Canardia
Canardia garonnensis
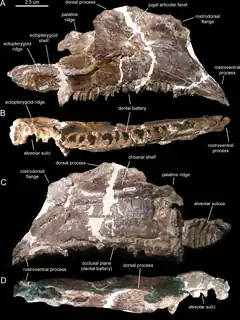
Canardia est un genre éteint de dinosaures hadrosauridés, classé dans la sous-famille des Lambeosaurinae. Ses restes fossiles ont été retrouvés près du village de Marignac-Laspeyres dans le département de la Haute-Garonne, en région Occitanie (France). Ils proviennent de la Formation des Marnes d’Auzas, datée du Maastrichtien terminal (Crétacé supérieur)[1], il y a entre 67,5 et 66 millions d'années, juste avant l'extinction massive de la fin du Crétacé il y a 66 millions d'années[2]. L'espèce type et seule espèce connue, Canardia garonnensis, a été décrite et nommée en 2013 par Albert Prieto-Márquez (d), Fabio M. Dalla Vecchia, Rodrigo Gaete et Àngel Galobart[2]. Elle n’est connue que par des spécimens juvéniles. Le nom du genre vient du français « canard », une allusion au fait que cet animal appartient aux hadrosauridés, également appelés « dinosaures à bec de canard ». L'épithète spécifique garonnensis fait référence au département de la Haute-Garonne où ce dinosaure a été trouvé. Bien qu'universellement reconnu comme un lambeosauriné, sa position précise au sein de ceux-ci est débattue. Certains auteurs le considèrent comme un proche parent du genre Aralosaurus d’Asie centrale avec lequel il formerait la tribu des Aralosaurini[2], alors que d’autres l’incluent dans un clade plus dérivé, les Arenysaurini dans lequel sont placés tous les lambéosaurinés d’Europe et d’Afrique du Nord[3]. Canardia fut l'un des derniers dinosaures non aviens et a vécu dans ce qui était à l'époque l'île Ibéro-Armoricaine, une terre émergée qui comprenait une grande partie de la France et de l'Espagne actuelle[2] - [4].
Découverte

Canardia est connue par divers restes crâniens et postcraniens trouvés dans plusieurs sites du sud-ouest de la France[2]. La localité type de Tricouté 3, près du village de Marignac-Laspeyres, a été découverte en 1999 par une équipe du Musée des dinosaures d’Espéraza[1]. Ce site est situé dans la partie basale de la Formation des Marnes d’Auzas, laquelle correspond à un environnement laguno-continental[1] - [2]. Les restes de cet hadrosaure étaient conservés dans un petit niveau marno-gréseux lenticulaire intercalé dans un épais banc de grès[1] - [2]. Ils appartiennent à au moins deux individus différents, comme l'indique la présence de deux scapulas gauches parmi les échantillons. Les éléments crâniens comprennent un maxillaire droit presque complet (l'holotype), un maxillaire gauche partiel, un préfrontal droit, un postorbitaire droit incomplet, un carré gauche, un ptérygoïde droit, un surangulaire gauche partiel, un articulaire droit, une couronne isolée d’une dent du dentaire, et une portion de batterie dentaire de la mandibule. Le matériel post-crânien est représenté par deux scapulas gauches partielles, une plaque sternale gauche incomplète, un humérus gauche, et un pubis droit partiel. En 2003, ces spécimens ont été provisoirement attribués à Pararhabdodon sp., un genre espagnol qui était à l'époque le seul lambéosauriné connu en Europe. Ce n'est qu'en 2013 que Prieto-Márquez et des collègues on reconnu ces os comme ceux d'un nouveau genre. Tous les os sont de tailles relativement petites (16,9 cm pour le maxillaire holotypique, 21,8 cm pour la scapula la plus complète, et 19,4 cm pour l’humérus) et appartiennent à des individus immatures[1] - [2].
Les mêmes auteurs ont également attribué à Canardia un maxillaire associé à un carré trouvés dans les dépôts marins de la Formation des Marno-calcaires de Gensac dans la carrière de Larcan, à une vingtaine de kilomètres à l’ouest de Marignac-Laspeyres[2]. Les deux os, décrits pour la première fois en 2010 par Bilotte et des collègues, étaient situés à environ 1 mètre sous un niveau d'iridium marquant la limite Crétacé-Paléogène[5]. Ce niveau est l’équivalent latéral de la partie la plus haute de la Formation des Marnes d’Auzas. L’individu de Larcan est ainsi géologiquement plus jeune que les spécimens de Marignac-Laspeyres. Cette découverte indique également que Canardia fut l'un des derniers dinosaures non aviens à exister en Europe juste avant l'événement d'extinction Crétacé-Paléogène[5] - [2]. Le spécimen de Larcan appartient à un individu juvénile (le maxillaire mesure seulement 11,5 cm), dont les restes ont été transportés par une rivière jusqu'à la mer[5].
Description

Canardia est principalement caractérisé par son maxillaire montrant une région rostrodorsale élargie qui forme un bord subrectangulaire proéminent s’élevant verticalement au-dessus du processus rostroventral. Ce caractère est également connu chez Aralosaurus tuberiferus du Kazakhstan, qui vécu 20 millions d'années avant Canardia. Cependant, le maxillaire de Canardia diffère de celui d’Aralosaurus par son plateau ectoptérygoïde subhorizontal (il est parallèle au segment postérieur de la rangée dentaire) alors qu’il est clairement incliné chez Aralosaurus. Le maxillaire de Canardia porte au moins 26 rangées de dents[2] contre 30 chez Aralosaurus[6], mais la totalité de la rangée dentaire n'est pas préservée chez Canardia. Les dents maxillaires ont une carène médiane alors que les dents dentaires présentent une crête accessoire devant la carène sub-centrale, comme c’est le cas chez Aralosaurus mais aussi chez beaucoup d’autres hadrosaures[2] - [6]. Canardia se distingue également d’Aralosaurus par la forme de son préfrontal en vue dorsale. Chez Canardia, le processus rostroventral du préfrontal est rostrocaudalement étroit, alors que chez Aralosaurus, le même processus osseux est large et bien exposé latéralement. De plus, le préfrontal de Canardia présente une saillie osseuse dorsomédiale qui est absente chez Aralosaurus[2].
Classification
La classification précise de Canardia parmi les lambéosaurinés est encore incertaine. En ce basant sur les similitudes entre les maxillaires de Canardia et d’Aralosaurus, Prieto-Márquez et des collègues avaient en 2013 réunis ces deux genres dans un nouveau groupe de lambeosaurinés basaux appelé Aralosaurini[2] - [7]. Cependant, une classification différente a été proposée en 2021 par Longrich et des collègues lors de la description du genre Ajnabia du Maroc. Contrairement aux travaux précédents qui répartissaient les lambéosaurinés européens dans différentes lignées (Lambeosaurini, Parasaurolophini, Tsintaosaurini, et Aralosaurini), l’étude de Longrich et al. considère que tous les lambéosaurinés européens forment un clade monophylétique nommé Arenysaurini, dont fait également partie le genre nord-africain nouvellement décrit. Canardia n’est pas ici identifié comme étant particulièrement proche d’Aralosaurus, et occupe une position plus dérivée[3].
Les analyses phylogénétiques effectuées par Prieto-Márquez et ses collègues ont conduit au cladogramme suivant[2] :
Ci-dessous le cladogramme des lambéosaurinés publié par Longrich et al.[3].
| Hadrosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Crête
On ne sait pas si Canardia avait une crête osseuse creuse sur le crâne, comme la plupart des lambeosaurinés. Bien qu'aucun crâne complet ne soit connu pour les deux groupes d’hadrosaures auxquels il a été successivement rattaché, les deux ont une conformation crânienne très différente. Aralosaurus, l’un des lambéosaurinés les plus primitifs, n’avait pas de crête sur le toit crânien car ce dernier n’était pas modifié pour recevoir une telle structure. A la place, il possédait une structure creuse devant les orbites[8]. Cependant, comme seul un fragment de cette structure a été préservé, sa taille et sa forme sont malheureusement inconnues. En revanche, la morphologie du toit crânien d’Arenysaurus montre clairement qu’il supportait une crête osseuse dont la forme est toutefois encore inconnue[9].
Paléobiogéographie
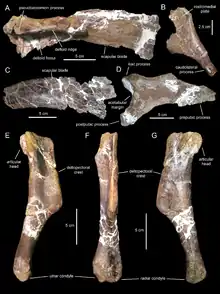
Les paléontologues s’accordent sur une probable origine asiatique des lambéosaurinés européens. Selon Prieto-Marquez et des collègues, Canardia et le genre espagnol Pararhabdodon avaient respectivement comme plus proche parent les genres asiatiques Aralosaurus et Tsintaosaurus. Ces derniers sont respectivement 20 et 15 millions d’années plus anciens que leurs cousins européens présumés. Ainsi, Canardia et Pararhabdodon appartiendraient tous deux à des groupes de lambéosaurinés plutôt archaïques pour leur époque et l'île Ibéro-Armoricaine aurait constituée un ultime refuge pour ces groupes anciens d’hadrosaures lambéosaurinés[2] - [4]. Longrich et des collègues, lesquels placent Canardia et tous les autres lambéosaurinés européens dans le clade des Arenysaurini, suggèrent également une origine asiatique pour des raisons paléogéographiques. Comme on ne connaissait pas de lambeosaurinés dans les riches gisements du Campanien et du Maastrichtien inférieur de l’île Ibéro-Armoricaine, il a été suggéré que ce groupe avait dû atteindre cette île assez tard, peut-être à la fin du Maastrichtien inférieur ou au cours du Maastrichtien supérieur[2]. La découverte ultérieure en Espagne de nombreux restes de lambéosaurinés dans les niveaux les plus élevés du Maastrichtien inférieur plaide en faveur de la première hypothèse[10] - [11]. Cette migration a probablement été réalisée en plusieurs temps, d’abord par des liaisons terrestres temporaires entre la masse continentale de l’Asie de l’ouest et les îles orientales de l’archipel européen, puis entre les différentes îles européennes jusqu’à l’île Ibéro-Armoricaine[4]. De cette île, les lambeosaurinés auraient ensuite atteints l’Afrique du Nord par dispersion océanique, les hadrosaures nageant ou dérivant sur des radeaux naturels (dans le cas d’hadrosaures nouveau-nés) sur de grandes distances, l’île Ibéro-Armoricaine et les côtes du Maroc (où vivait Ajnabia) étant séparés à l’époque par 500 km de pleine mer[3].
Paléoécologie
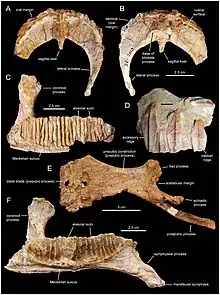
La Formation des Marnes d’Auzas a une épaisseur de 100 m. Elle correspond à des sédiments dont l'environnement de dépôt a évolué au cours du temps, passant du domaine paralique (lagunes, marais littoraux, chenaux de marée) à la base de la formation, vers un milieu plus continental (plaine alluviale, chenaux fluviatiles) dans sa partie supérieure[1]. D’un point de vue paléogéographique, les sites de la Formation des Marnes d’Auzas étaient situés sur la côte ouest de l’île Ibéro-Armoricaine, face au golfe Atlantique[4].
Peu de fossiles de vertébrés ont été trouvés avec les restes de Canardia. La localité type de Tricouté 3 , dans la partie basale de la Formation des Marnes d’Auzas, n’a produit qu’une dent de crocodilien[1], alors que les dépôts marins de la Formation des Marno-calcaires de Gensac (l’équivalent latéral du sommet des Marnes d'Auzas) n'a pas encore livré d'autres restes de vertébrés terrestres[5]. Cependant, une faune plus diversifiée est connue dans les gisements de Cassagnau, dans la partie centrale des Marnes d’Auzas[12] - [1]. La faune de Cassagnau, également située près de Marignac-Laspeyres, comprend plusieurs poissons (Lepisosteidae, Phyllodontidae, Sparidae)[12] - [1], des amphibiens (Albanerpetontidae)[12] - [1], plusieurs tortues dont une carapace complète d’Iberoccitanemys convenarum[12] - [1] - [13], divers crocodiliens représentés notamment par des dents de forme semblable à celle de Musturzabalsuchus et par un crâne complet attribué à Thoracosaurus neocesariensis[12] - [1] - [14], deux squamates (un lézard indéterminé, et un varanoïde[12] - [1] qui est probablement un mosasauroïde d’eau douce[15]), un probable oiseau énantiornithe[12] - [1], un dromaeosauridé indéterminé (connu seulement par des dents)[12] - [1], un théropode indéterminé de plus grande taille (uniquement connu par une dent caractérisée par sa carène antérieure s'étendant sur le côté lingual de la dent)[1], un titanosaure indéterminé représenté par des dents et un métacarpien[12] - [1] (la mophologie des dents suggère qu'elles appartiennent à une forme semblable à Atsinganosaurus)[16], et un hadrosaure lambéosauriné indéterminé. Ce dernier est exclusivement représenté par des individus juvéniles et comprend un squelette post-crânien articulé, des dents, et deux dentaires gauches, dont l’un était associé à un prédentaire[12] - [1] - [2]. Aucun maxillaire n’est préservé sur ces spécimens, il est donc impossible de savoir si cet hadrosaure appartient à Canardia ou à un autre lambéosauriné. La Formation des Marnes d’Auzas est également présente dans le département de l’Ariège, notamment près du village de Mérigon. Là, un lit de grès grossier au sommet de la formation a donné une dent et un fragment de dentaire d'un hadrosaure indéterminé, ainsi qu'une vertèbre cervicale d'un ptérosaure géant de 9 mètres d'envergure de la famille des Azhdarchidae[17] - [1].
Notes et références
Références
- Y. Laurent, « Les faunes de vertébrés continentaux du Maastrichtien supérieur d’Europe : systématique et biodiversité », Strata, vol. 41, , p. 1-81
- (en) Albert Prieto-Márquez, Fabio M. Dalla Vecchia, Rodrigo Gaete et Àngel Galobart, « Diversity, Relationships, and Biogeography of the Lambeosaurine Dinosaurs from the European Archipelago, with Description of the New Aralosaurin Canardia garonnensis », PLoS ONE, vol. 8, no 7, , e69835 (DOI 10.1371/journal.pone.0069835)
- (en) N.R. Longrich, X. Pereda Suberbiola, R.A. Pyron et N-E. Jalil, « The first duckbill dinosaur (Hadrosauridae : Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography », Cretaceous Research, vol. 120, , p. 104678 (DOI 10.1016/j.cretres.2020.104678)
- (en) Z. Csiki-Sava, E. Buffetaut, A. Ősi, X. Pereda-Suberbiola et S.L. Brusatte, « Island life in the Cretaceous-faunal composition, biostratigraphy, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago », ZooKeys, vol. 469, , p. 1-161 (DOI 10.3897/zookeys.469.8439, lire en ligne)
- M. Bilotte, Y. Laurent et D. Teodori, « Restes d’Hadrosaure dans le Crétacé terminal marin de Larcan (Petites Pyrénées, Haute-Garonne, France) », Carnets de Géologie / Notebooks on Geology - Note brève / Letter, vol. 2010/02, , p. 1-10 (DOI 10.4267/2042/32428)
- (en) A.K. Rozhdestvensky, « Gadrozavry Kazakhstana [Hadrosaurs of Kazakhstan]. [Upper Paleozoic and Mesozoic Amphibians and Reptiles] », Akademia Nauk SSSR, Moscow, , p. 97-141 (lire en ligne)
- (en) A. Prieto-Marquez, G.M. Erickson et J.A. Ebersole, « A primitive hadrosaurid from southeastern North America and the origin and early evolution of 'duck-billed' dinosaurs », Journal of Vertebrate Paleontology, vol. 36, no 2, , e1054495 (DOI 10.1080/02724634.2015.1054495)
- (en) P. Godefroit, V. Alifanov et Y. Boltsky, « A re-appraisal of Aralosaurus tuberiferus (Dinosauria, Hadrosauridae) from the Late Cretaceous of Kazakhstan », Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, vol. 74, , p. 139-154 (lire en ligne)
- (en) Xabier Pereda-Suberbiola, José Ignacio Canudo, Penélope Cruzado-Caballero, José Luis Barco, Nieves López-Martínez, Oriol Oms et José Ignacio Ruiz-Omeñaca, « The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the Uppermost Cretaceous of Aren (Huesca, Spain) », Comptes Rendus Palevol, vol. 8, no 6, , p. 559–572 (DOI 10.1016/j.crpv.2009.05.002)
- (en) V. Fondevilla, F.M. Dalla Vecchia, R. Gaete, À. Galobart, B. Moncunill-Solé et M. Köhler, « Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) remains from Basturs Poble bonebed (late early Maastrichtian, tremp Syncline, Spain) », PLos ONE, vol. 13(10): e0206287, , p. 1-33 (DOI 10.1371/journal.pone.0206287)
- (en) A. Prieto-Márquez, V. Fondevilla, A.G. Sellés, J.R. Wagner et À. Galobart, « Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European Archipelago », Cretaceous Research, vol. 96, , p. 19-37 (DOI 10.1016/j.cretres.2018.12.002)
- (en) Y. Laurent, M. Bilotte et J. Le Loeuff, « Late Maastrichtian continental vertebrates from southwestern France: correlation with marine fauna », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 187, , p. 121-135 (DOI 10.1016/S0031-0182(02)00512-6)
- (en) A. Pérez-García, F. Ortega et X. Murelaga, « A new genus of Bothremydidae (Chelonii, Pleurodira) in the Cretaceous of southwestern Europe », Geobios, vol. 45(2), , p. 219-229 (DOI 10.1016/j.geobios.2011.03.001)
- Y. Laurent, E. Buffetaut et J. Le Loeuff, « Un crâne de Thoracosaurine (Crocodylia, Crocodylidae) dans le Maastrichtien supérieur du sud de la France », ORYCTOS, vol. 3, , p. 19-27
- (en) G. Garcia, N. Bardet, A. Houssaye, X. Pereda-Suberbiola et X. Valentin, « Mosasauroid (Squamata) discovery in the Late Cretaceous (Early Campanian) continental deposits of Villeveyrac-L’Olivet, Southern France – Découverte de Mosasauroidea (Squamata) dans le Crétacé supérieur (Campanien inférieur) continental de Villeveyrac – L’Olivet, sud de la France », Comptes Rendus Palevol, vol. 14(6-7), , p. 495-505 (DOI 10.1016/j.crpv.2015.05.002)
- (en) G. Garcia, S. Amicot, F. Fournier, E. Thouand et X. Valentin, « A new Titanosaur genus (Dinosauria, Sauropoda) from the Late Cretaceous of southern France and its paleobiogeographic implications », Bulletin de la Société Géologique de France, vol. 181(3), , p. 269-277 (DOI 10.2113/gssgfbull.181.3.269)
- (en) E. Buffetaut, Y. Laurent, J. Le Loeuff et M. Bilotte, « A terminal Cretaceous giant pterosaur from the French Pyrenees », Geological Magazine, vol. 134(4), , p. 553-556 (DOI 10.1017/S0016756897007449)
Voir aussi
Références taxinomiques
- (en) Référence Paleobiology Database : Canardia Prieto-Marquez et al., 2013