont un impact particulier sur les hypothèses basées sur l'amélioration des facultés du système déjà présentes chez l'animal, comme les hypothèses sur les glandes à sel ou l'olfaction[40], et démontrent qu'elles n'étaient pas les fonctions principales de la crête. De plus, les travaux sur les cavités nasales des lambeosaurinés montrent que les nerfs olfactifs et les tissus sensoriels étaient à l'extérieur de la portion des sinus dans la crête, donc l'accroissement de la crête a peu de chose à voir avec le sens de l'odorat[41].
Fonctions sociales
De plus en plus d'auteurs soutiennent que la crête devait avoir des fonctions sociales et des fonctions physiologiques permettant l'identification, la communication visuelle et auditive. Il est clair que cette imposante crête est un signal visuel qui différencie chaque animal de ses voisins. La large orbite de l'hadrosauridé et la présence d'un anneau sclérotique suggèrent que l'animal possédait une vision perçante et des habitudes diurnes, et donc que la vue était un sens important chez ces animaux. Si, comme il apparaît sur certaines illustrations, une peau s'étendait de la crête au cou ou jusqu'au dos, l'exposition visuelle aurait été encore plus voyante[42]. Comme le laissent supposer les autres crânes des lambeosaurinés, la taille et la forme de la crête de Parasaurolophus permettaient probablement aux animaux de se reconnaître en se distinguant par exemple du Corythosaurus ou du Lambeosaurus[41].
Fonction sonore
L'apparence externe de la crête cache une anatomie interne complexe. Les sinus nasaux qui la composent suggèrent l'existence d'une autre fonction[41]. Carl Wiman fut le premier à proposer en 1931 que ces sinus servaient de signalement auditif, comme un cromorne[30] ; Hopson et David B. Weishampel ont revisité cette idée dans les années 1970 et 1980[42] - [51] - [52]. Hopson a fourni des preuves montrant que les hadrosauridés avaient une bonne audition. Il y a au moins un exemple chez Corythosaurus qui possède de minces stapes reptiliens combinés avec un large espace pour un tympan indiquant une oreille moyenne sensible. De plus, le conduit de la cochlée des hadrosauridés est allongé comme celui des crocodiliens, indiquant que la portion auditive de l'oreille interne était bien développée[42]. Weishampel a estimé que P. walkeri était capable de produire des fréquences de 48 à 240 Hz, et lP. cyrtocristatus de 75 à 375 Hz. En se basant sur les similitudes entre les oreilles internes des hadrosauridés et celles des crocodiliens, il a également proposé que les adultes hadrosauridés étaient sensibles aux hautes fréquences, comme celles que leur progéniture pouvait produire. Selon Weishampel, ce fait est en adéquation avec le nécessaire besoin de communication entre les parents et leurs progénitures[51].
Un modèle informatisé du spécimen basé sur un crâne de P. tubicen en bonne conservation, espèce qui possédait des sinus plus complexes que celle de P. walkeri, a permis la simulation des divers sons que la crête pouvait produire[53].
Le principal canal possédait une fréquence de résonance à 30 Hz, mais l'anatomie complexe des sinus devait créer des modulations des sons[54].
Fonction de thermorégulation
La large surface et la vascularisation de la crête suggèrent qu'elle avait une fonction thermorégulatrice[55]. P.E. Wheeler a proposé le premier cette utilisation en 1978 : la crête permettrait de préserver le cerveau de la chaleur[56]. Teresa Maryańska et Halszka Osmólska ont formulé la même hypothèse à peu près en même temps[57]. En 2006, dans sa discussion des fonctions de la crête des lambeosaurinés, David Evans était favorable à cette hypothèse, du moins comme un facteur d'évolution de la crête[41].
Dans la culture populaire
Malgré sa rareté, Parasaurolophus est très représenté dans la culture populaire ; il est de ce fait l'Hadrosauridae le plus connu auprès du grand public.
Cinéma et télévision
- Dans le film Jurassic Park, on aperçoit un bref instant un groupe de Parasaurolophus, lors de la première apparition des Brachiosaurus[58].
- Dans le film Le Monde perdu : Jurassic Park, on en aperçoit pendant la chasse des braconniers.
- Dans Jurassic Park 3, des Parasaurolophus sont vus en compagnie de corythosaures.
- Dans Jurassic World, des Parasaurolophus peuvent être vus lorsque les enfants dans une gyrosphère visitent la vallée des herbivores.
- Dans Jurassic World : Fallen Kingdom, un parasaurolophus apparaît à la fin du film lorsque les dinosaures sont sur le point d’êtres libérés par Maisie.
- Dans la partie "Le sacre du printemps" du film Fantasia de Disney, des Parasaurolophus peuvent être vus, ils possèdent une voile à l'arrière de la tête.
- Dans le film Dinosaure de Disney, des Parasaurolophus peuvent être vus durant le film parmi les autres dinosaures.
- Dans la série des dessins animés « Le Petit Dinosaure » Becky et sa famille sont souvent confondus avec des Parasaurolophus (ils sont en fait des Saurolophus).
- Dans la série télévisée Dinotopia, et dans la série de romans éponymes de James Gurney, les Parasaurolophus sont utilisés comme montures par les humains[59].
- On peut voir un troupeau de Parasaurolophus dans l'épisode 6 de Prehistoric Park[60].
- Dans Hancock , Aaron donne un Parasaurolophus en plastique à Hancock quand il se retrouve en prison.
- Dans l'anime tiré du jeu Dinosaur King, Paris, le dinosaure de Zoé, est un Parasaurolophus.
Philatélie

La Roumanie a émis en 1994 un timbre à l'effigie du Parasaurolophus.
Numismatique
- La Monnaie Royale Canadienne a édité fin 2007, des pièces commémoratives de 4 dollars à l'effigie d'un Parasaurolophus[61].
Jeu vidéo
- Dans le jeu ParaWorld, les unités "Chevaucheurs gatling" sont des Parasaurolophus montés. On y trouve également des Parasaurolophus sauvages qui peuvent être chassés.
- Dans le jeu Turok de 2008, des parasaurolophus peuvent être rencontrés durant toute l'aventure, ils sont inoffensifs sauf s'ils sont provoqués.
- Dans le jeu Lost Eden, Dina et Mango, les deux alliés du héros, Adam, sont des Parasaurolophus.
- Dans le jeu Jurassic Park Operation Genesis, Parasaurolophus est considéré comme un « dinosaure star » et sa popularité est de trois étoiles.
- Dans le jeu ARK : Survival Evolved, il est possible d'apprivoiser un Parasaur (nom du dinosaure dans le jeu) et monter sur lui.
- Dans la saga Monster Hunter, l'Aptonoth, l'herbivore inoffensif qui se retrouve dans tous les jeux de la franchise, ressemble à Parasaurolophus.
- Dans l'application mobile "Jurassic Park Builder", il est possible d’élever un parasaurolophus.
- Dans le jeu Android "Jurassic World le jeu", il est possible de fusionner un parasaurolophus avec un Bonitasaura pour obtenir un hybride : le parasaura.
- Dans Jurassic World : Evolution et Jurassic World Evolution 2, Parasaurolophus fait partie du bestiaire du jeu.
- Dans Jurassic Park: The Game, Parasaurolophus est présent avec le statut « extra rare ».
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- (en) Richard Swann Lull et Nelda E. Wright, Hadrosaurian Dinosaurs of North America, vol. 40, Geological Society of America, coll. « Geological Society of America Special Paper », , 242 p. (OCLC 547322).

Publication originale
- (en) William Arthur Parks, « Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur », University of Toronto Studies, Geology Series, vol. 13, , p. 1-32.
Voir aussi
Liens externes
- Ressources relatives au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- (fr) Le Parasaurolophus sur DinoNews.net
Notes
Références taxonomiques
- (en) Référence Paleobiology Database : Parasaurolophus Parks, 1922
Références
- (en) Référence Paleobiology Database : †Parasaurolophus Parks 1922 (lambeosaurine) (consulté le )
- (en) Henry George Liddell et Robert Scott, A Greek-English Lexicon (Abridged Edition), United Kingdom, Oxford University Press, , 804 p. (ISBN 0-19-910207-4)
- (fr) « parasaurolophus », Dinonews
- (en) William Arthur Parks, « Parasaurolophus walkeri, a new genus and species of crested trachodont dinosaur », University of Toronto Studies, Geology Series, vol. 13, , p. 1-32
- « Tubicen », sur mediterranees.net
- (en) John R. Horner, David B. Weishampel (dir.), Catherine A Forster, Halszka Osmólska (dir.) et Peter Dodson (dir.), The Dinosauria, Berkeley, University of California Press, (réimpr. 2e), 438-463 p. (ISBN 0-520-24209-2), « Hadrosauridae »
- (en) « cyrto-, cyrt-, kyrto-, kyrt- (Greek: bent, curved). », sur wordsources.info
- (Lull, 1943)
- (Lull, 1943)
- (en) Michael K. Brett-Surman, Jonathan R. Wagner et Kenneth Carpenter (dir.), Horns and Beaks : Ceratopsian and Ornithopod Dinosaurs, Bloomington et Indianapolis, Indiana University Press, , 135-169 p. (ISBN 0-253-34817-X), « Appendicular anatomy in Campanian and Maastrichtian North American hadrosaurids »
- (fr) « Fiche du Parasaurolophus sur Dinomania »
- (en) Donald F. Glut, Dinosaurs : The Encyclopedia, Jefferson, Caroline du Nord, McFarland & Co, , 678-684 p. (ISBN 0-89950-917-7), « Parasaurolophus »
- (en) EnchantedLearning.com
- (en) Robert M Sullivan et Thomas E. Williamson, A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus, vol. 15, Albuqueque, Nouveau-Mexique, New Mexico Museum of Natural History and Science, coll. « New Mexico Museum of Natural History and Science Bulletin », , 1-52 p.
- (en) Ostrom John H., « A new species of hadrosaurian dinosaur from the Cretaceous of New Mexico », Journal of Paleontology, vol. 35, no 3, , p. 575-577 (résumé)
- Fiche du Parasaurolophus sur Dinosoria
- (en) Charles Whitney Gilmore, « On the genus Stephanosaurus, with a description of the type specimen of Lambeosaurus lambei, Parks », Canada Department of Mines Geological Survey Bulletin, vol. 38, no 43, , p. 29-48
- (en) David B. Weishampel (dir.), Jack R. Horner, Halszka Osmólska (dir.) et Peter Dodson (dir.), The Dinosauria, Berkeley, University of California Press, (réimpr. Ire), 534-561 p. (ISBN 0-520-06727-4), « Hadrosauridae »
- (en) Evans David C. et Reisz, Robert R., « Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta », Journal of Vertebrate Paleontology, vol. 27, no 2, , p. 373-393 (résumé)
- (en) Godefroit Pascal ; Shuqin Zan; et Liyong Jin, « Charonosaurus jiayinensis n. g., n. sp., a lambeosaurine dinosaur from the Late Maastrichtian of northeastern China », Comptes rendus de l'Academie des Sciences, Paris, Sciences de la Terre et des planètes, vol. 330, , p. 875-882 (DOI 10.1016/S1251-8050(00)00214-7, résumé)
- (en) Albert Prieto-Márquez, Fabio M. Dalla Vecchia, Rodrigo Gaete et Àngel Galobart, « Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis », PLoS ONE, vol. 8, no 7, , e69835 (DOI 10.1371/journal.pone.0069835)
- (en) A. Prieto-Marquez, G.M. Erickson et J.A. Ebersole, « A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs », Journal of Vertebrate Paleontology, vol. 36, no 2, , e1054495 (DOI 10.1080/02724634.2015.1054495)
- (en) Thomas E. Williamson, S.G. Lucas (dir.) et A.B. Heckert (dir.), Dinosaurs of New Mexico, vol. 17, Albuquerque, New Mexico Museum of Natural History and Science, coll. « New Mexico Museum of Natural History and Science Bulletin », , 191-213 p., « Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico »
- (en) Eberth, D.A. 2005. The geology. In: Currie, P.J., and Koppelhus, E.B. (eds), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington and Indianapolis,p.54-82. (ISBN 0-253-34595-2).
- (en) D.C. Evans, R. Bavington et N.E. Campione, « An unusual hadrosaurid braincase from the Dinosaur Park Formation and the biostratigraphy of Parasaurolophus (Ornithischia: Lambeosaurinae) from southern Alberta », Canadian Journal of Earth Sciences, vol. 46, no 11, , p. 791–800 (DOI 10.1139/E09-050, lire en ligne)
- (en) Michael J. Ryan, David C. Evans, Phillip J. Currie (dir.) et Eva Koppelhus (dir.), A Spectacular Ancient Ecosystem Revealed [« Dinosaur Provincial Park »], Bloomington, Indiana University Press, , 312-348 p. (ISBN 0-253-34595-2), « Ornithischian Dinosaurs »
- (en) Evans David C.; Reisz, Robert R.; Dupuis, Kevin, « A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus », Journal of Vertebrate Paleontology, vol. 27, no 3, , p. 642–650 (DOI 10.1671/0272-4634(2007)27[642:AJPOHB]2.0.CO;2)
- (en) Phillip J Currie et Dale A. Russell, Geographic and stratigraphic distribution of dinosaur remains [« Dinosaur Provincial Park »], 553 p.
- (en) Sullivan, R.M., and Lucas, S.G. 2006. "." New Mexico Museum of Natural History and Science, Bulletin 35: p. 7-29
- (de) Carl Wiman, « Parasaurolophus tubicen, n. sp. aus der Kreide in New Mexico », Nova Acta Regia Societas Scientarum Upsaliensis, series 4, vol. 7, no 5, , p. 1–11
- (en) David B. Weishampel, « Parasaurolophus (Reptilia: Hadrosauridae) from Utah », Journal of Paleontology, vol. 53, no 6, , p. 1422–1427
- (en) Alan L. Titus, Mark A. Loewen, 2013, « The Kaiparowits Formation: A Remarkable Record of Late Cretaceous Terrestrial Environments, Ecosystems, and Evolution in Western North America », At the Top of the Grand Staircase, The Late Cretaceous of Southern Utah Edited by Alan L. Titus and Mark A. Loewen.
- (en) A.A. Farke, D.J. Chok, A. Herrero, B. Scolieri, S. Werning et John Hutchinson (dir.), « Ontogeny in the tube-crested dinosaur Parasaurolophus (Hadrosauridae) and heterochrony in hadrosaurids », PeerJ, vol. 1, , e182 (PMID 24167777, PMCID 3807589, DOI 10.7717/peerj.182, lire en ligne)
- Joël Ignasse, « Le fossile complet d'un bébé dinosaure », sur Sciences et Avenir, (consulté le ).
- (en) Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loeuff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth, M.P.; and Noto, Christopher R. (2004). "Dinosaur Distribution", in The Dinosauria (2nd), pp. 517–606.
- (en) Eberth, David A. 2005. "The geology", in Dinosaur Provincial Park, pp. 54-82
- (en) Braman, Dennis R., and Koppelhus, Eva B. 2005. "Campanian palynomorphs", in Dinosaur Provincial Park, pp. 101-130.
- (en) Dale A. Russell, An Odyssey in Time : Dinosaurs of North America, Minocqua, Wisconsin, NorthWord Press, (ISBN 1-55971-038-1), p. 160-164
- (en) Robert Bakker, New Theories Unlocking the Mystery of the Dinosaurs and their Extinction [« The Dinosaur Heresies »], New York, William Morrow, , 194 p. (ISBN 0-8217-2859-8)
- Norman David B., An Original and Compelling Insight into Life in the Dinosaur Kingdom [« The Illustrated Encyclopedia of Dinosaurs »], New York, Crescent Books, , 122-127 p. (ISBN 0-517-46890-5), « Hadrosaurids II »
- (en) Evans David C., « Nasal cavity homologies and cranial crest function in lambeosauriné dinosaurs », Paleobiology, vol. 32, no 1, , p. 109–125 (résumé)
- (en) James Hopson, « The evolution of cranial display structures in hadrosaurian dinosaurs », Paleobiology, vol. 1, no 1, , p. 21–43
- (en) Alfred Sherwood Romer, Vertebrate Paleontology, University of Chicago Press, , 491 p. (OCLC 1186563)
- (en) Wilfarth Martin, « Russeltragende Dinosaurier », Orion (Munich), vol. 2, , p. 525–532
- Charles Mortram Sternberg, « Hooded hadrosaurs of the Belly River Series of the Upper Cretaceous », Canada Department of Mines Bulletin, vol. 77, no 52, , p. 1–37
- Colbert Edwin H., The Ruling Reptiles and their Relatives [« The Dinosaur Book »], vol. 14, New York, American Museum of Natural History, coll. « Man and Nature Publications », , 156 p. (OCLC 691246)
- (de) Othenio Abel, « Die neuen Dinosaurierfunde in der Oberkreide Canadas », Jarbuch Naturwissenschaften, vol. 12, no 36, , p. 709–716 (DOI 10.1007/BF01504818)
- (en) Halszka Osmólska, « Nasal salt glands in dinosaurs », Acta Palaeontologica Polonica, vol. 24, , p. 205–215
- John Ostrom, « The cranial crests of hadrosaurian dinosaurs », Postilla, vol. 62, , p. 1–29
- Gish Duane T., Dinosaurs by Design, Green Forest, Master Books, , 82 p. (ISBN 0-89051-165-9)
- (en) David B. Weishampel, « Acoustic analyses of potential vocalization in lambeosauriné dinosaurs (Reptilia:Ornithischia) », Paleobiology, vol. 7, no 2, , p. 252–261
- (en) Weishampel David B., « The nasal cavity of lambeosauriné hadrosaurids (Reptilia:Ornithischia): comparative anatomy and homologies », Journal of Paleontology, vol. 55, no 5, , p. 1046–1057
- « Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur », Sandia National Laboratories, (consulté le )
- (en) Diegert Carl F. et Williamson, Thomas E., « A digital acoustic model of the lambeosaurinae hadrosaur Parasaurolophus tubicen », Journal of Vertebrate Paleontology, vol. 18, no 3, Suppl., , p. 38A
- (en) Sullivan Robert M. et Williamson, Thomas E., « A new skull of Parasaurolophus (long-crested form) from New Mexico: external and internal (CT scans) features and their functional implications », Journal of Vertebrate Paleontology, vol. 16, no 3, Suppl., , p. 68A
- (en) Wheeler P.E., « Elaborate CNS cooling structure in large dinosaurs », Nature, vol. 275, , p. 441–443 (DOI 10.1038/275441a0)
- (en) Teresa Maryańska, « Aspects of hadrosaurian cranial anatomy », Lethaia, vol. 12, , p. 265–273 (DOI 10.1111/j.1502-3931.1979.tb01006.x)
- « Les Dinosaures de Jurassic Park », sur DinoNews
- [image] « Photo issue de la série télévisée »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?)
- « (en)description de l'épisode 6 de Prehistoric Park »
- (fr) « Parasaurolophus », Monnaie Royale Canadienne
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}