Boverisuchus
Weigeltisuchus

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Reptilia |
| Ordre | Crocodilia |
| Famille | † Planocraniidae |
Espèces de rang inférieur
- † B. magnifrons Kuhn, 1938 espèce type
- † B. vorax Troxell, 1925
- Crocodylus vorax Troxell, 1925
- Pristichampsus vorax (Troxell, 1925) Langston, 1975
- Weigeltisuchus Kuhn, 1938
- Weigeltisuchus geiseltalensis Kuhn, 1938
Boverisuchus est un genre fossile de reptiles carnivores terrestres. Cet animal était soit membre, soit proche parent, du clade des crocodiliens, qui est défini comme le groupe-couronne des crocodiles actuels, autrement dit comme l'ensemble des descendants, actuels ou fossiles, du dernier ancêtre commun des crocodiles actuels. Il a vécu en Europe et en Amérique du Nord durant le Paléogène, de 61,6 à 41,3 millions d'années avant le présent, première période du Cénozoïque (autrefois appelée ère Tertiaire)[1] - [2].
Les premiers fragments fossiles liés à ce genre ont été découverts à Argenton-sur-Creuse, dans l'Indre (centre de la France) et étudiés en 1820 par le naturaliste Georges Cuvier. Un chapitre est consacré à ce Crocodile des marnières d'Argenton dans son ouvrage Recherches sur les ossements fossiles de quadrupèdes (1812). Par la suite, des spécimens proches ont été découverts en Italie, aux États-Unis, en Allemagne et en Chine. Leur attribution a changé plusieurs fois.
Alors que ses ancêtres avaient un mode de vie aquatique similaire à celui des crocodiles actuels, Boverisuchus a évolué vers un mode de prédation terrestre. Ses dents aplaties et en forme de scie (similaires à celles des dinosaures théropodes ou de certains varans), son armure d'ostéodermes très complète et ses griffes en forme de sabots prouvent cette spécialisation. Il pourrait avoir été facultativement bipède, mais ce point reste incertain. Ses principales proies sont probablement de petits mammifères, comme les ongulés présents à l'époque, ainsi que des oiseaux terrestres.
Historique de l'étude des fossiles
Le « crocodile de marnières d'Argenton » de G. Cuvier
En , Georges Cuvier décrit, dans ses Recherches sur les ossements fossiles de quadrupèdes, le Crocodile des marnières d'Argenton, sur la base de quelques fragments fossiles découverts dans l'Indre, dans le centre de la France.
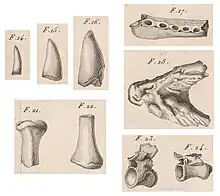
Il remarque la forme particulière des dents, qui suggère un comportement de prédation terrestre, et fait le rapprochement avec la denture des grands varans. Le Britannique George Robert Gray décrit, sur la base des schémas de Cuvier, l'espèce Crocodilus rollinati, transférée en 1853 par le zoologiste français Paul Gervais vers le nouveau genre Pristichampsus[3].

Le Crocodilus de Monte Bolca de F. Sacco
En , Federico Sacco (it) publie la description de Crocodilus bolcensis, du nom du gisement de fossiles où il a été découvert, Monte Bolca dans la province de Vérone. Le spécimen en question, qui est aujourd'hui visible au Museo Regionale di Scienze Naturali di Torino, pourrait être rattaché au genre Boverisuchus, mais cette identification reste incertaine[4].
Spécimens supplémentaires découverts au XXe siècle

Le genre Boverisuchus et l'espèce Boverisuchus vorax sont décrits en 1925 sur la base de fossiles trouvés dans le Wyoming. Un fossile type, découvert dans le gisement de fossiles du Geiseltal (Saxe-Anhalt, Allemagne), est utilisé pour définir Weigeltisuchus geiseltalensis en [3]. Dans les années 1960, il est établi que Boverisuchus, Weigeltisuchus et Pristichampsus sont synonymes. Conformément aux règles de la nomenclature, l'usage du terme publié en premier, Pristichampsus, est alors préféré[3].
Deux autres espèces, Pristichampsus birjukovi et Pristichampsus kuznetzovi, sont proposées en sur la base de fossiles trouvés au Kazakhstan. Néanmoins, les fossiles sont très fragmentaires et, selon le paléontologue américain Christopher Andrew Brochu, professeur associé à l'université de l'Iowa et spécialiste des suchiens fossiles[5], ces spécimens devraient être réexaminés[3].
Genre
En , Brochu établit que le fossile étudié par Cuvier, à partir duquel Pristichampsus a été défini, est trop fragmentaire pour servir de fossile-type. Depuis lors, le nom Boverisuchus est à nouveau valide pour désigner le genre[6].
Les spécimens découverts en Amérique du Nord ont un crâne plus large que ceux des spécimens allemands, mais cette différence n'étant pas plus importante que les variations observées à l'intérieur d'une même espèce chez des crocodiliens actuels, la distinction entre les deux espèces pourrait être contestée. Par ailleurs, si le spécimen étudié par Sacco appartient à la même espèce que les spécimens allemands, le nom Bolcensis étant antérieur, la désignation Boverisuchus magnifrons serait alors appelée à être remplacée par Boverisuchus bolcensis[3].
Caractéristiques

Aucun fossile connu ne permet d'établir la transition entre ses ancêtres aquatiques et Boverisuchus, forme terrestre. Le caïman de Schneider, un crocodilien (mais d'un autre genre, Paleosuchus et appartenant à une espèce actuelle) tend lui aussi vers un mode de vie plus terrestre, ce qui pourrait fournir une analogie. La forme assez plate de ses dents, sa capacité à tenir la tête plus haute que la majorité des crocodiliens, la forme de son nez et de ses orbites représentent tous des parallèles avec les traits de Boverisuchus[3].
Dimensions
L'animal adulte avait une longueur estimée de 2,20 à 3,60 m, pour une masse corporelle moyenne de l'ordre de 100 kg[7].
Crâne et mâchoire

Les crocodiles aquatiques possèdent des dents de forme conique, adaptées à leur mode de prédation : ils happent une proie dans leur mâchoire et l'entraînent sous l'eau. Ils ne peuvent guère mâcher leur victime, et en avalent d'énormes pièces[8]. À contrario, Boverisuchus possède des dents de forme aplatie et présentant des dentelures, ce qui correspond à un prédateur terrestre : il tue ses victimes en les saignant et a une meilleure capacité à mâcher. Ce trait, appelé ziphodontie, est d'ailleurs récurrent dans l'évolution de nombreux clades de prédateurs terrestres, comme d'autres suchiens terrestres (dont Notosuchia) et les dinosaures théropodes[3].

Les dents de Boverisuchus et de ses parents sont remarquablement similaires à celles des dinosaures théropodes, au point que des dents isolées, découvertes dans des strates éocènes, ont parfois été, à tort, interprétées comme des preuves de la survie de certains dinosaures non-aviens après l'extinction Crétacé-Paléogène[3].
Protection
Alors que les crocodiles aquatiques possèdent des ostéodermes essentiellement sur le dos, chez Boverisuchus cette protection est beaucoup plus complète : elle s'étend sur les flancs et le ventre. Il s'agit là encore d'un trait cohérent avec l'adaptation au mode de vie terrestre, des combats sur la terre ferme demandant une protection plus complète[9].
Locomotion
Boverisuchus est adapté à la course terrestre. Seuls trois doigts de chaque patte touchent le sol ; les griffes de doigts avaient pris une forme se rapprochant de celle de sabots. Le terme de « crocodiles à sabots » est d'ailleurs utilisé de façon informelle pour désigner ces animaux. Les pattes sont très droites et très hautes pour un crocodilien. Elles sont assez similaires à celles des lointains ancêtres des chevaux, comme Mesohippus. Les sabots supportent tous le poids de l'animal et lui permettent de galoper. Cependant, le galop n'est peut-être pas sa seule méthode de locomotion : les pattes arrière étant plus développées que ses pattes avant, il a peut-être une aptitude à la course bipède, à l'image du basilic. Ce point reste cependant très incertain : certains spécialistes pensent que le centre de gravité du corps de l'animal se situe bien trop en avant pour permettre la bipédie[3].
Inversement, Boverisuchus est moins bien adapté à la locomotion aquatique que les crocodiles actuels. Sa queue est de section assez circulaire (ressemblant à celle d'un dinosaure), et non aplatie verticalement, ce qui la rend moins efficace pour la propulsion dans l'eau. De plus, elle manque probablement de souplesse pour cela, du fait de la disposition des ostéodermes qui protègent les flancs de l'animal. La forme de son crâne n'est pas particulièrement hydrodynamique[9].
Histoire évolutive
Les changements d'habitat sont récurrents dans l'évolution des crocodylomorphes. Les premiers d'entre eux, à la fin du Trias, sont des reptiles terrestres. Une ramification, Thalattosuchia, évolue vers un mode de vie marin, tandis que Neosuchia occupe l'habitat d'eau douce encore associé à ses représentants actuels. Mais, parmi les Neosuchia, certains clades adoptent à leur tour un habitat marin (Pholidosauridae, Dyrosauridae), alors que les Planocraniidae (dont Boverisuchus) et les Quinkana deviennent des animaux terrestres[10].
La première moitié de l'Éocène est la période la plus chaude du Cénozoïque, et même probablement la plus chaude depuis le Trias. La plupart des terres émergées sont occupées à cette époque par des écosystèmes similaires aux actuelles forêts tropicales, même en Antarctique, sous une paléolattitude de 70 degrés sud. Un tel environnement est favorable aux espèces ectothermes[11] - [12].
À la suite de l'extinction Crétacé-Paléogène, de très nombreuses niches écologiques se sont retrouvées vacantes, avec l'extinction non seulement des dinosaures non aviens, mais aussi, plus proches de Boverisuchus, de la plupart des membres du sous-ordre des Notosuchia, un groupe très diversifié de crocodyliformes terrestres. En particulier, il y a, au début du Cénozoïque, assez peu de grands carnivores terrestres dans l'hémisphère nord. En Amérique du Sud, les Sebecidae, représentants survivants de Notosuchia, sont les prédateurs dominants. En revanche, nombre de crocodiliens se disputent les proies disponibles dans les zones humides. Boverisuchus et les genres apparentés représentent une adaptation d'un crocodilien (ou d'un très proche parent des crocodiliens) pour occuper la niche écologique des milieux terrestres de l'hémisphère nord, alors vacante[13][14].
Les premiers membres connus de la famille des Planocraniidae se trouvent en Chine pendant le Paléocène, époque précédant l'Éocène. Les Planocraniidae semblent s'être répandus de l'Asie vers l'Europe et vers l'Amérique du Nord, ou de l'Asie vers l'Europe par l'Amérique du Nord[15]. En effet, au début de l'Éocène, il existe encore une connexion terrestre entre l'Europe et l'Amérique du Nord, par le Groenland, et le climat est si chaud que la migration d'espèces à sang froid est possible via cette bande de terre émergée[16].
Les raisons de l’extinction du genre Boverisuchus ne sont pas connues, mais celle-ci s’inscrit dans un contexte plus large : en effet, à la fin de l'Éocène, le climat s'est fortement refroidi. Ce refroidissement a longtemps été attribué à la capture de CO2 par les réactions chimiques avec des minéraux mis à disposition par l'orogenèse Himalayenne, mais, en , cette interprétation est remise en cause[17]. Ce refroidissement crée des conditions beaucoup moins favorables aux animaux ectothermes. En conséquence, la diversité des suchiens diminue, et leur extension géographique se réduit vers la zone intertropicale, les mammifères dominant de plus en plus dans les régions de latitude moyenne[3].
Classification
Famille
L'espèce Planocrania Datagensis, espèce-type du genre Planocrania a été décrite en par Li Jinling sur la base d'un fossile trouvé dans le Guangdong. Ce spécimen est plus ancien (Paléocène) et son adaptation à la vie terrestre est moins complète. Ses dents sont aplaties, mais ne sont pas dentelées comme celles de Boverisuchus. Le genre Planocrania a été regroupé avec Pristichampsus (renommé Boverisuchus depuis) dans la même famille, celle des Planocraniidae[18]. Un troisième genre a aussi été rattaché à cette famille : il s'agit de Duerosuchus, connu uniquement par un crâne découvert à Corrales del Vino, en Espagne[19].
Relation avec l'ordre des Crocodilia
.jpg.webp)
Le clade des Crocodilia est défini comme le groupe-couronne des crocodiles actuels (Gavial du Gange, Crocodile du Nil, Alligators…)[20]. L'appartenance ou non des Planocraniidae, dont Boverisuchus, à ce clade, reste incertaine. Une étude phylogénétique britannique publiée en [21] place les Planocraniidae dans le clade des Crocodilia, comme groupe frère des longirostres (groupe formé de gavialoidea et crocodyloidea), tout en citant des études antérieures qui, utilisant des méthodes de comparaison anatomiques légèrement différentes, placent les Planocraniidae comme divergeant légèrement avant le dernier ancêtre commun des Crocodilia, donc hors de ce clade. Il n'y a pas de consensus scientifique sur la question[21].
| Cladogramme publié en 2018[22] | Cladogramme publié en 2021[21] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Paléobiologie


Les proies de Boverisuchus sont probablement principalement des petits ongulés comme les Lophiodon et Propalaeotherium, très présents en Europe pendant cette période. Il pouvait aussi s'attaquer à des oiseaux terrestres comme les Palaeotis et de jeunes spécimens de Gastornis, ainsi qu'à des poissons et des reptiles [23].
Le mode de prédation est probablement basé sur l'affût, à l'image du dragon de Komodo, prédateur terrestre ectotherme moderne de taille similaire[3].
Les autres crocodiliens présents dans les mêmes gisements fossiles en Allemagne sont les genres Asiatosuchus, Allognathosuchus et Diplocynodon. Les différences de taille (Asiatosuchus est nettement plus grand que Boverisuchus, tandis que Allognathosuchus et Diplocynodon sont de taille plus modeste), et de mode de vie, indiquent que ces animaux ne sont pas en compétition trop directe pour les mêmes proies[23].
Références
- (en) F.M. Gradstein, J.G Ogg, M. Schmitz et G. Ogg, The Geologic Time Scale 2012, Elsevier, , 1176 p. (ISBN 978-0-444-59448-8, lire en ligne).
- « Charte stratigraphique internationale (2012) » [PDF], sur http://www.stratigraphy.org/ (consulté le ).
- Brochu 2012, p. 521–550.
- Simone Matteo Seghetti, Georgios L. Georgalis, Emanuel Tschopp et Massimo Delfino, « A historical overview of the reptile fauna from the Eocene Bolca Fossil-Lagerstätte (Italy) », Bollettino della Società Paleontologica Italiana, vol. 61, no 2, , p. 119–143 (ISSN 0375-7633, DOI 10.4435/BSPI.2022.14, lire en ligne, consulté le )
- (en) « Christopher Brochu, Ph.D. | Environmental Sciences - The University of Iowa », sur environmentalsciences.uiowa.edu (consulté le )
- (en) Xiao-Chun Wu et Donald B. Brinkman, « A new crocodylian (Eusuchia) from the uppermost Cretaceous of Alberta, Canada », Canadian Journal of Earth Sciences, vol. 52, no 8, , p. 590–607 (ISSN 0008-4077 et 1480-3313, DOI 10.1139/cjes-2014-0133, lire en ligne, consulté le )
- (en) Masaya Iijima et Tai Kubo, « Vertebrae-Based Body Length Estimation in Crocodylians and Its Implication for Sexual Maturity and the Maximum Sizes », Integrative Organismal Biology, vol. 2, no 1, (ISSN 2517-4843, PMID 33791579, PMCID PMC7891683, DOI 10.1093/iob/obaa042, lire en ligne, consulté le )
- (en-US) Geology Page, « Tiny new fossil crocodile-relative had mammal-like teeth », sur Geology Page, (consulté le )
- (en) Torsten Rossmann, « Studies on Cenozoic crocodiles: 5. Biomechanical investigation on the postcranial skeleton of the Palaeogene crocodile Pristichampsus rollinatii (Eusuchia: Pristichampsidae) », Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, vol. 217, no 3, , p. 289–330 (ISSN 0077-7749, DOI 10.1127/njgpa/217/2000/289, lire en ligne, consulté le )
- (en) Eric W. Wilberg, Alan H. Turner et Christopher A. Brochu, « Evolutionary structure and timing of major habitat shifts in Crocodylomorpha », Scientific Reports, vol. 9, no 1, (ISSN 2045-2322, PMID 30679529, PMCID PMC6346023, DOI 10.1038/s41598-018-36795-1, lire en ligne, consulté le )
- (en) Integrated Ocean Drilling Program Expedition 318 Scientists, Jörg Pross, Lineth Contreras et Peter K. Bijl, « Persistent near-tropical warmth on the Antarctic continent during the early Eocene epoch », Nature, vol. 488, no 7409, , p. 73–77 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature11300, lire en ligne, consulté le )
- (en) Margot J. Cramwinckel, Matthew Huber, Ilja J. Kocken et Claudia Agnini, « Synchronous tropical and polar temperature evolution in the Eocene », Nature, vol. 559, no 7714, , p. 382–386 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/s41586-018-0272-2, lire en ligne, consulté le )
- Hastings et Hellmund 2017, p. 119–146.
- (en) Eduardo Puértolas-Pascual, Alejandro Blanco, Christopher A. Brochu et José Ignacio Canudo, « Review of the Late Cretaceous-early Paleogene crocodylomorphs of Europe: Extinction patterns across the K-PG boundary », Cretaceous Research, vol. 57, , p. 565–590 (DOI 10.1016/j.cretres.2015.08.002, lire en ligne, consulté le )
- (en) T Kotsakis, M Delfino et P Piras, « Italian Cenozoic crocodilians: taxa, timing and palaeobiogeographic implications », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 210, no 1, , p. 67–87 (DOI 10.1016/j.palaeo.2004.03.013, lire en ligne, consulté le )
- (en) Bruce H. Tiffney, « The Eocene North Atlantic Land Bridge: Its Importance in Tertiary and Modern Phytogeography of the Northern Hemisphere », Journal of the Arnold Arboretum, vol. 66, no 2, , p. 243–273 (ISSN 0004-2625, lire en ligne, consulté le )
- (en) Peter D. Clift et Tara N. Jonell, « Himalayan‐Tibetan Erosion Is Not the Cause of Neogene Global Cooling », Geophysical Research Letters, vol. 48, no 8, (ISSN 0094-8276 et 1944-8007, DOI 10.1029/2020GL087742, lire en ligne, consulté le )
- D.M. Martill, « Fake Fossils », dans Encyclopedia of Geology, Elsevier, (lire en ligne), p. 169–173
- (en) Iván Narváez, Ane De Celis, Fernando Escaso et Santiago Martín De Jesús, « Redescription and phylogenetic placement of the Spanish middle Eocene eusuchian Duerosuchus piscator (Crocodylia, Planocraniidae) », Journal of Vertebrate Paleontology, vol. 41, no 3, (ISSN 0272-4634 et 1937-2809, DOI 10.1080/02724634.2021.1974868, lire en ligne, consulté le )
- « Order Crocodilia », dans The Reptile Ear, Princeton University Press, (lire en ligne), p. 923–964
- (en) Jonathan P. Rio et Philip D. Mannion, « Phylogenetic analysis of a new morphological dataset elucidates the evolutionary history of Crocodylia and resolves the long-standing gharial problem », PeerJ, vol. 9, , e12094 (ISSN 2167-8359, PMID 34567843, PMCID PMC8428266, DOI 10.7717/peerj.12094, lire en ligne, consulté le )
- (en) Michael S. Y. Lee et Adam M. Yates, « Tip-dating and homoplasy: reconciling the shallow molecular divergences of modern gharials with their long fossil record », Proceedings of the Royal Society B: Biological Sciences, vol. 285, no 1881, , p. 20181071 (ISSN 0962-8452 et 1471-2954, PMID 30051855, PMCID PMC6030529, DOI 10.1098/rspb.2018.1071, lire en ligne, consulté le )
- (en) Alexander K. Hastings et Meinolf Hellmund, « Evidence for prey preference partitioning in the middle Eocene high-diversity crocodylian assemblage of the Geiseltal-Fossillagerstätte, Germany utilizing skull shape analysis », Geological Magazine, vol. 154, no 1, , p. 119–146 (ISSN 0016-7568 et 1469-5081, DOI 10.1017/S0016756815001041, lire en ligne, consulté le )
Voir aussi
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- (en) Alexander K. Hastings et Meinolf Hellmund, « Evidence for prey preference partitioning in the middle Eocene high-diversity crocodylian assemblage of the Geiseltal-Fossillagerstätte, Germany utilizing skull shape analysis », Geological Magazine, vol. 154, no 1, , p. 119–146 (ISSN 0016-7568 et 1469-5081, DOI 10.1017/S0016756815001041, lire en ligne, consulté le ).

- (en) Christopher A. Brochu, « Phylogenetic relationships of Palaeogene ziphodont eusuchians and the status of Pristichampsus Gervais, 1853 », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 103, nos 3-4, , p. 521–550 (ISSN 1755-6910 et 1755-6929, DOI 10.1017/S1755691013000200, lire en ligne, consulté le ).
- Georges Cuvier, Recherches sur les ossements fossiles de quadrupèdes, où l'on rétablit les caractères de plusieurs espèces d'animaux que les révolutions du globe paraissent avoir détruites (4 volumes, 1812). Textes en ligne : volume 1, volume 2 lire en ligne sur Gallica, volume 3 lire en ligne sur Gallica, volume 4 lire en ligne sur Gallica.
Médiagraphie
Articles connexes
- Autres crocodylomorphes terrestres : Notosuchia, Mekosuchus et l'article Eusuchia
- Grande Coupure
Liens externes
- Ressources relatives au vivant :