Dyrosauridae
Les Dyrosauridae sont une famille éteinte et fossile de crocodyliformes (un taxon regroupant bon nombre de groupes, dont seul subsiste le groupe des crocodiliens actuels), apparus il y a probablement un peu plus de 113 millions d'années (Crétacé inférieur) et qui se sont éteints il y a 38 millions d'années. C'est donc uniquement par l'intermédiaire des fossiles que nous connaissons ce groupe.
Stefano, 1903
- † Archosauria
- † Crurotarsi
- † Crocodyliformes
- † Dyrosauridae
- † « Phosphatosaurinae »
- † « Hyposaurinae »
- † Dyrosauridae
- † Crocodyliformes
- † Crurotarsi
Description
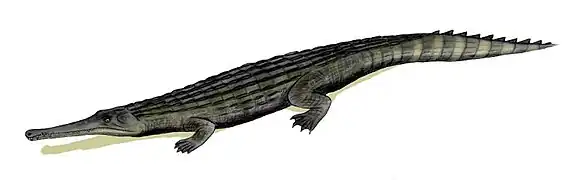
C'est un groupe (à une ou deux exceptions près) de crocodiles marins, bien adaptés à la vie aquatique (leur queue, principal agent de locomotion chez les crocodiles, est très haute, permettant une nage active et efficace), mais ils sont également très à l'aise sur la terre ferme grâce à leurs membres robustes.
Les particularités anatomiques communes à toutes les espèces de ce groupe sont assez nombreuses :
- museau généralement très allongé (marquant un régime alimentaire essentiellement piscivore) ;
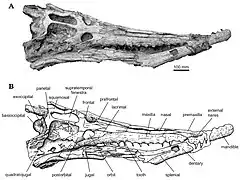
- fenêtres supratemporales (trous à l'arrière des orbites, servant à l'insertion des muscles de la mâchoire sur le crâne) très grandes et allongées ;
- septième dent toujours réduite et proche de la huitième.
Il y a 65 millions d’années, la fin du Crétacé est marquée par la célèbre disparition des dinosaures sur la terre ferme, mais aussi par la disparition des mosasaures et des plésiosaures dans le milieu marin. Les Dyrosauridae, quant à eux, traversent sans encombre la grande crise biologique, et il semble même que les Dyrosauridae, occupant des niches écologiques similaires à celles occupées par les mosasaures et plésiosaures, aient profité de la disparition de ces derniers pour coloniser le milieu marin, où ils ont largement prospéré, avant de régresser fortement à leur tour il y a 40 millions d’années, peut-être à cause de la concurrence créée par l’émergence des premiers mammifères marins, avant de s'éteindre complètement. Aujourd’hui, les crocodiles (les cousins des Dyrosauridae) sont des animaux fluviatiles, deux espèces (le Crocodile marin et le Crocodile du Nil, très éloignés des dyrosaures) fréquentent le milieu marin et une seule se nourrit en mer (le Crocodile marin).
Taxonomie
La famille des Dyrosauridae a été créée par Stefano (1903) pour l’espèce tunisienne Dyrosaurus phosphaticus ; cette espèce, décrite par Thomas (1893) sous le nom de Crocodilus phosphaticus sur la base de quelques dents et fragments postcrâniens, fut considérée comme un rhynchocéphale par Stefano (1903) et comme un champsosaure par Pomel (1894a). Ce dernier décrivit une seconde espèce en Algérie, D. thevestensis (Pomel, 1894b) avant de la synonymiser avec D. phosphaticus (Pomel, 1894a).
En fait, les premiers restes appartenant à cette famille furent d’abord décrits par Owen(1849) avec Hyposaurus rogersii, qui considérait cette espèce du Maastrichtien et Paleocène du New Jersey comme un « Teleosauroid ». Sauvage (1904), Nopsca (1905) et Thévenin (1911a, b) quant à eux, classaient les Dyrosauridae dans le groupe des « mésosuchiens ».
La validité de cette famille fut souvent discutée, Thévenin (1911a, b), Dollo (1914) et Abel (1919) considérant Dyrosaurus comme un Teleosauridae, Piveteau (1935), Arambourg (1952), Bergounioux (1956) et Romer (1956) comme un pholidosauridé et Roux et Douvillé (1910) comme un Plesiosauridae. Seuls von Huene (1956), Kuhn (1968) et Steel (1973) considérèrent les Dyrosauridae comme une famille distincte.
De nombreux noms de familles furent par ailleurs donnés pour les restes de Dyrosauridae, ainsi Louis Dollo (1914) créa les Congosauridae pour Congosaurus bequaerti, un dyrosaure du Paléocène de Cabinda (Angola), Swinton (1930) les Rhabdognathidae pour son Rhabdognathus rarus du Paléocène du Nigeria, et Bergounioux (1955) les Rhabdosauridae pour son Rhabdosaurus acutidentatus de Tunisie. Le nom Congosauridae fut d’ailleurs accepté par plusieurs auteurs comme Kuhn (1968) et Steel (1973), qui classaient dans cette famille tous les dyrosauridae connus à cette époque tel que C. bequaerti, Dyrosaurus phosphaticus, Phosphatosaurus gavialoides, Rhabdognathus, Rhabdosaurus, Wurnosaurus et Sokotosaurus (parmi ceux-ci, certains noms d’espèces n’étant plus considérés comme valides). La clarification du statut de cette famille ne fut apportée que par Buffetaut, qui considéra définitivement les Dyrosauridae comme une famille à part entière distincte (Buffetaut, 1976a, 1978b, 1981a), lui synonymisant tous les noms définis postérieurement. Il rapporta à cette famille, en plus de Dyrosaurus phosphaticus de Tunisie, Sokotosuchus ianwilsoni du Maastrichtien du Nigeria et précédemment considéré comme un Teleosauridae (Halstead, 1975) et Hyposaurus rogersii d’Amérique du Nord et considéré jusque-là comme un Goniopholididae (Mook, 1925 ; Steel, 1973) ou un Pholidosauridae (Kälin, 1955). Buffetaut(1976) créa plus tard le groupe des Tethysuchia, parfaitement synonyme du nom Dyrosauridae, et divisa ceux-ci en deux groupes : les Phosphatosaurinae (Dyrosauridae à dents robustes) et les Hyposaurinae (Dyrosauridae à dents grêles) (Buffetaut, 1981a).
Cladogramme
Le cladogramme suivant de Daniel Fortier et ses collègues en 2011et_al.''_2011_1-0">[1] montre la position des espèces au sein de la famille des Pholidosauridae :
Répartition géographique
En Amérique du Nord, où le premier dyrosaure a été décrit, plusieurs espèces ont été identifiées dans les sédiments Maastrichtiens et Paléocènes (Danien). Ils proviennent principalement du New Jersey (Owen, 1849 ; Leidy, 1865 ; Cope, 1868 ; Troxell, 1925c ; Miller, 1955a ; Wolfe, 1977 ; Parris, 1986 ; Denton et al., 1994, 1997), mais leur présence a aussi été signalée en Caroline du Sud, Alabama et peut-être en Virginie et Mississippi (Denton et al., 1994, 1997).
De nombreuses espèces ont été nommées, avec Hyposaurus rogersii (Owen, 1849), Hyposaurus ferox (March, 1871), Hyposaurus natator (Troxell, 1925c) et Hyposaurus natator oweni (Troxell, 1925c). Parmi celles-ci, seules H. rogersii et H. natator furent reconnues comme des espèces valides par Norell et Storrs (1989), alors que Denton et al. (1994, 1997) n’ont quant à eux considéré qu’une seule espèce, H. rogersii.
En Amérique du Sud, le spécimen le mieux préservé est représenté par une mandibule et quelques fragments crâniens et postcrâniens du Crétacé de l'État du Pernambouc au Brésil (Cope, 1885, 1886 ; Oliveira Roxo, 1937). Cope (1886) nomma ce spécimen Hyposaurus derbianus, et Woodward (1888) attribua à cette espèce du nouveau matériel du Crétacé Inférieur de Bahia, matériel attribué par la suite à Goniopholis (Mawson et Woodward, 1907), mais appartenant probablement à Sarcosuchus (Buffetaut et Taquet, 1977 ; Norell et Storrs, 1989).
Du Brésil, des fragments de vertèbres (Carvalho et Azevedo, 1997) ou des dents (Gallo et al., 2000) ont aussi été découverts dans la Formation Maria Farinha. Des autres pays sud-américains, seuls le Crétacé de Colombie (Langston, 1965) et le Paléocène de Bolivie (Argallo et al., 1987; Buffetaut, 1991a) ont fourni quelques restes fragmentaires, l’espèce Sulcusuchus erraini du Maastrichtien d’Argentine (Gasparini et Spalletti, 1990 ; Gasparini, 1996), n’étant pas, quant à elle, un dyrosauridé comme elle fut d’abord identifiée, mais un plésiosaure (Gasparini et Fuente, 2000).
En Asie, seules quelques vertèbres ont été signalées dans le Paléocène et l’Éocène du Pakistan (Buffetaut, 1976b, 1977a, 1978c ; Storrs, 1986), l’Éocène Supérieur de Birmanie (Buffetaut, 1977a, 1978d), et le Crétacé Supérieur d’Inde (Rajendra, 1987). D’autres restes provenant du Crétacé Inférieur d’Inde pourraient aussi être attribués aux Dyrosauridae (Lydekker, 1879 ; Pilgrim, 1940), sans toutefois, au vu de la faible quantité de matériel, que cette attribution soit des plus définitives. De tous ces fossiles, les restes les mieux conservés sont ceux d’une colonne vertébrale en connexion du Paléocène du Pakistan (Storrs, 1986), qui, comme tous les autres fossiles asiatiques, restent malheureusement trop fragmentaires pour être identifiés plus précisément.
En Europe, seule une symphyse mandibulaire du Cénomanien du Portugal pourrait se rapporter aux dyrosaures (Buffetaut et Lauverjat, 1978), qui, si cette attribution taxonomique s’avérait exacte, représenterait le reste le plus ancien de dyrosauridé connu. Un autre fragment a bien été signalé dans le Paléocène d’Espagne par Costa et al. (1995), mais sa fenêtre supratemporale réduite l’exclut d’emblée des Dyrosauridae.
Ainsi, même si les dyrosauridés sont répandus sur presque tous les continents, ils sont largement plus nombreux sur le continent africain. En Afrique du Nord, ils sont présents en Tunisie (Thomas, 1893 ; Pomel, 1894a, b ; Stefano, 1903 ; Thévenin, 1911 ; Joleau, 1922, a, b ; Jouve, 2005), Algérie (Piveteau, 1935) et Maroc (Brives, 1919 ; Gentil, 1922 ; Joleaud, 1922a, b, 1923, 1926 ; Depéret et Russo, 1924, 1925 ; Arambourg, 1934, 1952 ; Gigout, 1951 ; Ennouchi, 1957 ; Buffetaut, 1979a ; Hua, 1995 ; Jouve, 2004 ; Jouve et al., 2004, 2005a, 2005b). Arambourg (1952) et Bergounioux (1956) fournirent les études les plus détaillées de ces dyrosaures nord-africains.
Arambourg (1952) décrivit principalement du matériel marocain, mais aussi tunisien, et nomma Hyposaurus paucidens à partir d’une mandibule et quelques restes postcrâniens. Bergounioux (1955, 1956) quant à lui, nomma six nouvelles espèces de Dyrosaurus dans l’Éocène Inférieur de Tunisie, alors qu’une seule n’avait été reconnue jusque-là. Il nomma aussi deux nouveaux genres, Phosphatosaurus gavialoides et Rhabdosaurus acutidentatus. Buffetaut (1978b) et Moody et Buffetaut (1981) ne considérèrent qu’une seule espèce de Dyrosaurus comme valide lors de leur révision, apportèrent Rhabdosaurus à Rhabdognathus, et fournirent une description plus détaillée de Phosphatosaurus gavialoides.
La présence de P. gavialoides a par ailleurs été signalée au Niger et au Mali (Buffetaut, 1979b, 1980b), pays partiellement recouverts par le Bassin des Iullemmeden du Crétacé Supérieur à l’Éocène, et où de nombreux restes de dyrosauridés ont été décrits. Le premier fossile fut récolté par Cortier (1908) et décrit par Lemoine (1909) et Thévenin (1911b). À cause de leur abondance dans ce bassin, les dyrosaures furent signalés à de nombreuses reprises (Nopsca, 1925 ; Bourcart et Keller, 1929 ; Pérébaskine, 1933a, b ; Monod, 1939 ; Arambourg et Joleaud, 1943 ; Cornet, 1943 ; Jones, 1948 ; Karpoff et Visse, 1950 ; Radier, 1953, 1959 ; Lavocat et Radier, 1953 ; Lavocat, 1953a, b, 1955b ; Karpoff et al., 1954 ; Swinton 1930 ; Kogbe, 1975, 1979 ; Halstead et Middleton, 1976), mais Swinton (1930) fut véritablement le premier à en fournir la première étude exhaustive, en nommant trois nouvelles espèces : Wurnosaurus wilsoni, Sokotosaurus nopcsai et Rhabdognathus rarus. La révision des espèces du Bassin des Iullemmeden permit à Buffetaut (1980b) de transférer W. wilsoni et S. nopcsai dans Hyposaurus, et d’ériger deux nouvelles espèces, R. compressus et Tilemsisuchus lavocati. Un crâne du Mali, particulièrement bien préservé, fut décrit bien plus tard et attribué à Rhabdognathus sp. par Brochu et al. (2002). Ce crâne et celui de Dyrosaurus phosphaticus décrit par Jouve (2004) restent les mieux préservés et décrits de tous les spécimens de dyrosauridés connus jusqu’à présent.
Le Maroc, a fourni récemment diverses nouvelles espèces telles Arambourgisuchus khouribgaensis (Jouve et al., 2005), et Chenanisuchus lateroculi (2005), particulièrement bien préservées.
Louis Dollo (1914) a brièvement décrit et nommé Congosaurus bequaerti, un dyrosaure du Paléocène d’Angola, spécimen décrit plus en détail par Swinton (1950), qu’Arambourg (1952) et Antunes (1964) considérèrent comme synonyme de Dyrosaurus. Buffetaut (1976a, 1980b) quant à lui, le considéra comme appartenant au genre Hyposaurus, cette dernière attribution étant actuellement celle acceptée par la majorité des auteurs.
De nombreux restes ont été découverts dans de nombreux autres pays africains comme le Maastrichtien d’Égypte (Gemmellaro, 1921), le Paléocène du Togo (Stromer, 1910 ; Furon et Kouriatchy, 1948), le Paléocène et Éocène du Sénégal (Tessier, 1952), l’Éocène de Libye (Arambourg et Magnier, 1961), le Maastrichtien du Soudan (Buffetaut et al., 1990, Werner, 1993, 1994, 1996), le Paléogène d’Éthiopie (Wood et al., 1993 ; Werner, 1995 ; Goodwin at al., 1994) et le Paléocène d’Arabie Saoudite (Langston, 1995). Dans ce dernier pays de nombreux restes sont attribués à Hyposaurus sp. et Rhabdognathus sp.
Galeries des Dyrosauridae


Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- Jouve et al. (2008)
- ^ Jump up to:a b Hastings, A.K., Bloch, J. and Jaramillo, C.A. (2011). "A new longirostrine dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) from the Paleocene of north-eastern Colombia: biogeographic and behavioural implications for new-world dyrosauridae" (PDF). Paleontology 54 (5): 1095–116. doi:10.1111/j.1475-4983.2011.01092.x. Retrieved 14 Sep2011.
- Jump up^ Greigert, J. (1966). "Description des formations Crétacées et Tertiaires du bassin des Iullemmeden (Afrique occidentale)". Direction des Mines et de la Géologie, Niger. Publication 2: 1–273.
- Jump up^ Reyment, R. (1980). "Biogeography of the Saharan Cretaceous and Paleocene epicontinental transgressions". Cretaceous Research 1: 299–327. doi:10.1016/0195-6671(80)90041-5.
- Jump up^ Buffetaut, E.; Bussert, R.; Brinkmann, W. (1990). "A new nonmarine vertebrate fauna in the Upper Cretaceous of northern Sudan". Berliner Geowissenschaftlische Abhandlungen 120: 183–202.
- Jump up^ Hastings, A. K; Bloch, J. I.; Cadena, E. A.; Jaramillo, C. A. (2010). "A new small short-snouted dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) from the Paleocene of northeastern Colombia". Journal of Vertebrate Paleontology 30 (1): 139–162. doi:10.1080/02724630903409204.
- Jump up^ Head, J. J.; Bloch, J. I.; Hastings, A. K.; Borque, J. R.; Cadena, E. A.; Herrera, F. A.; Polly, P. D.; Jaramillo, C. A. (2009). "Giant boid snake from the Palaeocene neotropics reveals hotter past equatorial temperatures". Nature 457 (7230): 715–717. doi:10.1038/nature07671. .
Voir aussi
Liens externes
- Ressources relatives au vivant :
Notes et références
- et_al.''_2011-1" class="mw-reference-text">(en) Daniel Fortier, Perea, Daniel et Schultz, Cesar, « Redescription and phylogenetic relationships of Meridiosaurus vallisparadisi, a pholidosaurid from the Late Jurassic of Uruguay », Zoological Journal of the Linnean Society, vol. 163, no Supplement S1, , S257 (DOI 10.1111/j.1096-3642.2011.00722.x).