Gisement de fossiles du Geiseltal
Le gisement de fossiles du Geiseltal se trouve dans la région où l'on exploitait du lignite au sud de la ville de Halle en Saxe-Anhalt. Il représente un gisement important de plantes et d'animaux morts, de l'époque de l'Éocène moyen (il y a 48 à 41 millions d'années). On a exploité au Geiseltal depuis 1698 du lignite à ciel ouvert, mais les premiers fossiles n'ont été remarqués qu'au début du XXe siècle plutôt par hasard. Les fouilles scientifiques planifiées ont commencé en 1925, animées par l'Université Martin-Luther de Halle-Wittenberg. Les recherches, interrompues par la seconde Guerre mondiale, peuvent être subdivisées en deux phases. En raison de la fin progressive des couches de lignite, les fouilles ont commencé à ralentir à partir du milieu des années 1980, et ont été définitivement terminées au début du troisième millénaire.

| Gisement de fossiles du Geiseltal | |||
 Vue du sud sur le Geiseltal est, au fond le terril de Klobikau. | |||
| Localisation | |||
|---|---|---|---|
| Coordonnées | 51° 18′ 28″ nord, 11° 52′ 09″ est | ||
| Pays | |||
| au sud de Halle | |||
| Informations géologiques | |||
| Période | Éocène moyen | ||
| Âge | 48–41 Ma | ||
| Géolocalisation sur la carte : Saxe-Anhalt
Géolocalisation sur la carte : Allemagne
| |||
Le lignite du Geiseltal est subdivisé en quatre filons principaux, les fossiles étant principalement contenus dans les trois inférieurs. Le centre de la distribution des fossiles est vers les parties méridionale et centrale du gisement. Les trouvailles comportent des restes de plantes et d'animaux. Une particularité est la découverte de feuilles, fruits et tiges presque complets, et aussi des squelettes de vertébrés et des restes d'insectes. Parmi les trouvailles les plus connues sont un squelette complet d'ancêtre du cheval Propalaeotherium, trouvé en 1933. De plus, on compte des vertébrés tels que des artiodactyles, des petits mammifères comme des insectivores et des chauves-souris, ainsi que des oiseaux, des crocodiles, des tortues, des serpents, des amphibiens et des poissons. Pour de nombreux vertébrés, on a aussi trouvé des restes des tissus mous. En tout, on connaît 80 sites de fouille qui ont fourni plus de 30 000 échantillons, dont 36 avec une bonne proportion de vertébrés. En raison de la bonne préservation des fossiles, et de leur variété, le Geiseltal est considéré comme un lagerstätte de conservation et de concentration.
Les spécificités des fossiles du Geiseltal, et avant tout des vertébrés, et spécialement des mammifères, résident dans leur conservation unique dans le lignite, ce qui n'a pas lieu ailleurs en Europe. Pour le développement des mammifères, le Geiseltal représente un gisement important, car on peut y observer le développement de divers groupes sur un espace de quelques millions d'années. Cette faune mammalienne est une référence stratigraphique pour l'intervalle de 47 à 43 millions d'années. Les trouvailles multiples du monde animal et végétal, ainsi que les nombreuses données géologiques permettent une reconstruction du paysage très précise. Dans ce cadre, à l'époque de la formation du lignite, il y avait une forêt à plusieurs étages à proximité de la côte, traversée par des ruisseaux, des étangs et des marais. La forêt était soumise à l'influence d'un climat subtropical humide, et hébergeait un monde animal diversifié.
Situation géographique
Le Geiseltal (vallée de la Geisel, petit affluent de la Saale) se trouve environ à 20 km au sud de Halle et 10 km au sud-ouest de Mersebourg en Saxe-Anhalt. Elle s'étend sur une longueur de 15 km du O-NO vers le E-SE, et est large de 0,5 à 5 km. Au nord, elle est limitée par le plateau de grès bigarré de Mersebourg, et au sud par le plateau de calcaire coquillier de Mücheln dans la dépression de Querfurt-Freyburg. Le Geiseltal est le lit de la rivière éponyme Geisel, qui sort de l'une des plus grandes sources jaillissantes d'Allemagne centrale à St Micheln près de Mücheln, puis se déverse au bout de 19 km, par les étangs de Gotthardt dans la Saale. Son bassin versant est d'environ 35 km2[1].
À l'origine, le territoire du Geiseltal était relativement plat, ce qui s'explique par les grès bigarrés qui s'étendent à l'ouest de la Saale. En général, les altitudes de la partie est se trouvaient vers 100 mètres, en montant à 150 mètres et plus vers l'ouest. Mais le paysage a été notablement changé par l'activité intensive d'exploitation des mines de lignite à ciel ouvert, notamment dans les 150 ans derniers, ce qui a conduit à la destruction de nombreux villages, et aussi par la déviation sur un domaine d'environ 90 kilomètres carrés à de nombreux endroits du cours de la Geisel vers le sud. À la suite de la renaturation des mines abandonnées, des lacs assez grands ont vu le jour, et à présent, ils déterminent l'ensemble du Geiseltal. Dès la seconde moitié du XXe siècle, les lacs de Südfeld (champ sud) et de Runstedt ont été créés dans les parties est et sud-est, tandis que dans la partie ouest, un des plus grands lacs artificiels d'Europe centrale, le Geiseltalsee a pu être complètement rempli en 2011. Aujourd'hui, les parties est et ouest du Geiseltal sont séparées par un barrage en remblais, d'une hauteur jusque 140 m, qui supporte, outre le passage du cours actuel de la Geisel, d'importantes voies de transport comme des routes et des voies ferrées[2].
Géologie
Depuis le début du XXe siècle, la géologie du Geiseltal a été bien explorée avec des forages profonds pour les recherches de situation des gisements de lignite. Dans certains domaines, ces forages étaient effectués jusqu'à 100 m de l'un à l'autre. Largement, le sous-sol est constitué de dépôts des permiens inférieur (Cisuralien), moyen (Guadalupien) et supérieur (Lopingien), datant d'entre 300 et 240 millions d'années. Là-dessus, il y a des couches sédimentaires de grès bigarré (datant d'entre 251 et 243 millions d'années), pour la plupart des couches inférieures et moyennes, puis au sud des restes des couches supérieures, surmontées par des dépôts de calcaire coquillier sur les bords de la vallée. Les couches supérieures comprennent le Keuper et le Crétacé d'une période d'environ 140 millions d'années[3] - [4].
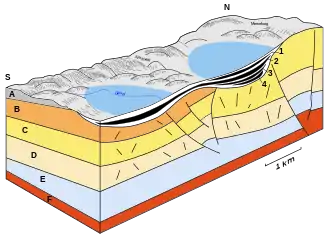
Le Geiseltal actuellement s'articule en plusieurs entités. Le seuil principal de Neumark, une élévation du grès bigarré, la partage en deux parties en gros égales, occidentale et orientale. Des bassins accessoires ou subordonnés sont le bassin d'Elise, d'Elisabeth, de Wernsdorf et autres, les seuils pré-tertiaires sont le seuil de Kayna et le seuil est[5]. Géologiquement, le Geiseltal représente une structure d'effondrement, dont l'origine n'est pas entièrement comprise, et qui a été le résultat de plusieurs processus. Il est possible que la dissolution du sel du schiste cuivreux pendant le Paléogène, reliée avec l'érosion chimique du calcaire coquillier (karst) a conduit à des pertes de masse en sous-sol, ce qui a abouti à un effondrement des couches supérieures et à la formation de bassins. En particulier, la dissolution des sels s'est conjuguée avec les forces tectoniques en sous-sol, résultant de la tectonique des plaques, qui ont aussi suscité la surrection des Alpes à la limite Crétacé-Tertiaire, il y a environ 65 millions d'années. Ceci provoque une élévation de la pression sur la couche de schiste cuivreux initialement plane, qui s'écoule alors de côté. Du côté nord du Geiseltal, on a détecté par des mesures sismiques des perturbations tectoniques, ce que l'on appelle la perturbation du bord nord du Geiseltal, qui parfois atteint une différence de niveau jusqu'à 200 mètres dans les couches supérieures (grès bigarré)[3] - [4] - [6].
Dépôts paléogènes
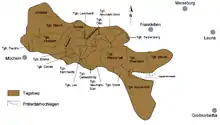
L'ensemble des dépôts paléogènes atteignent dans le Geiseltal une puissance allant jusqu'à 200 m, mais se limitent largement à l'Éocène (entre 56 et 34 millions d'années). Ces sédiments tertiaires ont été commencés par des argiles et des limons, et en partie des éléments plus grossiers. La puissance du lignite principalement en couches minces, voire en feuilles, se situe entre 30 et 80 m, mais dépasse par endroits 120 m[7]. En général, le lignite du Geiseltal est réparti en quatre filons principaux : inférieur, moyen (séparé en moyen inférieur et moyen supérieur) et supérieur, mais il présente aussi localement au nord un filon de base. Tous les filons sont séparés en sous-filons. Individuellement, les filons atteignent une puissance de 10 à 60 m. Ils sont séparés par des sédiments de limon sableux, qui représentent des horizons-guides et servent à la délimitation des filons. Dans le Geiseltal sud, dans la mine Cecilie, on a observé néanmoins un passage continu des filons moyen au supérieur. L'étendue des divers filons de lignite du nord vers le sud est marquée différemment, et reliée à l'effondrement à la suite des dissolutions et érosion souterraine, mais elle se continue en général avec l'altitude croissante des filons vers le sud. Seul le filon supérieur couvre la surface dans le Geiseltal. La collection de fossiles très riche de l'Éocène provient surtout des filons inférieur et moyen[3] - [8] - [9].
Couches couvrantes
La couverture consiste dans le Geiseltal en dépôts de 20 à 50 mètres d'épaisseur du Pléistocène (de 2,5 millions d'années à 10 000 ans), cependant les plus anciens ont été déplacés par les formations glaciaires ultérieures. Les dépôts contiennent à côté des pierrailles fluviales étalées sur de larges surfaces surtout des moraines de fond des âges glaciaires de l'Elster et de la Saale, les premières seulement au bord sud-est, les secondes sur toute la surface. L'ensemble des couches est recouvert par plusieurs mètres de lœss de la dernière période glaciaire (glaciation de Weichsel), qui supporte le sol de terre noire. Entre les moraines de la période de la Saale et les lœss du Weichsélien, on trouve plusieurs structures en bassin, surtout au nord-est, dues à nouveau à des effondrements, cette fois des couches supérieures du lignite. Ces couches ont été pendant les périodes périglaciaires des phases froides saturées d'un excès d'eau dans leur partie supérieure, et en raison de leur densité rendue ainsi faible, se sont écoulées sur le côté sur les dépôts environnants par la gravitation en régime plastique. Par ce processus, le diapirisme des mollisols, se sont formés d'une part des dômes de lignite typiques (diapirs), et d'autre part, par la fuite du lignite et le glissement des sédiments couvrants, des formes creuses d'un diamètre jusqu'à quelques centaines de mètres[10]. Dans les périodes interglaciaires qui ont suivi, en partie pendant la dernière (Éémien), il y a environ 110 000 ans, elles se sont remplies d'eau et ont formé des lacs, sur le bord desquels un très riche écosystème de plantes et d'animaux a vécu (notamment l'éléphant à défenses droites, l'aurochs, le bison et le daim). On y trouve aussi des restes de groupes de chasseurs-cueilleurs du paléolithique moyen (Neumark-Nord 1 et Neumark-Nord 2)[7] - [11] - [12].
Gisements fossilifères
Conservation et distribution des fossiles
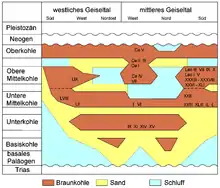
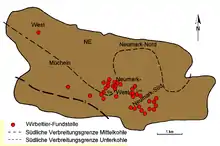

Alors que les plantes, et en partie aussi les invertébrés ont été trouvés dans tous les domaines des filons de lignite, les vertébrés sont reliés à des lieux définis, étroitement limités. En tout, on connaît plus de 80 sites, dont 36 présentent une proportion non négligeable de restes de vertébrés. Ceux-ci se trouvent principalement dans les filons inférieur et moyen, avec environ deux fois plus d'endroits dans le moyen que dans l'inférieur. Le filon supérieur ne comportait qu'un site de fouille intéressant, et était autrement largement dépourvu de fossiles, ce qui peut être rapporté aux transformations diagénétiques pendant le pléistocène sous l'action des conditions périglaciaires[9]. On n'a pu trouver aucune fossilisation dans le filon de base. La distribution spatiale des gîtes fossilifères de vertébrés reflète les conditions pour la conservation des fossiles, qui n'étaient pas uniformes dans l'ensemble du Geiseltal. C'est pourquoi la majeure partie des sites à vertébrés, et avant tout de ceux à squelettes ou parties molles largement préservés, se concentre dans le Geiseltal centre et sud, principalement près du seuil principal de Neumark, et dans la cuvette de Wernsdorf qui le jouxte à l'est. Le plus grand nombre de sites se trouve donc dans les mines à ciel ouvert Cecilie, Leo, Geiselröhlitz et Pfännerhall, s'étendant sur une surface de 20 km2 environ[13]. Vers le nord, l'ouest et en partie l'est, la qualité de la conservation des fossiles diminue considérablement[14] - [15].
Les gisements de vertébrés dans le lignite sont très rares dans le monde, et n'avaient pas été observés en Europe centrale à part dans le Geiseltal. La bonne conservation des fossiles dans le Geiseltal a plusieurs raisons. La raison principale est la présence d'eau calcaire circulant en sous-sol en provenance de roches avoisinantes triasiques, ici principalement du calcaire coquillier, en provenance du sud ou du sud-ouest du domaine de la dépression de Querfurt-Freyburg, pendant la formation du lignite. Cette eau calcaire a largement neutralisé les propriétés destructrices de l'acide humique des marais du Geiseltal d'alors, mais, comme on peut le voir sur la carte, n'était efficace que localement. En second lieu, l'acide silicique a aussi contribué à la bonne conservation des fossiles. Une autre influence sur la conservation des cadavres d'animaux est attribuée aux nombreuses inondations prouvées notamment dans le filon moyen, qui conduisaient à une couverture rapide des cadavres par des sédiments[16]. Les fossiles, comme le lignite, contiennent lorsqu'ils viennent d'être extraits, jusqu'à 50 % d'eau, et pendant le séchage, se désagrègent très vite, en s'effeuillant puis en tombant en poussière. C'est pourquoi, dès le début des années 1930, on a mis au point spécialement dans le Geiseltal pour la préservation des fossiles une méthode de tirage à la laque (de)[3] - [17].
Types de gisements
Outre les trouvailles isolées ou éparses, ainsi que les fossiles occasionnellement trouvés dans les carottes on peut en tout distinguer trois différents types de gisements de fossiles de vertébrés, qui peuvent en partie survenir en différentes combinaisons[3] - [18] - [19] :
- Entonnoirs d'effondrement
Il s'agit ici de dépressions circulaires, du genre doline, d'aspect symétrique. Elles sont dues à la dissolution du gypse dans le grès bigarré sous-jacent. En règle générale, les entonnoirs d'effondrement se reconnaissent aux perturbations des strates sur leurs bords, et aux déplacements tectoniques qui en résultent. On distingue deux types d'entonnoirs : l'entonnoir d'effondrement proprement dit, d'un diamètre de 12 à 18 mètres et l'entonnoir de tassement, plus petit et plus plat, de diamètre 3 à 8 mètres. Les enfoncements étaient souvent remplis d'eau à leur formation, et formaient de petits étangs ou mares. Les berges souvent raides formaient alors des pièges naturels pour les vertébrés, ce qui créait un cimetière naturel. Le sapropèle (boue de putréfaction) créé par le dépôt rapide de sédiments, et les conditions anaérobies ont facilité la fossilisation.
- Champs de cadavres
Ce sont des domaines plus étendus avec des gisements de fossiles, qui s'étendent de 80 à 100 mètres. La plupart du temps, les fossiles se trouvent dans des dépressions de grande surface, et en règle générale recouverts d'une couche de lignite de 20 à 30 centimètres. Ce sont des restes de domaines terrestres ou marécageux envahis par des inondations. Ils sont concentrés dans le Geiseltal central.
- Ruisseaux
Les ruisseaux, qui traversaient en partie le Geiseltal du domaine de calcaire coquillier occidental et débouchaient dans les bassins locaux, sont semblables aux champs de cadavres. Ce sont des creux en forme de gouttière dans le lignite, dans lesquels s'était déposé du sable en couches obliques. Ce n'est que dans les domaines plus profonds des bassins que l'on trouve des sédiments argileux, ce qui fait conclure à la diminution de la force d'entraînement par le courant des ruisseaux. La plupart des animaux sont venus mourir là comme près des entonnoirs d'effondrement à cause des rives souvent abruptes, mais y ont aussi été la proie de nombreux prédateurs.
Les entonnoirs d'effondrement et les champs de cadavres sont les plus fréquents, les ruisseaux sont assez rares, et n'ont été découverts qu'au milieu des années 1950. L'état de conservation des vertébrés dans les champs de cadavres et les ruisseaux est comparable, et fait voir certains déplacements par le mouvement de l'eau, liés à une dissociation des squelettes. Dans les entonnoirs d'effondrement, les restes de squelette complets n'apparaissent que dans les parties centrales, les plus profondes, là où les cadavres étaient complètement submergés. Au bord des entonnoirs, on peut observer des nettes désarticulations des squelettes[18] - [20]. Les trouvailles importantes de tissus mous fossilisés ne proviennent que des entonnoirs d'effondrement et des champs de cadavres[21].
Découvertes
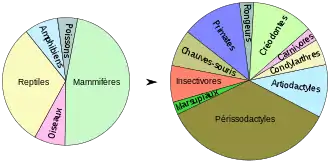
Les matériaux fossiles du lignite du Geiseltal sont très foisonnants et comprennent des restes de plantes et d'animaux. Les matériaux végétaux se présentent sous la forme de restes microscopiques et macroscopiques. La faune est représentée par des invertébrés et par des vertébrés dont il reste aussi un grand nombre de squelettes complets. Il faut souligner la bonne conservation des tissus mous, qui ailleurs, en général, sont très mal conservés dans les fossiles. Malgré les bonnes conditions de conservation, une assez grande partie des fossiles ne sont pas autochtones, mais ont été transportés pendant la fossilisation, à cause de l'écoulement de l'eau. Ceci est particulièrement vrai pour les champs de cadavres et les ruisseaux. En général, ces déplacements ne sont pas très importants, mais une partie peut être substantiellement déplacée. Le nombre exact des découvertes n'est pas précis, la collection de l'ex-musée du Geiseltal comprend en gros 30 000 objets, en majorité des vertébrés. Pendant la phase intensive de fouilles des années 1960, cette collection s'accroissait de 5 830 objets par an. Une analyse de plus de 10 000 restes de vertébrés donna avec 5 000 pièces presque une moitié de mammifères, tandis que les reptiles avec 2 000 pièces sont le deuxième groupe le plus représenté. L'exploitation scientifique de l'ensemble des fossiles continue comme par le passé, de nombreux êtres fossiles ont reçu à partir des trouvailles du Geiseltal leur première description. Les vertébrés à eux seuls appartiennent à plus de 120 taxons[3].
Flore

Parmi les fort nombreux restes de plantes, on trouve, à côté de pollen et de spores qui forment la microflore et représentent plus de 100 taxons pertinents pour la stratigraphie[22], ce sont avant tout les restes macroscopiques qui sont importants. Ceux-ci comprennent des feuilles, des tiges, des branches, des écorces et des troncs, ainsi que des fruits, des semences et des fleurs. Simplement avec les fruits et les semences, on peut au moins déterminer 18 familles. Au total, on connaît plus de deux douzaines de familles avec environ 40 genres. Outre les algues, les mousses, les psilophytes et les fougères - ces dernières en partie en enrichissements concentrés notamment de blechnums en épi et de lygodiums[23], il y a surtout des restes de spermaphytes plus évoluées. Les gymnospermes sont représentées par des cycadales parfois avec leur houppe entière, et des conifères avec des cônes et des rameaux. Ces derniers comprennent des pins et des sequoioideae, qui apparaissent en partie accumulés. Les restes d'angiospermes sont notablement plus riches. Le plus grand groupe est celui des dicotylédones, avec environ 20 familles. Ce groupe contient des lauracées et des fagacées. Ces dernières forment parfois d'épaisses couches de feuilles du genre Dryophyllum. Les myricacées, surtout des Comptonia, sont très nombreuses. Les restes de plantes prouvés en grand nombre proviennent de tilleuls, d'icacinacées et de myrtacées, parmi lesquelles le Rhodomyrtophyllum est dominant. Les feuilles longues et étroites d'apocynophyllum de la famille des apocynacées sont caractéristiques. On attribue aussi à cette famille des restes d'écorces et les tubes à latex fossilisés, appelés « poils de singe ». Les monocotylédones sont peu diversifiés. Parmi ceux-ci, les palmiers paraissent en grand nombre, et sont représentés en particulier par des sabals et des phoenicites. Mais la plupart des troncs de palmiers trouvés ne peuvent pas être identifiés avec précision dans la taxonomie[24]. D'autres représentants des monocotylédones ont aussi été décrits, par exemple des liliacées et des pandanacées[25].
Invertébrés
Les invertébrés ont été évoqués pour la première fois en 1913[26], et sont très nombreux. Les mollusques, auxquels appartiennent les coquillages et les escargots, et qui apparaissent parfois en masses, sont représentés par environ 20 genres, et sont parfaitement adaptés pour la reconstruction de biotopes locaux[27]. Ceci vaut aussi pour les ostracodes, représentés par 10 genres[28]. D'autres trouvailles, d'arthropodes, sont attribuées à des décapodes. Mais ceux-ci, avec seulement 15 exemplaires, sont extrêmement rares, et la plupart du temps conservés avec seulement des restes de la carapace[29]. Les découvertes les plus fréquentes d'invertébrés sont les insectes. Ici dominent les coléoptères, et parmi eux, 28 % de buprestidae. Le genre Psiloptera est présent avec huit espèces démontrées, mais buprestis et anthaxia sont représentés par plusieurs espèces. À côté, on trouve au moins six autres familles de coléoptères, notamment des scarabées, des dryopidae et des chrysomèles. Comme on ne retrouve presque que les dos des insectes, on peut retrouver la coloration souvent somptueuse des animaux, qui permet dans bien des cas, mais pas tous, une assignation précise de l'espèce[30]. On a aussi pu trouver des insectes volants, comme des éphémères et des libellules, mais les demoiselles ne sont identifiées que par des petites pontes de quelques millimètres[31]. Les arachnides et mille-pattes sont en très petit nombre. La découverte d'un nématomorphe du genre gordius, parasite d'insectes, représente une particularité, car ce genre de parasite n'apparaît que rarement comme fossile[32].
Poissons et amphibiens
Les poissons sont très nombreux, avec environ 2 000 fossiles trouvés, parmi lesquels de nombreux ont un squelette complet. Mais on ne trouve que 5 familles, et presque autant de genres. Les plus nombreux sont les poissons osseux, représentés par Thaumaturus et Palaeoesox. Le premier appartient à la famille des salmonidés, le second aux ésociformes[33]. Un autre poisson osseux est l'Anthracoperca, un perciforme. Cyclurus est un représentant de la famille actuellement presque éteinte des Holostei[34] - [35].
Parmi les amphibiens, on trouve principalement des grenouilles et des urodèles, dont on a de 200 à 300 exemplaires de chaque, sans compter les nombreux os isolés. Les squelettes articulés des grenouilles nous sont tous arrivés dans un état très fragile. Le plus commun est Eopelobates, de la famille des Pelobates, à laquelle appartient l'espèce pélobate brun actuelle. Comme ce dernier, Eopelobates vivait essentiellement à terre. Par contre, le palaeobatinopsis, aussi abondant, de la famille des Palaeobatrachidae, une famille aujourd'hui éteinte, vivait dans l'eau. D'autres matériaux fossiles peuvent appartenir à une espèce très voisine[36]. Parmi les urodèles, Palaeoproteus est particulièrement abondant. Il s'agit d'un protée amphibie, qui atteignait 25 centimètres de long, avec des membres très courts. Plus rare est le tylototriton. Ce genre existe encore à l'est et au sud-est de l'Asie, et est très proche de la sous-famille des vraies salamandres[37].
Reptiles et oiseaux
Les reptiles font partie des découvertes de fossiles les plus nombreuses et peuvent être attribués à près d'une douzaine de familles avec environ 20 genres. En raison du milieu humide, de nombreuses tortues sont parvenues, en particulier les restes de leurs carapaces, qui sont la plupart du temps aplatis par le poids des sédiments. Toutes les tortues connues dans le Geiseltal appartiennent aux cryptodires, et on en compte environ 500 exemplaires. On y compte notamment le petit genre Geomyda de la famille des Emydidae, avec une carapace bien ossifiée d'environ 18 cm de long. À l'autre bout des ordres de grandeur, on trouve Geochelone, une tortue terrestre, qui possédait une carapace de 1,2 m de long[38]. La famille des tortues à carapace molle est représentée par Trionyx, dont la carapace atteignait environ 30 cm de long. Cet animal était bien adapté à la vie dans l'eau[39].
Les squamates sont démontrés par près de 300 exemplaires complets, sans compter les nombreux restes démembrés. Parmi eux, eolacerta était un grand animal, qui atteignait 60 cm de long, et que l'on a retrouvé avec plusieurs squelettes complets et des mues. On le compte souvent parmi les vrais lézards, cependant certains critères anatomiques rendent ce classement incertain[40]. Geiseltaliellus est à classer parmi les iguanes, c'est un saurien léger, avec une queue extrêmement longue et une peau finement écailleuse, probablement un arboricole. En raison des dimensions très courtes de ses membres antérieurs, on suppose que le Geiseltaliellus pouvait à l'occasion se déplacer uniquement sur ses membres postérieurs, comme les basilics actuels[41]. Il n'y a que très peu de cousins des varans actuels. Un squelette partiel mal conservé d'eosaniwa a un crâne de 19 cm de long, et avec cela, ce saurien était un des plus grands du Geiseltal[42]. 60 exemplaires complets sont à rapporter aux serpents, dont notamment deux genres des serpents géants de la famille des boas. Paleryx atteignait 2,3 m de long, et pour un exemplaire largement complet, on a pu reconnaître 243 vertèbres[43]. Ensuite vient un palaeopython, ainsi que quelques faux coraux[44] - [45].

Les crocodiles sont également nombreux, avec 120 squelettes complets, plus de nombreux os et dents isolés. Allognathosuchus y représente les alligators. Il atteint 2 mètres et possède un museau très étroit. Diplocynodon fait partie des formes les plus nombreuses de crocodiles dans le Geiseltal et on le range dans la famille éteinte des Leidyosuchidae. Il vivait dans de petites étendues d'eau, tout en atteignant bien 2,5 mètres de long. Boverisuchus était à peu près aussi grand et nombreux, mais il représentait les vrais crocodiles. En raison des phalanges terminales de ses doigts élargies en sabot, on suppose que ce crocodile avait un mode de vie plutôt terrestre. Le plus grand prédateur du Geiseltal était Asiatosuchus, dont la longueur du corps atteignait 5 mètres. Écologiquement, il ressemblait au crocodile du Nil actuel et préférait les eaux ouvertes. On en a trouvé de nombreux squelettes complets. Par contre, Bergisuchus, dont l'holotype a été découvert sur le site fossilifère de Messel, n'apparaît que très rarement sous forme de quelques fragments de mandibule. C'était aussi probablement un crocodile plutôt terrestre, de petite taille avec sa longueur de 1,5 m[46]. Des recherches détaillées ont conclu que tous les crocodiles du Geiseltal occupaient des niches écologiques différentes[47]. En outre, on a trouvé des fragments d'œufs de crocodiles[48] et de nombreux gastrolithes ou pierres d'estomac, qui attestent même en l'absence de fossiles la présence des crocodiles[3].

Les restes d'oiseaux sont plutôt rares, avec une demi-douzaine de familles et le double de genres. La plupart du temps, on n'a trouvé que des restes d'extrémités. Saurornis paraît assez primitif. Il atteint la taille d'un nandou, et on ne le connaît que par un tarsométatarse de 20 cm de long dont l'extrémité articulaire supérieure n'est pas encore complètement fusionnée avec les trois métatarsiens originaux comme c'est la règle pour les oiseaux actuels[49]. Palaeotis est un relativement petit représentant des oiseaux coureurs, dont on n'a aussi trouvé que les extrémités postérieures[50]. Gastornis (ou Diatryma), avec sa hauteur totale moyenne de 1,8 m est nettement plus grand. Il y a une quarantaine de restes de ce grand ratite, plusieurs os des jambes, mais aussi des restes de crânes et des parties d'ailes, ce qui représente la plus grande collection de tous les gisements au sein de l'Europe[51] - [52]. On a longtemps considéré gastornis comme un carnivore, jusqu'à ce que des études isotopiques sur quelques os du Geiseltal montrent que l'oiseau était principalement végétarien[53]. Eocathartes a été initialement compté parmi les vrai carnivores. Il était censé représenter les vautours du Nouveau Monde dans le Geiseltal et a été décrit à partir de parties du squelette et de plumes. En outre, Geiseloceros, comme membre de la famille des bucérotidés (calaos) avaient un statut exotique. Son existence a été démontrée à partir d'os de jambes et de restes d'ailes non démembrés, sur lesquels on pouvait encore reconnaître les rémiges d'un bleu brillant[54]. Ces deux taxons sont placés dans la famille des Strigogyps, une famille de grands ratites[55]. Il a aussi été montré l'existence d'Aegialornis, un apodidé fossile[56].
Mammifères
La très riche faune de mammifères comprend des restes d'environ deux douzaines de familles, avec 50 genres. Les plus primitifs d'entre eux sont des marsupiaux, représentés par Peratherium et Amphiperatherium. Il existe plusieurs crânes et mandibules, surtout de peratherium[57]. Ces deux genres sont proches de la lignée ancestrale de l'opossum sud-américain. Il est par contre difficile de situer Microtarsioides sur le plan taxonomique, car un squelette relativement complet nous est parvenu, mais il provient d'un animal juvénile[58].
Les euthériens, animaux placentaires, sont réellement plus riches et variés. Quelques os longs de forme puissante caractéristique ont été attribués au tamandu européen[58]. Initialement considéré comme un myrmécophagidé[59], il présente, après des recherches plus approfondies la forme de base des pangolins, avec le même mode de vie[60]. Parmi les petits mammifères, on compte Heterohyus, caractérisé par deux doigts allongés et des incisives recourbées en cercle. Un squelette partiel et une mandibule attestent l'existence de Leptictidium. Cet animal prédateur se déplaçait en sautant sur ses membres arrière, ce que montre la position de base des membres de l'ordre des leptictida, et qui ne se présente plus chez les insectivores actuels[61]. Le Buxolestes, qui apparaît rarement, était au contraire un animal trapu, semi-aquatique de la famille des Pantolestidae, dont l'extérieur et la manière de vivre correspondaient à peu près à la loutre d'Europe. Très rare aussi, mais représenté par de nombreuses mandibules en partie avec toutes leurs dents, est l'insectivore Saturninia, de la famille des nyctitheriidae[58] - [62]. Les rongeurs sont rares et apparaissent sous peu de formes. Ailuravus était très grand, à peu près 1 m de long, et rappelle les sciuridés actuels. Peu de restes de mâchoires proviennent de Masillamys, nettement plus petit, appelé souris de Messel d'après le site fossilifère de Messel, avec une très longue queue et une longueur totale de 40 cm. Les restes de chauves-souris sont importants, on en trouve 25 squelettes complets avec des parties des ailes. On peut surtout les ranger dans les genres Matthesia et Cecilionycteris. Ils appartiennent à la famille des Palaeochiropterygidae, dont les représentants, d'après la structure des osselets de leurs oreilles, chassaient déjà par écholocalisation dans la gamme des 30 à 70 kHz[63].
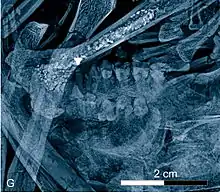
Des restes remarquables proviennent aussi de primates. Ici, les plus importants sont les adapiformes, dont on trouve plusieurs genres dans le Geiseltal, et qui sont peut-être des précurseurs des lémuriens. On a trouvé plusieurs crânes et mandibules d'europolemur. Un des rares squelettes presque entiers de primate adapiforme dans le monde est celui d'un godinotia. Ce primate précoce, qui appartenait à la proche parenté du darwinius trouvé à Messel, possédait un crâne court sur le visage et des grandes orbites, qui suggèrent une activité nocturne[64] - [65] - [66]. Nannopithex apparaît très diversifié, avec beaucoup d'espèces. Il appartient à une famille plus moderne de primates[67].
Les créodontes, aujourd'hui éteints, vivaient une vie de prédation. On y compte surtout les Hyaenodontidae, avec environ 50 trouvailles, dont surtout des mandibules, mais aussi des crânes, des fragments de mâchoire supérieure, et des restes isolés de l'appareil locomoteur. On y a classé des eurotheriums, des prodissopsalis et des leonhardtinas, les deux derniers étant les plus nombreux et comprenant chacun une douzaine de fossiles. Au total, tous les créodontes trouvés dans le Geiseltal étaient relativement petits, par exemple un crâne complet d'eurotherium n'atteignait que 12 cm de long[68] - [69]. Une seule prémolaire supérieure arrière représente les vrais carnivores. Elle fut initialement attribuée à un miacis, mais maintenant à un quercygale[70]. Seuls deux fragments osseux peuvent être attribués à des condylarthra, une famille aussi éteinte actuellement. Un crâne de 12 cm de long appartient à un vulpavoides et un fragment de mandibule de 5 cm de long à un pugiodens[71].

Les matériaux fossiles d'artiodactyles sont proportionnellement plus importants, avec environ 170 fossiles appartenant à 60 individus. Les artiodactyles du Geiseltal se distinguaient tous par un corps de conformation primitive, avec une courbe du dos bombée vers le haut, de très longues queues et des membres courts à l'avant, et longs à l'arrière. Ils étaient nettement plus petits que leurs parents actuels, et atteignaient au plus la taille d'un cochon. Un des plus primitifs était diacodexis, de la famille des diacodexeidae, mais dont on n'a trouvé que quelques restes d'os. Un peu plus élaboré était eurodexis, de la famille proche des dichobunidae[72]. Dans la proche parenté, on a trouvé messelobunodon et aumelasia, les deux n'étant cependant représentés que par peu de fossiles. Les plus nombreux observés jusqu'ici sont les choeropotamidae, un peu plus élaborés, que l'on place dans la proche parenté des hippopotames, et qui sont répandus en Europe actuelle de façon endémique. Un squelette de masillabune presque complet a été trouvé[73], alors que hallebune, de la même famille, n'est représenté que par un petit nombre de fragments de mâchoire[74] - [75]. Plusieurs restes de squelettes, dont sept presque entiers appartiennent à des amphirhagatheriums. Cet animal de 70 cm de long, pesant 4 kg était non seulement le plus nombreux mais aussi le plus grand des artiodactyles du Geiseltal, et est largement distribué dans les filons inférieur et moyen[76] - [77]. En outre, il y a quelques rares restes d'autres artiodactyles comme haplobunodon ou rhagatherium[78].

C'est parmi les périssodactyles que se trouve le matériel fossile le plus abondant. Ceux-ci avaient aussi une conformation primitive, avec un dos voûté, quatre doigts aux pattes de devant et trois à celles de derrière, ce qui n'arrive plus actuellement que chez les tapirs. Plus de 310 mandibules et 120 restes de mâchoire supérieure, y compris deux squelettes complets reviennent à des proches parents des chevaux, de la famille des palaeotheriidae[79]. Il faut souligner le petit ancêtre du cheval propalaeotherium, représenté par plusieurs espèces. Un squelette complet, avec un corps long de 56 cm, un crâne long de 20 cm et une hauteur au garrot de 40 cm a été découvert en 1933. D'autres genres présents sont lophiotherium, hallensia et eurohippus. Il ne reste que quelques dents du très rare plagiolophus[80] - [81] - [82]. Lophiodon, un parent du tapir, est aussi très abondant, avec 180 individus. La longueur de son corps est de 2,5 m et il a une hauteur au garrot de 1 m, ce qui en fait le plus grand mammifère du Geiseltal. Sur une surface de 131 m2, on a découvert 110 individus, sans doute les restes d'une zone de dévoration par les crocodiles. Les fossiles de lophiodon sont connus dans tous les filons comportant des fossiles, le genre parcourant du bas vers le haut une augmentation substantielle de la taille du corps[83] - [84] - [85]. À côté de cela, son parent hyrachyus paraît plus rare, et ne peut être trouvé que grâce à environ 75 restes de mâchoires, et de quelques parties du squelette du corps[86]. Hyrachyus a une conformation plus moderne que lophiodon, et se tient au départ du développement des rhinocéros et des tapirs[87].
Parties molles, fossiles chimiques et restes de nourriture
Dans le lignite du Geiseltal, des restes des parties molles se sont aussi conservés, ce qui arrive extrêmement rarement. C'est en 1934 que des parties molles fossiles ont été identifiées pour la première fois. Elles appartiennent ainsi aux premières trouvailles de ce genre de l'éocène. Ce n'est qu'avec la mise au point du « tirage à la laque » que ces découvertes ont réussi. C'est ainsi que l'on a pu reconnaître chez des poissons les fins reliefs de leurs écailles, par exemple chez thaumaturus, et aussi chez des squamates, et plus rarement chez des crocodiles[88]. On a pu étudier la peau de nombreuses grenouilles, dont les différentes couches, l'épiderme et le derme, jusqu'aux cellules, sont identifiables[21]. Sur certains restes d'oiseaux, on a pu identifier des plumes, encore en partie colorées[89]. On a aussi retrouvé des membranes de l'aile de chauves-souris. On a retrouvé des parties de pelage de mammifères, comme amphirhagatherium ou plus rarement propalaeotherium[90]. On a aussi réussi à identifier des muscles, du cartilage et autres tissus, jusqu'à des cellules sanguines[91].
En outre, on a pu reconnaître la coloration initiale de certaines parties molles, en particulier de la peau. Ainsi, le poisson palaeoesox avait le corps zébré, avec un dos foncé, tandis que pour les grenouilles, on a pu observer une couleur de peau verdâtre. Chez les insectes, les nuances de couleur des animaux ont été conservées, et dans les plantes, la chlorophylle a survécu[92]. À ces fossiles chimiques appartient aussi le « poil de singe, » une substance fibreuse, aplatie par le séjour dans le lignite, de couleur brun clair, qui paraît souvent en grandes quantités. Il s'agit de vaisseaux à latex de buissons produisant du caoutchouc de la famille des apocynaceae, dont les feuilles du genre apocynophyllum sont présentes dans le Geiseltal. Mais le poil de singe est attribué plutôt au coumoxylon, du taxon du genre récent couma[93].
D'autres trouvailles sont avant tout les restes de nourriture de nombreux animaux, qui ont été découverts tôt aussi, comme en 1935 des masses verdâtres de plantes du parent du tapir, le lophiodon[94]. Elles viennent aussi d'autres mammifères comme le cheval antique propalaeotherium[95], ou l'artiodactyle amphirhagatherium[76] - [77].
Datation
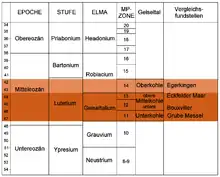
Pour une détermination précise de l'âge du lignite du Geiseltal, les restes de la faune sont importants, en particulier ceux des mammifères, qui permettent une comparaison avec d'autres gisements fossillifères et des conclusions. Cette détermination de l'âge biostratigraphique est largement définie par les modifications de la morphologie des dents de groupes bien définis de mammifères, ce qui permet de déduire la présence dans le temps d'espèces et de genres maintenant disparus. Les premiers fossiles de vertébrés découverts au début du XXe siècle, des restes de dents du parent du tapir, le lophiodon, ont permis l'insertion du lignite dans l'époque géologique de l'éocène moyen. C'est ainsi que l'on a pu déterminer avec précision l'âge relatif des lignites, qui n'avait jusque là été que supposée[96]. L'éocène moyen a un âge de 47,8 à 41,3 millions d'années et est composé d'une couche inférieure, le lutetium et d'une couche supérieure, le bartonium. Par la présence de certaines formes de mammifères, l'âge précis du lignite du Geiseltal peut être assez précisément déterminé. On place ainsi les riches fossiles du filon inférieur jusqu'au filon moyen supérieur dans la zone du geiseltalium, une période au sein de la stratigraphie des mammifères terrestres européens (European Land Mammal Ages, ELMA). La faune du Geiseltal sert de référence pour le geiseltalium, qui forme un segment moyen du lutetium. Les quelques fossiles du niveau supérieur sont placés dans une zone plus récente, le robiacium. Le début du geiseltalium va ainsi de l'apparition des formes précoces du cheval primitif propalaeotherium, mais aussi de son parent le lophiotherium, ou du primate europolemur, tandis que sa fin est indiquée par la première apparition de primates comme l'adapis[14] - [97].
On ne possède pas de datation absolue au moyen de mesures radiométriques pour le Geiseltal. Cependant, ce genre de mesure absolue a été entreprise pour des gisements d'âge égal. Ainsi, la datation par le potassium-argon a fourni des dates de l'important site fossilifère de Messel, qui du point de vue biostratigraphique correspond au filon inférieur du Geiseltal. Ces dates donnent un âge de 47,8 millions d'années. Comme le basalte étudié provient d'une région située sous les sédiments contenant les fossiles de Messel, cet âge doit être considéré comme un maximum, les fossiles eux-mêmes étant quelque peu plus jeunes[98]. À l'opposé, le maar d'Eckfeld est un équivalent biostratigraphique de la couche supérieure du filon moyen. Les données obtenues par la même méthode à partir des pyroclastites situées sous la couche de fossiles, ont donné une valeur de 44,3 millions d'années. Ceci doit aussi être considéré comme une limite inférieure de l'âge[99]. On en déduit une estimation de l'âge du geiseltalium, auquel on donne actuellement une valeur de 47,4 à 43,4 millions d'années, ce qui cale avec sûreté les trouvailles du Geiseltal.
Reconstruction du paysage
Les nombreux restes de plantes et d'animaux, mais aussi le nombre élevé de données géologiques permettent une reconstruction assez précise du paysage d'alors. À l'éocène moyen, le Geiseltal était un paysage de marécage très riche en eau, qui s'étendait en direction nord-sud sur une longueur de 4 à 5 km et se déversait à proximité de la côte dans une baie pénétrant largement dans la terre ferme. Cette découpure de la côte, connue comme l'estuaire moyen-allemand était soumise au moins à certaines époques aux marées, comme le montrent des pollens fossiles du genre nypa du filon moyen[100]. Au milieu de l'éocène (plus précisément au lutetium), cet estuaire formait de nombreuses baies, des bassins et des dépressions, qui favorisaient la formation de lignite, comme à Helmstedt dans le nord-ouest ou à Egeln et Edderitz vers le sud. Ce système ramifié a aussi conduit dans le Geiseltal à la formation d'alluvions marines, saumâtres, fluviatiles et lacustres. Le climat alors subtropical contribuait à la formation de nombreux marais, étangs et mares, avec une riche végétation, sur les marges de l'estuaire. Par la carbonisation des matériaux des plantes mortes, il est survenu pendant l'espace de six à peut-être huit millions d'années la formation de puissants filons de lignite[4]. Le territoire était limité au sud par un plateau de calcaire coquillier, avec des pentes abruptes vers le nord. Ce plateau était lui-même en partie karstifié. Par érosion chimique, des eaux calcaires se formaient, s'écoulaient vers les marais et aboutissaient à une préservation remarquable des fossiles[101].
Le territoire marécageux était envahi par de nombreux petits étangs et mares, d'un diamètre de 8 à 10 m, signalés par des entonnoirs d'effondrement et qui servaient d'abreuvoirs pour le monde animal fossile. Sur leurs rives poussaient des écosystèmes d'acrostichum, de myricaceae et de restionaceae. Plus loin, on trouvait des paysages de buissons et de forêts de palmiers et de résineux, parsemés de séquoias et autres arbres résineux ou feuillus, riches en herbes et en lumière, qui se déployaient sur plusieurs étages. Ces forêts étaient parcourues par un réseau de cours d'eau faisant des méandres et débordant de leurs rives périodiquement. Vers le sud, en direction du plateau de calcaire, ces forêts se transformaient en forêts vierges épaisses de lauracées. Dans l'ensemble, le climat était subtropical, avec des hivers sans gelée et secs, avec une température moyenne minimale de 5 °C, et des étés humides avec une période de végétation jusqu'à 9 mois et en moyenne 25 °C pendant le mois le plus chaud, ce que l'on a pu établir en raison des nombreux fruits et semences. Le paysage avait ainsi une périodicité de pluies et de temps sec, ce que l'on peut retrouver par les cernes des arbres et de fines varves dans le lignite. C'est pourquoi il faut considérer la chute des feuilles comme probable, si bien qu'elles formaient une couche légère répandue sur le sol. Cependant, de l'évolution des filons inférieur au moyen, on peut déduire une sécheresse croissante du climat[47] - [77] - [102].
Ce paysage varié était habité par de nombreux animaux. Dans les forêts vivaient des mammifères végétariens comme lophiodon, propalaeotherium et amphirhagatherium. La riche faune d'insectes était à son tour la base de la nourriture de nombreux autres animaux, comme des grenouilles, des urodèles, des squamates, mais aussi des leptictida, des chauves-souris et quelques primates. Les étangs et les mares hébergeaient des animaux aquatiques ou amphibies. À noter les nombreuses formes de crocodiles, qui préféraient un climat significativement chaud, comme actuellement, et qui sont donc un indicateur important sur le climat[47] - [102].
Comparaison avec des gisements fossilifères régionaux importants
Dans le voisinage régional du Geiseltal, les plus importants sont les mines à ciel ouvert de lignite de même âge des environs de Leipzig (le bassin de l'Elster blanche), ou la zone de Helmstedt. Bien qu'il n'y ait pas de restes de vertébrés, les fossiles végétaux sont innombrables. Ainsi, dans les mines de Helmstedt, on trouve des restes macroscopiques comme des cônes de conifères, et des feuilles de palmiers, dans celle de Profen de nombreux fossiles de dicotylédones[24]. L'histoire de la végétation, déduite de la microflore, correspond largement à celle du Geiseltal[22]. On ne connaît pas de gisement de fossiles de vertébrés dans une région proche. Mais avec les gisements de Walbeck, près d'Oebisfelde-Weferlingen dans le nord-ouest de Saxe-Anhalt, il y a une des plus nombreuses collections connues de mammifères du paléocène moyen, il y a juste 60 millions d'années. Cette faune, comprenant plusieurs milliers de restes d'os et de dents se distingue de celle du Geiseltal par la dominance de mammifères primitifs des familles des procreodi, des condylarthra ou des leptictida, tandis que les représentants de lignages plus modernes, encore subsistants aujourd'hui comme les primates sont rares, et les artiodactyles et périssodactyles manquent complètement. Walbeck présente ainsi un important témoignage de la phase précoce de radiation évolutive des mammifères peu après l'extinction des dinosaures[103] - [104].
Dans la comparaison à distance supérieure à la région, les sites de Messel en Hesse et le maar d'Eckfeld en Rhénanie-Palatinat ont une importance remarquable. Ces deux sites sont dans des anciens maars, Messel est contemporain du filon inférieur du Geiseltal, tandis qu'Eckfeld l'est du filon moyen supérieur. Outre les fossiles de plantes et d'invertébrés, on connaît un grand nombre de restes de vertébrés des deux sites, à Messel, environ 130 taxons, la faune d'Eckfeld étant plus restreinte. Tandis que la composition de la faune des mammifères est en général en accord, il y a dans le détail des différences significatives. Par exemple, dans le Geiseltal et à Eckfeld, les rongeurs et les chauves-souris sont sous-représentés[63], tandis qu'ils sont plus nombreux à Messel, et de formes plus variées. Par contre, Messel se distingue par un faible nombre de primates et d'artiodactyles, qui sont bien attestés dans le Geiseltal et à Eckfeld, sous de nombreuses formes, malgré des différences marquées dans l'abondance des divers genres. On peut trouver des différences semblables dans les genres de mammifères, par exemple lophiodon, un périssodactyle, qui est une des formes dominantes dans le Geiseltal, n'apparaît à Messel que par un juvénile et quelques dents, et à Eckfeld par une mandibule et aussi quelques dents. Il est remarquable à Eckfeld de remarquer l'absence jusqu'à présent de familles primitives de mammifères, comme les créodontes ou les condylarthra, qui apparaissent à Messel et dans le Geiseltal au moins en petit nombre[3] - [105] - [106]. En outre, on trouve des différences dans d'autres groupes de fossiles. Par exemple, les buprestidae, avec plus d'un quart de tous les fossiles de coléoptères, sont relativement nombreux dans le Geiseltal, tandis qu'à Messel, avec moins d'un dixième, ils sont relativement rares, mais on y trouve en tout une plus grande variété de formes de coléoptères. Le fait que par exemple seules peu de formes de buprestidae apparaissent dans les deux sites, indique d'importantes différences écologiques, qui se répercutent aussi dans la multiplicité différente des diverses familles de plantes et d'animaux, et montrent que les gisements fossilifères ne représentent à chaque fois qu'une coupe du riche paysage d'alors[30].
Histoire de la recherche
Fouilles scientifiques

Bien que l'exploitation du lignite dans le Geiseltal soit démontrée dès 1698[2] - [107], les premiers fossiles ont été découverts relativement tard. La première trouvaille de fossile date de 1908, et comprend des restes du parent du tapir, le lophiodon, rassemblé par un contremaître dans la mine Cecilie. D'autres trouvailles ont été sauvées en 1912, plusieurs restes de mâchoire de trois individus du même genre. L'année suivante, ce sont pour la première fois des restes de tortues qui ont été observés dans une couche de lignite contenant des plantes, mais une partie des fossiles trouvés, une fois dégagés, se sont effrités. Quelques fossiles conservés ont été confiés à l'Établissement géologique du Land de Prusse à Berlin[26] - [96].
En 1925 commencent les fouilles scientifiques systématiques, que le géologue Johannes Walther (1860–1937) a commencées, et que l'université Martin-Luther de Halle-Wittenberg a coordonnées. Après la retraite de Walther, les fouilles ont été continuées par Johannes Weigelt (1890–1948). Pour obtenir une meilleure durabilité des fossiles, Ehrhard Voigt (1905–2004) mit au point au début des années 1930 la méthode du « tirage à la laque[17] » . Un premier succès d'envergure a été la découverte en 1933 d'un squelette complet du cheval primitif propalaeotherium, de 74 cm de long, dans la mine Cecilie, au centre d'un entonnoir d'effondrement, au milieu de nombreux restes de crocodiles et de squelettes partiellement complets de lézards[20]. Dans la même mine, seulement un an après, a été étudié le seul gisement important dans le filon supérieur, avec plus de 20 individus de lophiodon, dont un squelette presque complet[94]. Les recherches scientifiques dans la mine Cecilie se sont arrêtées en 1935 en raison de l'épuisement des ressources en lignite. D'autres fouilles ont eu alors lieu dans la mine Leonhardt, mais elles se sont arrêtées en 1938. La déclaration de la seconde Guerre mondiale en 1939 termina définitivement cette première période importante de recherches[13].
En 1949, les activités de fouille ont repris, et par la suite, elles ont été poussées dans un cadre plus vaste par le gouvernement de la RDA. En outre, de nombreux scientifiques d'autres nations ont pu être associés à la mise en valeur des fossiles. Par l'agrandissement de la mine, qui a atteint son maximum au milieu du XXe siècle, de nombreux autres gisements ont pu être ouverts, notamment dans les mines Pfännerhall et Mücheln (qui comprenait les domaines précédents Pauline, Elisabeth, Emma et Elise II du Geiseltal ouest). C'est avec le gisement XIV que les scientifiques en ont découvert un des plus riches, et ont pu ajouter un nouveau type de gisements à ceux déjà définis par Weigelt, celui des « ruisseaux ». L'exploitation élevée du lignite a conduit à ce que les mesures de fouilles accompagnant l'exploitation soient renforcées aussi, si bien que l'étude du Geiseltal a atteint un sommet dans les années 1960 et 1970. Ce n'est qu'avec l'épuisement des ressources en lignite qui s'annonçait (le Geiseltal était déjà épuisé à l'est et au sud-est dans les années 1970, et les mines abandonnées partiellement inondées) et le déplacement de l'exploitation vers les parties plus au nord et à l'ouest du Geiseltal, où la conservation des fossiles n'était pas aussi bonne, les recherches scientifiques ont diminué et se sont terminées pour l'instant en 1985. Pendant cette phase de recherche, plus de 55 nouveaux gisements ont été découverts, dont 23 avec des restes de vertébrés[13] - [15].
Après le changement politique en RDA en 1989, la situation changea fondamentalement. Tandis qu'initialement, l'exploitation du lignite au Geiseltal ouest devait continuer jusqu'en 1998, elle fut arrêtée en mi-1993 en raison du changement des conditions du marché, et on commença à renaturer l'ensemble du territoire de la mine. C'est ainsi que du printemps à l'automne 1992 on en arriva aux dernières recherches sur un gisement au Geiseltal sud, avec la participation de collaborateurs de l'institut de recherche Senckenberg. Dans l'année suivante, ce dernier gisement à vertébrés était inondé par de l'eau souterraine qui remontait[15]. Entre 2000 et 2003, les dernières activités de terrain dans le Geiseltal ouest ont eu lieu, toujours avec l'institut de recherche Senckenberg, dans un filon de lignite de 20 m de puissance, qui contenait d'innombrables matériaux végétaux, jusqu'à des troncs de 25 m de long. En raison des propriétés chimiques du lignite dans cette région du Geiseltal, on n'a pas pu trouver de restes de vertébrés[31]. En commence l'inondation prévue du Geiseltal ouest en « lac du Geiseltal, » qui atteint son niveau final au printemps 2011, ce qui crée le plus grand lac artificiel d'Europe centrale. Ainsi se terminent les activités scientifiques sur place[2].
Présentation des découvertes

Dès 1934, seulement 9 ans après le début des activités de fouille, et en raison de l'immense quantité de matériaux trouvés, le musée du Geiseltal a été ouvert dans la « Nouvelle résidence » érigée sur commande du cardinal Albert II de 1531 à 1537, près de la cathédrale de Halle. Le musée a servi en même temps de dépôt des collections et de lieu d'exposition, ce dernier étant aménagé sur une surface de 267 m2 dans la chapelle de la Toussaint. Les autres espaces de la Nouvelle résidence ont été utilisés par l'Institut de paléontologie de l'université Martin-Luther de Halle-Wittenberg. La mascotte du musée est le squelette attribué au propalaeotherium, en 1933. Peu avant la fin de la seconde Guerre mondiale, en 1945, l'exposition a été interrompue, les fossiles ayant dû être déménagés pour des raisons de sécurité[3] - [108].
En 1950, l'exposition a rouvert, sous le titre « Promenade à travers l'histoire géologique d'Allemagne centrale » - agrandie de huit salles et retravaillée professionnellement en 1954. À la suite de la réforme des universités de la fin des années 1960, il aurait fallu déménager le musée et la collection, ce qui a pu être évité par le directeur de l'époque Horst Werner Matthes. Cependant, la nouvelle exposition ouverte en 1950 a dû être fermée ou réduite. Des restrictions techniques et financières ont conduit au fait que les années suivantes, on n'a pas pu investir ni dans une modernisation de l'exposition, ni dans les réparations des bâtiments de la Nouvelle résidence, ce qui a résulté dans la nécessité de préserver certains fossiles notamment des fuites de pluie, ou du dégel. Ce n'est qu'après 1989, avec le changement politique en RDA que la situation s'est améliorée[108]. Cependant, les bâtiments de la Nouvelle résidence restaient en mauvais état. Quand l'Institut pour les sciences de la Terre s'installa en 2004 dans les locaux nouveaux et plus modernes sur le nouveau campus de la place von Seckendorf, seul le musée du Geiseltal resta sur place, avec ses collections, et il y fêta ses 70 ans d'existence[31]. Fin 2011, le musée ferma définitivement, si bien que les collections ne sont plus accessibles au public. Ce n'est que pour 2015 qu'il est planifié de rendre les trouvailles du Geiseltal accessibles au public dans une nouvelle exposition[109].
Bibliographie
- (de) Günter Krumbiegel, Ludwig Rüffle et Hartmut Haubold, Das eozäne Geiseltal: ein mitteleuropäisches Braunkohlenvorkommen und seine Pflanzen- und Tierwelt, Wittenberg, Ziemsen, (OCLC 9998655), p. 1–227.
- (de) Meinolf Hellmund, « Exkursion: Ehemaliges Geiseltalrevier, südwestlich von Halle (Saale). Aus der Vita des eozänen Geiseltales : 34. Tagung des Arbeitskreises für Wirbeltierpaläontologie der Paläontologischen Gesellschaft 16. bis 18. März 2007 in Freyburg/Unstrut », dans Jörg Erfurt, Lutz Christian Maul, Hallesches Jahrbuch für Geowissenschaften, vol. B 23, , p. 1–16
Liens externes
- (en) « Geiseltalsammlung an der Martin-Luther Universität Halle-Wittenberg » (consulté le )
- (en) « Geiseltalmuseum Halle » (consulté le )
Références
- Krumbiegel, Rüffle et Haubold 1983, p. 7–14
- (de) Joachim Wirth, Rudolf Eichner et Andreas Schroeter, « Revier Halle und Geiseltal », dans Gerhard H. Bachmann, Bodo-Carlo Ehling, Rudolf Eichner, Max Schwab, Geologie von Sachsen-Anhalt, Stuttgart, , p. 491–493
- Hellmund 2007
- (de) Karl-Heinz Radzinski, Bodo-Carlo Ehling, Reinhard Kunert et Gerhard Beutler, « Südöstliches Harzvorland », dans Gerhard H. Bachmann, Bodo-Carlo Ehling, Rudolf Eichner, Max Schwab, Geologie von Sachsen-Anhalt, Stuttgart, , p. 458–471
- Krumbiegel, Rüffle et Haubold 1983, p. 23
- (de) Matthias Thomae et Ivo Rappsilber, « Zur Entstehung der Geiseltalsenke », dans Harald Meller, Elefantenreich – Eine Fossilwelt in Europa, Halle/Saale, , p. 27–33
- (de) Matthias Thomae et Ivo Rappsilber, « Beitrag zur Klärung der Lagerungsverhältnisse des Quartärs im Tagebau Neumark-Nord », dans Dietrich Mania et al., Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen, Halle/Saale, coll. « Veröffentlichungen des Landesmuseums für Vorgeschichte, 62 », , p. 71–80
- (de) Horst Blumenstengel et Wilfried Krutzsch, « Tertiär », dans Gerhard H. Bachmann, Bodo-Carlo Ehling, Rudolf Eichner, Max Schwab, Geologie von Sachsen-Anhalt, Stuttgart, , p. 267–292
- Hartmut_Hauboldclass="lang-de"_lang="de">Matthias_Thomae1990">(de) Hartmut Haubold et Matthias Thomae, « Stratigraphische Revision der Wirbeltierfundstellen des Geiseltales », Hallesches Jahrbuch für Geowissenschaften, no 15, , p. 3–20
- Lothar_Eißmann1978">(de) Lothar Eißmann, « Mollisoldiapirismus », Zeitschrift für angewandte Geologie, vol. 24, , p. 130–138
- (de) Dietrich Mania, « Zur Geschichte des Geiseltals im Quartär », dans Harald Meller, Elefantenreich – Eine Fossilwelt in Europa, Halle/Saale, , p. 34–38
- (de) Dietrich Mania, « Quartärforschung im Tagebau Neumark-Nord, Geiseltal (Sachsen-Anhalt) und ihre bisherigen Ergebnisse », dans Dietrich Mania et al., Neumark-Nord: Ein interglaziales Ökosystem des mittelpaläolithischen Menschen, Halle/Saale, coll. « Veröffentlichungen des Landesmuseums für Vorgeschichte » (no 62), , p. 11–69
- Hartmut_Haubold1995">(de) Hartmut Haubold, « Wirbeltiergrabung- und forschung im Geiseltaleozän », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 1–18
- Hartmund_Haubold1989">(de) Hartmund Haubold, « Die Referenzfauna des Geiseltalium, MP Levels 11 bis 13 (Mitteleozän, Lutetium) », Palaeovertebrata, vol. 19, no 3, , p. 81–93
- Meinolf_Hellmund1997">(de) Meinolf Hellmund, « Letzte Grabungsaktivitäten im südwestlichen Geiseltal bei Halle (Sachsen-Anhalt, Deutschland) in den Jahren 1992 und 1993 », Hercynia, vol. N. F. 30, , p. 163–176
- _Hans_Gallwitz1955">(de) Hans Gallwitz, « Kalk, Kieselsäure und Schwefeleisen in der Braunkohle des Geiseltales und ihre Bedeutung für die Fossilisation », Paläontologische Zeitschrift, vol. 29, nos 1/2, , p. 33–37
- Ehrhard_Voigt1933">(de) Ehrhard Voigt, « Die Übertragung fossiler Wirbeltierleichen auf Zellulose-Filme, ein neue Bergungsmethode für Wirbeltiere aus der Braunkohle », Paläontologische Zeitschrift, vol. 15, , p. 72–78
- Günter_Krumbiegel1962">(de) Günter Krumbiegel, « Die Fossilfundstellen der mitteleozänen Braunkohle des Geiseltales », Wissenschaftliche Zeitschrift der Martin-Luther-Universität Halle-Wittenberg, Mathematisch-Naturwissenschaftliche Reihe , vol. 11, no 6, , p. 745–762
- Krumbiegel, Rüffle et Haubold 1983, p. 25–34
- Johannes_Weigelt1934">(en) Johannes Weigelt, « Die Geiseltalgrabungen des Jahres 1933 und die Biostratonomie der Fundschichten », Nova Acta Leopoldina, vol. N F 1, nos 4/5, , p. 552–600 (578)
- Ehrhard_Voigt1935">(de) Ehrhard Voigt, « Die Erhaltung von Epithelzellen mit Zellkernen, von Chromatophoren und Corium in fossiler Froschhaut aus der mitteleozänen Braunkohle des Geiseltales », Nova Acta Leopoldina, vol. N F 3, no 14, , p. 339–360
- (de) Wilfried Krutzsch, « Die Mikroflora des Geiseltales : IV: Die stratigraphische Stellung des Geiseltalprofils im Eozän und die sporenstratigraphische Untergliederung des mittleren Eozän », Abhandlungen des Zentralen Geologischen Institutes, Paläontologische Abhandlungen , vol. 26, , p. 47–92 (47, 51–57)
- Volker_Wilde2001">(de) Volker Wilde, « Ein Farnhorizont aus dem Mitteleozän des Geiseltales (Sachsen-Anhalt, Deutschland) », Hallesches Jahrbuch für Geowissenschaften , vol. BH 13, , p. 69–75
- Volker_Wilde1995">(de) Volker Wilde, « Die Makroflora aus dem Mitteleozän des Geiseltalgebietes, kurze Übersicht und Vergleiche », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 121–138
- L._Rüffle2008">(de) E. Kahlert et L. Rüffle, « Monokotylen in der mitteleozänen Braunkohle des Geiseltales bei Merseburg (Sachsen-Anhalt) und ihre ökologische Bedeutung », Feddes Repertorium, vol. 119, nos 3–4, , p. 217–224
- W._Salzmann1914">(de) W. Salzmann, « Das Braunkohlenvorkommen im Geiseltal mit besonderer Berücksichtigung der Genesis », Archiv für Lagerstättenforschung, vol. 17, , p. 1–105 (31–33)
- (de) Günter Krumbiegel, « Molluskenfunde (Gastropoda der Ordnung Pulmonata CUVIER 1795) in der mitteleozänen Braunkohle des Geiseltales und ihre fazielle und stratigraphische Bedeutung », Geologie, vol. 11, no 5, , p. 557–603
- (de) Günter Krumbiegel, « Ostracodenfunde im Tagebau Neumark-Süd (Geiseltal) », Geologie, vol. 11, no 3, , p. 334–353
- Karl_Beurlen1938">(de) Karl Beurlen, « Crustaceenreste aus der Geiseltalbraunkohle », Nova Acta Leopoldina, vol. N F 5, , p. 361–368
- Thomas_Hörneschmeyerclass="lang-de"_lang="de">Gert_Trösterclass="lang-de"_lang="de">Sonja_Weidmann1995">(de) Thomas Hörneschmeyer, Gert Tröster et Sonja Weidmann, « Die eozänen Käferfaunen des Geiseltales und der Grube Messel – ein Vergleich unter systematischen und paläoökologischen Gesichtspunkten », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 107–119
- Volker_Wildeclass="lang-de"_lang="de">Meinolf_Hellmund2006">(de) Volker Wilde et Meinolf Hellmund, « Neue Geländearbeiten im ehemaligen Braunkohlerevier Geiseltal », Natur und Museum, vol. 136, nos 7/8, , p. 162–173
- Ehrhard_Voigt1938">(de) Ehrhard Voigt, « Ein fossiler Saitenwurm (Gordius tenuifibrosus n. sp.) aus der eozänen Braunkohle des Geiseltales », Nova Acta Leopoldina, vol. N F 5, , p. 352–360
- (en) Jean Gaudant, « An attempt at the palaeontological history of the European mudminnows (Pisces, Teleostei, Umbridae) », Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, vol. 263, no 2, , p. 93–109
- Norbert_MicklichGaudant1989">(de) Norbert Micklich et Jean Gaudant, « Anthracoperca siebergi VOIGT 1934 (Pisces, Perciformes) – Ergebnisse einer Neuuntersuchung der mitteleozänen Barsche des Geiseltales », Zeitschrift für geologische Wissenschaften, vol. 17, no 5, , p. 503–521
- Krumbiegel, Rüffle et Haubold 1983, p. 114–118
- Michael_WuttkePřikrylYu._RatnikovDvořák2012">(en) Michael Wuttke, Tomáš Přikryl, Viacheslav Yu. Ratnikov et Zdeněk Dvořák, « Generic diversity and distributional dynamics of the Palaeobatrachidae (Amphibia: Anura) », Palaeobiology Palaeoenvironment, vol. 92, , p. 367–395
- Krumbiegel, Rüffle et Haubold 1983, p. 119–124
- Stefanie_Zimmermann-Rollius1967">(de) Stefanie Zimmermann-Rollius , « Beiträge zur Schildkrötenfauna der mitteleozänen Braunkohle des Geiseltales », Hercynia, vol. N F 4, , p. 83–104
- Günter_Krumbiegel1963">(de) Günter Krumbiegel, « Trionychidenfunde (Flußschildkröten) aus der Gattung Trionyx GEOFFROY 1809 aus der eozänen Braunkohle des Geiseltales », Geologie, vol. 12, , p. 196–223
- Johannes_Müller2001">(en) Johannes Müller , « Osteology and relationships of Eolacerta robusta, a lizard from the Middle Eocene of Germany (Reptilia, Squamata) », Journal of Vertebrate Paleontology , vol. 21, no 2, , p. 261–278
- Krister_T._Smith2009">(en) Krister T. Smith, « Eocene Lizards of the Clade Geiseltaliellus from Messel and Geiseltal, Germany, and the Early Radiation of Iguanidae (Reptilia: Squamata) », Bulletin of the Peabody Museum of Natural History , vol. 50, no 2, , p. 219–306 (256–257)
- (en) Olivier Rieppel, Jack L. Conrad et Jessica A. Maisano, « New morphological data for Eosaniwa koehni and revised phylogenetic analysis », Journal of Paleontology, vol. 81, no 4, , p. 760–769
- (en) Johannes Weigelt, « Die Wirbeltierausgrabungen im Geiseltal », Die Naturwissenschaften, vol. 21, , p. 321–325
- (de) Oskar Kuhn, « Die Schlangen (Boidae) aus dem Mitteleozän des Geiseltales », Nova Acta Leopoldina, vol. N F 7, , p. 119–133
- Krumbiegel, Rüffle et Haubold 1983, p. 142–145
- Michael_RauheOrtega2000">(en) Torsten Rossmann, Michael Rauhe et Francesco Ortega, « Studies on Cenozoic crocodiles: 8. Bergisuchus dietrichbergi KUHN (Sebecosuchia: Bergisuchidae n. fam.) from the Middle Eocene of Germany, some new systematic and biological conclusions », Paläontologische Zeitschrift, vol. 74, no 3, , p. 379–392
- Michael_Rauhe1995">(de) Michael Rauhe , « Die Lebensweise und Ökologie der Geiseltal-Krokodilier – Abschied von traditionellen Lehrmeinungen », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 65–80
- Rolf_Kohring1995">(de) Rolf Kohring, « Ultrastrukturen und Erhaltungsbedingungen fossiler Krokodil-Eierschalen aus dem Geiseltal (Mitteleozän) », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 93–105
- Karl-Heinz_Fischer1967">(de) Karl-Heinz Fischer, « Ein neuer großer Laufvogel aus dem Eozän des Geiseltales bei Halle (Saale) », Berichte der deutschen Gesellschaft für geologische Wissenschaften, vol. A 12, no 5, , p. 601–605
- (de) Kálmán Lambrecht, « Palaeotis Weigelti n. g. n. sp., eine fossile Trappe aus der mitteleozänen Braunkohle des Geiseltales », Jahrbuch des Halleschen Verbandes, vol. N F 7, , p. 20–29
- Karl-Heinz_Fischer1962">(de) Karl-Heinz Fischer, « Der Riesenlaufvogel Diatryma aus der eozänen Braunkohle des Geiseltales », Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte, vol. 4, , p. 26–33
- Meinolf_Hellmund2013">(en) Meinolf Hellmund, « Reappraisal of the bone inventory of Gastornis geiselensis (Fischer, 1978) from the Eocene “Geiseltal Fossillagerstätte” (Saxony-Anhalt, Germany) », Neues Jahrbuch für Geologie und Paläontologie Abhandlungen , vol. 269, no 2, , p. 203–220
- (de) « Harmloser Terrorvogel », sur Max-Planck-Gesellschaft , (consulté le ), Forschung/Aktuelles
- (de) Kálmán Lambrecht, « Drei neue Vogelformen aus dem Lutétium des Geiseltales », Nova Acta Leopoldina, vol. 3, , p. 361–367
- (en) Gerald Mayr, « Synonymy and actual affinities of the putative Middle Eocene « New World vulture » Eocathartes LAMBRECHT, 1935 and « hornbill » Geiseloceros LAMBRECHT, 1935 (Aves, Ameghinornithidae) », Paläontologische Zeitschrift, vol. 81, no 4, , p. 457–462
- Dennis_Stefan_Peters1998">(de) Dennis Stefan Peters, « Erstnachweis eines Seglers aus dem Geiseltal (Aves: Apodiformes) », Senckenbergiana lethaea, vol. 76, nos 1/2, , p. 211–212
- Florian_Heller1936">(de) Florian Heller, « Neue Beuteltierreste aus der mitteleozänen Braunkohle des Geiseltales bei Halle (Saale) », Nova Acta Leopoldina, vol. N F 4, , p. 311–316
- Gerhard_StorchHaubold1989">(en) Gerhard Storch et Hartmut Haubold, « Additionals to the Geiseltal Mammalian faunas, Middle Eocene: Didelphidae, Nyctitheriidae. Myrmecophagidae », Palaeovertebrata, vol. 19, no 3, , p. 95–114
- Gerhard_Storch1981">(de) Gerhard Storch , « Eurotamandua joresi, ein Myrmecophagide aus dem Eozän der « Grube Messel » bei Darmstadt (Mammalia, Xenarthra) », Senckenbergiana lethaea, vol. 61, nos 3/6, , p. 247–289 (253)
- Timothy_J._Gaudinclass="lang-en"_lang="en">Robert_J._Emryclass="lang-en"_lang="en">John_R._Wible2009">(en) Timothy J. Gaudin , Robert J. Emry et John R. Wible, « The Phylogeny of Living and Extinct Pangolins (Mammalia, Pholidota) and Associated Taxa: A Morphology Based Analysis », Journal of Mammal Evolution, vol. 16, , p. 235–305 (241, 274)
- Jerry_J._Hooker2013">(en) Jerry J. Hooker, « Origin and evolution of the Pseudorhyncocyonidae, a European Paleogene family of insectivorous placental mammals », Palaeontology, vol. 56, no 4, , p. 807–835 (818–820)
- Gerhard_Storch2001">Bernhard Sigé et Gerhard Storch, « Un nouveau Saturninia (Nyctitheriidae, Lipotyphla, Mammalia) de l'assise OK (Oberkohle, MP 14) du bassin lignitifère de Geiseltal (Eocène moyen supérieur d'Allemagne) », Senckenbergiana lethaea, vol. 81, no 2, , p. 343–346
- Gerhard_Storch1995">(de) Gerhard Storch, « Kleinsäugetiere aus dem Geiseltal und Messel im Kontext alttertiärer Faunenentfaltungen », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 59–64
- Urs_Thalmannclass="lang-de"_lang="de">Hartmut_HauboldDenis_Martin1989">(en) Urs Thalmann, Hartmut Haubold et Robert Denis Martin, « Pronycticebus neglectus – An almost complete adapid primate specimen from the Geiseltal (GDR) », Palaeovertebrata, vol. 19, no 3, , p. 115–130
- Jens_Lorenz_Franzen2000">(de) Jens Lorenz Franzen, « Der sechste Messel-Primate (Mammalia, Primates, Notharctidae, Cercamoniinae) », Senckenbergiana lethaea, vol. 80, no 1, , p. 289–303
- Jens_Lorenz_Franzenclass="lang-en"_lang="en">Philip_D._Gingerichclass="lang-de"_lang="de">Jörg_HabersetzerHurum2009">(en) Jens Lorenz Franzen , Philip D. Gingerich, Jörg Habersetzer, Jørn H. Hurum, Wighart von Koenigswald et B. Holly Smith, « Complete Primate Skeleton from the Middle Eocene of Messel in Germany: Morphology and Paleobiology », PLoS ONE, vol. 4, no 5, , e5723 (DOI 10.1371/journal.pone.0005723)
- Krumbiegel, Rüffle et Haubold 1983, p. 160–165
- (de) Horst Werner Matthes, « Ein neuer Creodontier: Prodissopsalis theriodis van Valen 1965 aus der eozänen Braunkohle des Geiseltales », Hallesches Jahrbuch für Mitteldeutsche Erdgeschichte , vol. 8, , p. 7–11
- Brigitte Lange-Bardi et Hellmund Haubold, « Les créodontes (Mammifères) du gisement du Geiseltal (Eocène Moyen, RDA) », Geobios, vol. 23, no 5, , p. 607–637
- (en) Leigh van Valen, « Some Europaea Proviverrini (Mammalia, Deltatheridida) », Palaeontology , vol. 8, no 4, , p. 638–665 (653–658)
- Krumbiegel, Rüffle et Haubold 1983, p. 165–172
- Jörg_ErfurtSudre1996">(de) Jörg Erfurt et Jean Sudre, « Eurodexeinae, eine neue Unterfamilie der Artiodactyla (Mammalia) aus dem Unter- und Mitteleozän Europas », Palaeovertebrata, vol. 25, no (2–4), , p. 371–390
- Jörg_Erfurtclass="lang-de"_lang="de">Hartmut_Haubold1989">(de) Jörg Erfurt et Hartmut Haubold, « Artiodactyla aus den eozänen Braunkohlen des Geiseltales bei Halle (DDR) », Palaeovertebrata, vol. 19, no 1, , p. 131–160 (152–155)
- Jörg_ErfurtSudre1995">Jörg Erfurt et Jean Sudre, « Un Haplobunodontidae nouveau, Hallebune krumbiegeli nov. gen. nov. sp. (Artiodactyla, Mammalia) dans L'Éocène Moyen du Geiseltal près Halle (Sachsen-Anhalt, Allemagne) », Palaeovertebrata, vol. 24, nos 1–2, , p. 84–99
- Jerry_J._Hookerclass="lang-en"_lang="en">Katherine_M._Thomas2001">(en) Jerry J. Hooker et Katherine M. Thomas, « New species of Amphirhagatherium (Choeropotamidae, Artiodactyla, Mammalia) from the late Eocene Headon Hill Formation of Southern England and phylogeny of endemic European Anthracotheroids », Palaeontology, vol. 44, no 5, , p. 827–853
- Jörg_Erfurt2000">(de) Jörg Erfurt, « Rekonstruktion des Skelettes und der Biologie von Anthracobunodon weigelti (Artiodactyla, Mammalia) aus dem Eozän des Geiseltales », Hallesches Jahrbuch für Geowissenschaften, vol. B, no 12, , p. 57–141, ici 59, 107
- Jörg_Erfurtclass="lang-de"_lang="de">Hans_Altner2003">(de) Jörg Erfurt et Hans Altner, « Habitus-Rekonstruktion von Anthracobunodon weigelti (Artiodactyla, Mammalia) aus dem Eozän des Geiseltales », dans Jan Michal Burdukiewicz, Lutz Fiedler, Wolf-Dieter Heinrich, Antje Justus, Enrico Brühl, Erkenntnisjäger. Festschrift für Dietrich Mania, Halle/Saale, Veröffentlichungen des Landesmuseums für Vorgeschichte in Halle, 57, , p. 153–176
- Jörg_Erfurt1995">(de) Jörg Erfurt, « Taxonomie der eozänen Artiodactyla (Mammalia) des Geiseltales mit besonderer Berücksichtigung der Gattung Rhagatherium », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 47–58
- (de) Horst Werner Matthes, « Weitere eozäne Equiden aus dem Geiseltal », Zeitschrift für Geologische Wissenschaften, vol. 7, no 6, , p. 783–805
- Jens_Lorenz_FranzenHaubold1986">(de) Jens Lorenz Franzen et Hartmut Haubold, « Revision der Equoidea aus den eozänen Braunkohlen des Geiseltales bei Halle (DDR) », Palaeovertebrata, vol. 16, no 1, , p. 1–34
- Jens_Lorenz_Franzen2006">(en) Jens Lorenz Franzen, « Eurohippus n. g., a new genus of horses from the Middle to Late Eocene of Europe », Senckenbergiana lethaea, vol. 86, no 1, , p. 97–102
- _Meinolf_Hellmund2000">(de) Meinolf Hellmund, « Erstnachweis von Plagiolophus cartieri Stehlin (Palaeotheriidae, Perissodactyla) in der Unteren Mittelkohle (uMK, MP 12) des Geiseltales bei Halle (Sachsen-Anhalt, Deutschland) », Neues Jahrbuch für Geologie und Paläontologie, no 4, , p. 205–216
- Karl-Heinz-Fischer1964">(de) Karl-Heinz-Fischer, « Die tapiroiden Perissodactylen aus der eozänen Braunkohle des Geiseltales », Geologie, vol. 45, , p. 1–101 (71)
- Karl-Heinz-Fischer1977">(de) Karl-Heinz-Fischer, « Neue Funde von Rhinocerolophiodon (n. gen.), Lophiodon, und Hyrachyus (Ceratomorpha, Perissodactyla, Mammalia) aus dem Eozän des Geiseltals bei Halle (DDR) : 1. Teil: Rhinocerolophiodon », Zeitschrift für geologische Wissenschaft, vol. 5, , p. 909–919
- (de) Karl-Heinz-Fischer, « Neue Funde von Rhinocerolophiodon (n. gen.), Lophiodon, und Hyrachyus (Ceratomorpha, Perissodactyla, Mammalia) aus dem Eozän des Geiseltals bei Halle (DDR) : 2. Teil: Lophiodon. », Zeitschrift für geologische Wissenschaft, vol. 5, , p. 1129–1152
- Karl-Heinz-Fischer1967">(de) Karl-Heinz-Fischer, « Zur systematischen Stellung von Chasmotherium RÜTIMEYER 1862 (Mammalia, Perissodactyla) », Berichte der deutschen Gesellschaft für geologische Wissenschaften, vol. 12A, no 5, , p. 595–600
- (de) Kerstin Hlawatsch et Jörg Erfurt, « Zahnmorphologie und stratigraphische Verbreitung von Hyrachyus minimus (Perissodactyla, Mammalia) in den eozänen Geiseltalschichten : 34. Tagung des Arbeitskreises für Wirbeltierpaläontologie der Paläontologischen Gesellschaft 16. bis 18. März 2007 in Freyburg/Unstrut », dans Jörg Erfurt, Lutz Christian Maul, Hallesches Jahrbuch für Geowissenschaften, BH 23, , p. 161–173
- Erhard_Voigt1937">(de) Erhard Voigt, « Weichteile an Fischen, Amphibien und Reptilien aus der eozänen Braunkohle des Geiseltales », Nova Acta Leopoldina, vol. N F 5, , p. 115–142
- Adolf_Brachofen-Echt1936">(de) Adolf Brachofen-Echt, « Das Vorkommen von Federn in der eozänen Braunkohle des Geiseltales », Nova Acta Leopoldina, vol. N F 4, , p. 335–340
- Erhard_Voigt1936">(de) Erhard Voigt, « Über das Haarkleid einiger Säugetieren aus der eozänen Braunkohle des Geiseltales », Nova Acta Leopoldina, vol. N F 4, , p. 317–334
- (de) Erhard Voigt, « Weichteile an Säugetieren aus der eozänen Braunkohle des Geiseltales », Nova Acta Leopoldina, vol. N F 4, , p. 301–310
- Krumbiegel, Rüffle et Haubold 1983, p. 189–193
- Angelika_Ottoclass="lang-de"_lang="de">Bernd_R._T._Simoneitclass="lang-de"_lang="de">Volker_Wilde2001">(en) Angelika Otto, Bernd R. T. Simoneit et Volker Wilde, « Initial results on the biomarker composition and „Affenhaar“ from Middle Eocene lignites of the Geiseltal (Sachsen-Anhalt, Germany) », Hallesches Jahrbuch für Geowissenschaften, vol. BH 13, , p. 57–68
- Johannes_Weigelt1935">(de) Johannes Weigelt, « Lophiodon in der oberen Kohle des Geiseltales », Nova Acta Leopoldina, vol. N F 3, no 14, , p. 369–402 (376, 384–385)
- Volker_Wildeclass="lang-de"_lang="de">_Meinolf_Hellmund2010">(en) Volker Wilde et Meinolf Hellmund, « First record of gut contents from a middle Eocene equid from the Geiseltal near Halle (Saale), Sachsen-Anhalt, Central Germany », Palaeobiodiversity and Palaeoenvironments, vol. 90, , p. 153–162
- Henry_Schroeder1913">(de) Henry Schroeder, « Das Vorkommen der Gattung Lophiodon in der Braunkohle Sachsens », Centralblatt für Mineralogie, Geologie und Paläontologie, vol. 11, , p. 351
- Jens_Lorenz_Franzen2005">(de) Jens Lorenz Franzen, « Warum Geiseltalium? », Courier des Forschungsinstitutes Senckenberg, vol. 255, , p. 77–79
- Dieter_F._Mertzclass="lang-de"_lang="de">Paul_R._Renne2005">(en) Dieter F. Mertz et Paul R. Renne, « A numerical age for the Messel fossil deposit (UNESCO World Heritage Site) derived from 40Ar/39Ar dating on a basaltic rock fragment », Courier des Forschungsinstitutes Senckenberg, vol. 255, , p. 67–75
- Dieter_F._MertzSwisher_IIIclass="lang-de"_lang="de">Jens_Lorenz_Franzenclass="lang-de"_lang="de">_Franz-Otto_Neuffer2000">(en) Dieter F. Mertz, Carl C. Swisher III, Jens Lorenz Franzen, Franz-Otto Neuffer et Herbert Lutz , « Numerical dating of the Eckfeld maar fossil site, Eifel, Germany: calibration mark for the Eocene time scale », Naturwissenschaften, vol. 8, , p. 270–274
- Horst_Blumenstengel2001">(de) Horst Blumenstengel, « Palynologische Untersuchungen tertiärer Ablagerungen aus dem ehemaligen Braunkohletagebau Mücheln-Westfeld (Geiseltal, Sachsen-Anhalt, Deutschland) », Hallesches Jahrbuch für Geowissenschaften, vol. BH 13, , p. 31–39
- Krumbiegel, Rüffle et Haubold 1983, p. 41–43
- Krumbiegel, Rüffle et Haubold 1983, p. 34–37
- Johannes_Weigelt1939">(de) Johannes Weigelt , « Die Aufdeckung der bisher ältesten tertiären Säugetierfauna Deutschlands », Nova Acta Leopoldina, vol. N F 7, , p. 515–528
- Kenneth_D._Roseclass="lang-de"_lang="de">Gerhard_Storchclass="lang-de"_lang="de">Katrin_Krohmann2013">(en) Kenneth D. Rose , Gerhard Storch et Katrin Krohmann , « Small-mammal postcrania from the middle Paleocene of Walbeck, Germany », Paläontologische Zeitschrift, (DOI 10.1007/s12542-013-0211-3)
- Kenneth_D._Rose2012">(en) Kenneth D. Rose, « The importance of Messel for interpreting Eocene Holarctic mammalian faunas », Palaeobiology Palaeoenvironment , vol. 92, , p. 631–647 (DOI 10.1007/s12549-012-0090-8)
- Herbert_Frankenhäuserclass="lang-de"_lang="de">Werner_Löhnertzclass="lang-de"_lang="de">Jens_L._Franzenclass="lang-de"_lang="de">Uwe_Kaufluss2009">(de) Herbert Frankenhäuser, Werner Löhnertz, Jens L. Franzen, Uwe Kaufluss, Martin Koziol, Herbert Lutz, Dieter F. Mertz, Jens Mingram, Torsten Wappler et Volker Wilde, « Das Eckfelder Maar in der Vulkaneifel – Fenster in einen küstenfernen Lebensraum vor 44 Millionen Jahren », Mainzer Naturwissenschaftliches Archiv, vol. 47, , p. 263–324 (301–303)
- (de) Geiseltal : Mitteldeutsches Braunkohlenrevier 3. Senftenberg , LMBV,
- (de) Hartmut Haubold, « 60 Jahre Geiseltalmuseum an der Martin-Luther-Universität Halle-Wittenberg », Hallesches Jahrbuch für Geowissenschaften, vol. B 17, , p. 19–25
- (de) « Geiseltalmuseum schließt – Fossilien ab 2015 im Naturkundlichen Universitätsmuseum », sur Communiqué de presse 237/2011 de l'université Martin-Luther de Halle-Wittenberg, (consulté le ) (de) « Version pdf » (consulté le )