Plesiosauria
Plésiosauriens

Taxons de rang inférieur
- super-famille †Plesiosauroidea
- sous-ordre †Pliosauroidea
Les Plésiosauriens forment l'ordre éteint des Plesiosauria (signifiant « presque reptiles », du grec πλησίος, plesios : « proche », et σάυρια, sauria : « lézard »), regroupant des reptiles marins qui vivait au Mésozoïque. Ce groupe fait partie des Sauroptérygiens, et inclut les plésiosaures et les pliosaures.
Ils apparurent pour la première fois dans la dernière période du Trias, peut-être à l'étage Rhaetien, il y a environ 203 millions d'années[1]. Ils proliférèrent durant le Jurassique jusqu'à l'extinction massive qui survint à la fin du Crétacé, il y a environ 66 Ma. Ils avaient une distribution océanique mondiale.
Les plésiosaures ont été parmi les premiers reptiles fossiles découverts. Au début du XIXe siècle, les scientifiques ont réalisé à quel point leur construction était distinctive et ils ont été nommés en tant qu'ordre distinct en 1835. Le premier genre de plésiosaure, l'éponyme Plesiosaurus, a été nommé en 1821. Depuis lors, plus d'une centaine d'espèces valides ont été décrit. Au début du XXIe siècle, le nombre de découvertes a augmenté, conduisant à une meilleure compréhension de leur anatomie, de leurs relations et de leur mode de vie.
Les plésiosaures avaient un corps large et plat et une queue courte. Leurs membres s'étaient transformés en quatre longues nageoires, alimentées par de puissants muscles attachés à de larges plaques osseuses formées par la ceinture scapulaire et le bassin. Les palmes faisaient un mouvement de vol dans l'eau. Les plésiosaures respiraient de l'air et portaient des petits vivants ; il y a des indications qu'ils étaient à sang chaud.
Les plésiosaures présentaient deux types morphologiques principaux. Certaines espèces, avec la constitution « plésiosauromorphe », avaient un cou (parfois extrêmement) long et une petite tête ; ceux-ci étaient relativement lents et attrapaient de petits animaux marins. D'autres espèces, certaines atteignant une longueur de dix-sept mètres, avaient la forme « pliosauromorphe » avec un cou court et une grosse tête ; c'étaient des superprédateurs, des chasseurs rapides de grosses proies. Les deux types sont liés à la division stricte traditionnelle des Plesiosauria en deux sous-ordres, les Plesiosauroidea à long cou et les Pliosauroidea à cou court. La recherche moderne, cependant, indique que plusieurs groupes « à long cou » pourraient avoir eu des membres à cou court ou vice versa. Par conséquent, les termes purement descriptifs « plésiosauromorphe » et « pliosauromorphe » ont été introduits, qui n'impliquent pas une relation directe. « Plesiosauroidea » et « Pliosauroidea » ont aujourd'hui un sens plus limité. Le terme « plésiosaure » est correctement utilisé pour désigner les Plesiosauria dans son ensemble, mais de manière informelle, il est parfois destiné à indiquer uniquement les formes à long cou, l'ancien Plesiosauroidea.
Histoire de leur découverte
Premières trouvailles
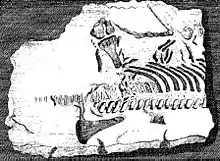
Les éléments squelettiques des plésiosaures sont parmi les premiers fossiles de reptiles éteints reconnus comme tels[2]. En 1605, Richard Verstegen d'Anvers a illustré dans son A Restitution of Decayed Intelligence des vertèbres de plésiosaure qu'il qualifiait de poissons et qu'il considérait comme la preuve que la Grande-Bretagne était autrefois connectée au continent européen[3]. Le Gallois Edward Lhuyd dans son Lithophylacii Brittannici Ichnographia de 1699 comprenait également des représentations de vertèbres de plésiosaure qui étaient à nouveau considérées comme des vertèbres de poisson ou d'Ichthyospondyli[4]. D'autres naturalistes au cours du XVIIe siècle ont ajouté des restes de plésiosaures à leurs collections, comme John Woodward ; ceux-ci n'ont été compris que beaucoup plus tard comme étant de nature plésiosaurienne et sont aujourd'hui en partie conservés au Sedgwick Museum[2].
En 1719, William Stukeley a décrit un squelette partiel d'un plésiosaure, qui avait été porté à son attention par l'arrière-grand-père de Charles Darwin, Robert Darwin d'Elston. La plaque de pierre provenait d'une carrière à Fulbeck dans le Lincolnshire et avait été utilisée, avec le fossile à sa face inférieure, pour renforcer la pente d'un point d'eau à Elston dans le Nottinghamshire. Après la découverte des ossements étranges qu'il contenait, il a été exposé dans le presbytère local comme la dépouille d'un pécheur noyé lors du déluge. Stukely a affirmé sa nature « diluviale », mais a compris qu'il représentait une créature marine, peut-être un crocodile ou un dauphin[5]. Le spécimen est aujourd'hui conservé au Muséum d'histoire naturelle de Londres, son numéro d'inventaire étant BMNH R.1330. C'est le premier squelette fossile plus ou moins complet de reptile découvert dans une collection de musée. Il peut peut-être être référé à Plesiosaurus dolichodeirus[2].
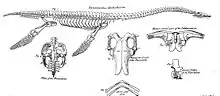
Au cours du XVIIIe siècle, le nombre de découvertes de plésiosaures anglais augmenta rapidement, bien qu'elles fussent toutes de nature plus ou moins fragmentaire. Des collectionneurs importants étaient les révérends William Mounsey et Baptist Noel Turner, actifs dans la vallée de Belvoir, dont les collections ont été décrites en 1795 par John Nicholls dans la première partie de son livre The History and Antiquities of the County of Leicestershire[6]. L'un des squelettes partiels de plésiosaure de Turner est toujours conservé en tant que spécimen BMNH R.45 au British Museum of Natural History ; c'est ce qu'aujourd'hui on appelé Thalassiodracon[2].
Dénomination du Plesiosaurus

Au début du XIXe siècle, les plésiosaures étaient encore mal connus et leur construction particulière n'était pas comprise. Aucune distinction systématique n'a été faite avec les ichtyosaures, de sorte que les fossiles d'un groupe ont parfois été combinés avec ceux de l'autre pour obtenir un spécimen plus complet. En 1821, un squelette partiel découvert dans la collection du colonel Thomas James Birch[7], a été décrit par William Conybeare et Henry Thomas De la Beche, et reconnu comme représentant un groupe distinctif. Un nouveau genre a été nommé, Plesiosaurus. Le nom générique est dérivé du grec πλήσιος, plèsios, « plus proche de » et du latinisé saurus, au sens de « saurien », pour exprimer que Plesiosaurus était dans la chaîne des êtres plus proche des Sauria, en particulier du crocodile, que d'Ichthyosaurus, qui avait la forme d'un poisson plus humble[8]. Le nom doit donc être lu plutôt comme « s'approchant du Sauria » ou « près du reptile » que comme « près du lézard »[9]. Des parties du spécimen sont encore présentes au Musée d'histoire naturelle de l'université d'Oxford[2].
Peu de temps après, la morphologie est devenue beaucoup plus connue. En 1823, Thomas Clark a signalé un crâne presque complet, appartenant probablement à Thalassiodracon, qui est maintenant conservé par le British Geological Survey en tant que spécimen BGS GSM 26035[2]. La même année, la collectionneuse commerciale de fossiles Mary Anning (1799 - 1847) et sa famille ont découvert un squelette presque complet de plésiosaure (Plesiosaurus dolichodeirus) à Lyme Regis dans le Dorset au Royaume-Uni, qui est devenu le 'fossile type'. Cette région de Grande-Bretagne est maintenant un site du Patrimoine mondial de l'UNESCO (voir le Littoral du Dorset et de l'est du Devon). Le squelette a été acquis par le duc de Buckingham, qui l'a mis à la disposition du géologue William Buckland. Il le laissa à son tour décrire par Conybeare le lors d'une conférence à la Société géologique de Londres[10], au cours de la même réunion au cours de laquelle pour la première fois un dinosaure fut nommé, Megalosaurus. Les deux découvertes ont révélé la construction unique et bizarre des animaux, comparée en 1832 par le professeur Buckland à « un serpent de mer traversant une tortue ». En 1824, Conybeare a également donné un nom spécifique à Plesiosaurus : dolichodeirus, qui signifie « longcou ». En 1848, le squelette a été acheté par le British Museum of Natural History et catalogué comme le spécimen BMNH 22656[2]. Lorsque la conférence a été publiée, Conybeare a également nommé une deuxième espèce : Plesiosaurus giganteus. Il s'agissait d'une forme à col court attribuée plus tard au Pliosauroidea[11].

Les plésiosaures sont devenus mieux connus du grand public grâce à deux publications richement illustrées du collectionneur Thomas Hawkins : Mémoires d'Ichthyosauri et Plesiosauri de 1834[12] et The Book of the Great Sea-Dragons de 1840. Hawkins avait une vision très idiosyncratique des animaux[13], les voyant comme des créations monstrueuses du diable, au cours d'une phase préadamitique de l'histoire[14]. Hawkins a finalement vendu ses spécimens précieux et joliment restaurés au British Museum of Natural History[15].
Au cours de la première moitié du XIXe siècle, le nombre de découvertes de plésiosaures a régulièrement augmenté, notamment grâce aux découvertes dans les falaises marines de Lyme Regis. Sir Richard Owen a nommé à lui seul près d'une centaine de nouvelles espèces. La majorité de leurs descriptions reposaient cependant sur des ossements isolés, sans diagnostic suffisant pour pouvoir les distinguer des autres espèces précédemment décrites. Bon nombre des nouvelles espèces décrites à cette époque ont par la suite été invalidées. Le genre Plesiosaurus est particulièrement problématique, car la majorité des nouvelles espèces y ont été placées de sorte qu'il est devenu un taxon poubelle. Peu à peu, d'autres genres ont été nommés. Hawkins avait déjà créé de nouveaux genres, bien que ceux-ci ne soient plus considérés comme valides. En 1841, Owen nomma Pliosaurus brachydeirus. Son étymologie faisait référence au Plesiosaurus dolichodeirus antérieur car il est dérivé de πλεῖος, pleios, « plus complètement », reflétant que selon Owen, il était plus proche du Sauria que du Plesiosaurus. Son nom spécifique signifie « avec un cou court »[16]. Plus tard, les Pliosauridae ont été reconnus comme ayant une morphologie fondamentalement différente des plésiosauridés. La famille Plesiosauridae avait déjà été inventée par John Edward Gray en 1825[17]. En 1835, Henri Marie Ducrotay de Blainville nomma l'ordre Plesiosauria lui-même[18].
Découvertes américaines
Dans la seconde moitié du XIXe siècle, d'importantes découvertes ont été faites en dehors de l'Angleterre. Bien que cela comprenait certaines découvertes allemandes, il s'agissait principalement de plésiosaures trouvés dans les sédiments de la voie maritime intérieure du Crétacé américain, la craie de Niobrara. Un fossile en particulier a marqué le début de la guerre des os entre les paléontologues rivaux Edward Drinker Cope et Othniel Charles Marsh.
En 1867, le médecin Theophilus Turner près de Fort Wallace au Kansas a découvert un squelette de plésiosaure, qu'il a fait don à Cope[19]. Cope a tenté de reconstruire l'animal en supposant que l'extrémité la plus longue de la colonne vertébrale était la queue, la plus courte le cou. Il s'aperçut bientôt que le squelette qui se formait sous ses mains avait des qualités très particulières : les vertèbres du cou avaient des chevrons et avec les vertèbres de la queue les surfaces articulaires étaient orientées d'arrière en avant[20]. Excité, Cope conclut avoir découvert un tout nouveau groupe de reptiles : les Streptosauria ou « Sauriens tournés », qui se distingueraient par des vertèbres inversées et un manque de membres postérieurs, la queue assurant la propulsion principale[21]. Après avoir publié une description de cet animal[22], suivie d'une illustration dans un manuel sur les reptiles et les amphibiens[23], Cope a invité Marsh et Joseph Leidy à admirer son nouvel Elasmosaurus platyurus. Après avoir écouté l'interprétation de Cope pendant un certain temps, Marsh a suggéré qu'une explication plus simple de la construction étrange serait que Cope avait inversé la colonne vertébrale par rapport au corps dans son ensemble. Lorsque Cope a réagi avec indignation à cette suggestion, Leidy a silencieusement pris le crâne et l'a placé contre la dernière vertèbre de la queue présumée, à laquelle il s'adaptait parfaitement : c'était en fait la première vertèbre du cou, avec encore un morceau de l'arrière du crâne attaché à elle[24]. Mortifié, Cope a essayé de détruire l'intégralité de l'édition du manuel et, lorsque cela a échoué, a immédiatement publié une édition améliorée avec une illustration correcte mais une date de publication identique[25]. Il a excusé son erreur en prétendant qu'il avait été induit en erreur par Leidy lui-même, qui, décrivant un spécimen de Cimoliasaurus, avait également inversé la colonne vertébrale[26]. Marsh a affirmé plus tard que l'affaire était la cause de sa rivalité avec Cope : « il a depuis été mon ennemi acharné ». Cope et Marsh, dans leur rivalité, ont nommé de nombreux genres et espèces de plésiosaures, dont la plupart sont aujourd'hui considérés comme invalides[27].
Au tournant du siècle, la plupart des recherches sur les plésiosaures ont été effectuées par un ancien élève de Marsh, le professeur Samuel Wendell Williston. En 1914, Williston a publié son Water reptiles of the past and present[28]. Malgré le traitement des reptiles marins en général, il restera pendant de nombreuses années le texte général le plus complet sur les plésiosaures[29]. En 2013, un premier manuel moderne était en préparation par Olivier Rieppel. Au milieu du XXe siècle, les États-Unis restent un centre de recherche important, notamment grâce aux découvertes de Samuel Paul Welles.
Découvertes récentes
Alors qu'au XIXe et pendant la majeure partie du XXe siècle, de nouveaux plésiosaures étaient décrits à raison de trois ou quatre genres chaque décennie, le rythme s'est soudainement accéléré dans les années 1990, avec dix-sept plésiosaures découverts au cours de cette période. Le rythme des découvertes s'est accéléré au début du XXIe siècle, avec environ trois ou quatre plésiosaures nommés chaque année[30]. Cela implique qu'environ la moitié des plésiosaures connus sont relativement nouveaux pour la science, résultat d'une recherche sur le terrain beaucoup plus intense. Certaines de ces activités se déroulent en dehors des zones traditionnelles, par ex. dans de nouveaux sites développés en Nouvelle-Zélande, en Argentine, au Chili[31], en Norvège, au Japon, en Chine et au Maroc, mais les emplacements des découvertes les plus originales se sont avérés encore productifs, avec de nouvelles découvertes importantes en Angleterre et en Allemagne. Certains des nouveaux genres sont un renommage d'espèces déjà connues, qui ont été jugées suffisamment différentes pour justifier un nom de genre distinct.
En 2002, le "Monstre d'Aramberri" est apparu dans la presse. Découvert en 1982 dans le village d'Aramberri, dans l'État mexicain de Nuevo León, il fut originellement considéré comme un dinosaure. Le spécimen est en réalité un très grand Plesiosauroidea, atteignant 15 m de long. Les médias ont publié une taille exagérée avec une longueur de 25 m et un poids de 150 000 kg, ce qui en aurait fait le plus grand prédateur de tous les temps. Cette erreur fut reprise dans le documentaire de la BBC Sur la terre des dinosaures, qui l'a aussi prématurément considéré comme un Liopleurodon ferox[32] - [33].
En 2004, ce qui semblait être un plésiosaure juvénile complètement intact a été découvert, par un pêcheur local, dans la réserve naturelle nationale de la baie de Bridgwater dans le Somerset, au Royaume-Uni. Le fossile, datant d'il y a 180 millions d'années comme l'indiquent les ammonites qui lui sont associées, mesurait 1,5 mètre de long et pourrait être apparenté au Rhomaleosaurus. C'est probablement le spécimen le mieux conservé d'un plésiosaure encore découvert[34] - [35] - [36].
En 2005, les restes de trois plésiosaures (Dolichorhynchops herschelensis) découverts dans les années 1990 près de Herschel, en Saskatchewan, se sont révélés être une nouvelle espèce, par le Dr Tamaki Sato, un paléontologue japonais des vertébrés[37].
En 2006, une équipe combinée d'enquêteurs américains et argentins (ce dernier de l'Institut antarctique argentin et du musée La Plata) a trouvé le squelette d'un plésiosaure juvénile mesurant 1,5 mètre de long sur l'île Vega en Antarctique[38]. Le fossile est actuellement exposé au musée géologique de la South Dakota School of Mines and Technology[39].
En 2008, les restes fossiles d'un plésiosaure non décrit nommé Predator X, maintenant connu sous le nom de Pliosaurus funkei, ont été déterrés à Svalbard. Il avait une longueur de 12 m et sa force de morsure de 149 kilonewtons est l'une des plus puissants connus[40].
En , un grand squelette de plésiosaure a été trouvé sur le continent de l'Antarctique, la plus ancienne créature du continent et la première de ses espèces en Antarctique[41].
Non seulement le nombre de découvertes sur le terrain a augmenté, mais aussi, depuis les années 1950, les plésiosaures font l'objet de travaux théoriques plus poussés. La méthodologie de la cladistique a, pour la première fois, permis le calcul exact de leurs relations évolutives. Plusieurs hypothèses ont été publiées sur la façon dont ils chassaient et nageaient, incorporant des idées modernes générales sur la biomécanique et l'écologie. Les nombreuses découvertes récentes ont testé ces hypothèses et en ont fait naître de nouvelles.
Évolution

Les Plesiosauria ont leurs origines au sein des Sauropterygia, un groupe de reptiles peut-être archosauromorphes qui sont retournés à la mer. Un sous-groupe avancé de sauroptérygiens, les Eusauropterygia carnivores avec de petites têtes et de longs cous, s'est divisé en deux branches au cours du Trias supérieur. L'un d'eux, les Nothosauroidea, a conservé des articulations fonctionnelles du coude et du genou ; mais l'autre, les Pistosauria, s'est mieux adaptée à un mode de vie marin. Leur colonne vertébrale est devenue plus rigide et la propulsion principale pendant la nage ne venait plus de la queue mais des membres, qui se sont transformés en nageoires[42]. Les Pistosauria sont devenus à sang chaud et vivipares, donnant naissance à des jeunes vivants[43]. Les premiers membres basaux du groupe, traditionnellement appelés « pistosauridés », étaient encore en grande partie des animaux côtiers. Leurs ceintures scapulaires restaient faibles, leur bassin ne pouvait pas supporter la puissance d'un coup de nage puissant et leurs nageoires étaient émoussées. Plus tard, un groupe de pistosauriens plus avancé s'est séparé : les Plesiosauria. Ceux-ci avaient des ceintures scapulaires renforcées, des bassins plus plats et des nageoires plus pointues. D'autres adaptations leur permettant de coloniser les mers ouvertes comprenaient des articulations raides des membres; une augmentation du nombre de phalanges de la main et du pied; une connexion latérale plus serrée de la série des phalanges des doigts et des orteils et une queue raccourcie[44] - [45].

Depuis le Jurassique le plus ancien, l'étage Hettangien, un riche rayonnement de plésiosaures est connu, ce qui implique que le groupe doit déjà s'être diversifié au Trias supérieur ; de cette diversification, cependant, seules quelques formes très basiques ont été découvertes. L'évolution ultérieure des plésiosaures est très controversée. Les différentes analyses cladistiques n'ont pas abouti à un consensus sur les relations entre les principaux sous-groupes de plésiosaures. Traditionnellement, les plésiosaures ont été divisés en Plesiosauroidea à long cou et Pliosauroidea à cou court. Cependant, la recherche moderne suggère que certains groupes généralement à long cou pourraient avoir eu des membres à cou court. Pour éviter toute confusion entre la phylogénie, les relations évolutives, et la morphologie, la façon dont l'animal est construit, les formes à long cou sont donc appelées « plésiosauromorphes » et les formes à cou court sont appelées « pliosauromorphes », sans que l'espèce « plésiosauromorphe » soit nécessairement plus étroitement liés les uns aux autres qu'aux formes « pliosauromorphes »[46].
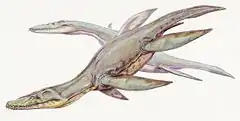
Le dernier ancêtre commun des Plesiosauria était probablement une forme plutôt petite à cou court. Au début du Jurassique, le sous-groupe avec le plus d'espèces était les Rhomaleosauridae, une scission peut-être très basale d'espèces qui étaient également à cou court. Les plésiosaures de cette période mesuraient au plus cinq mètres de long. Au Toarcien, il y a environ 180 millions d'années, d'autres groupes, parmi lesquels les Plesiosauridae, sont devenus plus nombreux et certaines espèces ont développé des cous plus longs, entraînant des longueurs totales de corps allant jusqu'à dix mètres[47].
Au milieu du Jurassique, de très grands Pliosauridae ont évolué. Ceux-ci étaient caractérisés par une grosse tête et un cou court, comme le Liopleurodon et le Simolestes. Ces formes avaient des crânes jusqu'à trois mètres de long et atteignaient une longueur allant jusqu'à dix-sept mètres et un poids de dix tonnes. Les pliosauridés avaient de grandes dents coniques et étaient les carnivores marins dominants de leur temps. Pendant la même période, il y a environ 160 millions d'années, étaient présents les Cryptoclididae, des espèces plus courtes avec un long cou et une petite tête[48].
Les Leptocleididae ont rayonné au Crétacé inférieur. Il s'agissait de formes plutôt petites qui, malgré leur cou court, auraient pu être plus étroitement apparentées aux Plesiosauridae qu'aux Pliosauridae. Plus tard au Crétacé inférieur, les Elasmosauridae sont apparus ; ceux-ci étaient parmi les plus longs plésiosaures, atteignant jusqu'à quinze mètres de longueur en raison de très longs cous contenant jusqu'à 76 vertèbres, plus que tout autre vertébré connu. Les Pliosauridae étaient toujours présents comme le montrent les grands prédateurs, tels que Kronosaurus[48].
Au début du Crétacé supérieur, les Ichthyosaures se sont éteints ; peut-être qu'un groupe de plésiosaures a évolué pour remplir leurs niches : les Polycotylidae, qui avaient un cou court et une tête particulièrement allongée avec un museau étroit. À la fin du Crétacé, les élasmosauridés comptaient encore de nombreuses espèces[48].
Tous les plésiosaures se sont éteints à la suite de l'événement K-T à la fin du Crétacé, il y a environ 66 millions d'années[49].
Relations
Dans la phylogénie moderne, les clades sont des groupes définis qui contiennent toutes les espèces appartenant à une certaine branche de l'arbre évolutif. Une façon de définir un clade est de le laisser se composer du dernier ancêtre commun de deux de ces espèces et de tous ses descendants. Un tel clade est appelé « clade de nœuds ». En 2008, Patrick Druckenmiller et Anthony Russell ont ainsi défini Plesiosauria comme le groupe constitué du dernier ancêtre commun de Plesiosaurus dolichocheirus et Peloneustes philarchus et de tous ses descendants[50]. Plesiosaurus et Peloneustes représentaient les principaux sous-groupes des Plesiosauroidea et des Pliosauroidea et ont été choisis pour des raisons historiques ; toute autre espèce de ces groupes aurait suffi.
Une autre façon de définir un clade est de le faire comprendre de toutes les espèces plus étroitement apparentées à une certaine espèce que l'on souhaite de toute façon inclure dans le clade qu'à une autre espèce que l'on souhaite au contraire exclure. Un tel clade est appelé « clade souche ». Une telle définition présente l'avantage qu'il est plus facile d'inclure toutes les espèces ayant une certaine morphologie. Plesiosauria était en 2010 par Hillary Ketchum et Roger Benson défini comme un tel taxon basé sur le tronc : « tous les taxons plus étroitement liés à Plesiosaurus dolichodeirus et Pliosaurus brachydeirus qu'à Augustasaurus hagdorni ». Ketchum et Benson (2010) ont également inventé un nouveau clade Neoplesiosauria, un taxon basé sur des nœuds défini par « Plesiosaurus dolichodeirus, Pliosaurus brachydeirus, leur ancêtre commun le plus récent et tous ses descendants »[48]. Le clade Neoplesiosauria est très probablement matériellement identique à Plesiosauria sensu Druckenmiller & Russell, donc désignerait exactement la même espèce, et le terme était censé remplacer ce concept.
Benson et al. (2012) ont trouvé que le Pliosauroidea traditionnel était paraphylétique par rapport au Plesiosauroidea. Rhomaleosauridae s'est avéré être en dehors de Neoplesiosauria, mais toujours dans Plesiosauria. Le pistosaure Bobosaurus du Carnien s'est avéré être un pas plus avancé qu'Augustasaurus par rapport au Plesiosauria et il représentait donc par définition le plésiosaure le plus connu. Cette analyse s'est concentrée sur les plésiosaures basaux et, par conséquent, seuls un pliosauridé dérivé et un cryptoclidien ont été inclus, tandis que les élasmosauridés n'ont pas été inclus du tout. Une analyse plus détaillée publiée à la fois par Benson et Druckenmiller en 2014 n'a pas été en mesure de résoudre les relations entre les lignées à la base de Plesiosauria[51].

Taxinomie
La classification des plésiosauriens a varié dans le temps ; la suivante présente une des versions actuelles (principalement celle d'O'Keefe de 2001)



- Super-ordre Sauropterygia
- Ordre Plesiosauria
- Super-famille Pliosauroidea
- Thalassiodracon
- Attenborosaurus
- Eurycleidus
- Famille Rhomaleosauridae
- Super-famille Plesiosauroidea
- Euplesiosauria
- Famille Elasmosauridae
- Super-famille Cryptoclidoidea
- Cryptoclididae
- Tricleidia
- Tricleidus
- Famille Cimoliasauridae
- Aristonectes
- Aristonectes parvidens
- Cimoliasaurus
- Scanisaurus
- Kimmerosaurus
- Aristonectes
- Famille Polycotylidae (= "Dolichorhynchopidae")
- Super-famille Pliosauroidea
- Ordre Plesiosauria
Le cladogramme des Plesiosauria suivant fait suite à une analyse de Benson & Druckenmiller (2014)[52].
| ||||||||||
| ◄ Plesiosauroidea |
| ||||||||||||||||||||||||||||||||||||
| ◄ Thalassophonea |
| |||||||||||||||||||||||||||||||||
Description
Taille

En général, les plésiosaures variaient en longueur d'adulte entre 1,5 mètre et environ 15 mètres. Le groupe contenait ainsi certains des plus grands superprédateurs marins des archives fossiles, équivalant à peu près à la taille des ichtyosaures, des mosasauridés, des requins et des baleines à dents les plus longs. Certains restes de plésiosaure, tels qu'un ensemble de mâchoires inférieures fragmentées et hautement reconstruites de 2,875 mètres de long conservés au musée de l'Université d'Oxford et se rapportant à Pliosaurus rossicus (précédemment appelé Stretosaurus[53] et Liopleurodon), indiquaient une longueur de 17 mètres. Cependant, il a été récemment soutenu que sa taille ne peut pas être actuellement déterminée en raison de leur mauvaise reconstruction et qu'une longueur de 12,7 mètres est plus probable[54]. MCZ 1285, un spécimen actuellement référé à Kronosaurus queenslandicus, du Crétacé inférieur d'Australie, aurait une longueur de crâne de 2,21 à 2,85 m[54] - [55].
Squelette

Les plésiosauriens possédaient un corps large et une queue relativement courte. Ils ont conservé leurs deux paires de membres ancestrales, qui avaient évolué en grandes nageoires[56]. Les plésiosauriens étaient eux-mêmes issus des nothosaures[57], qui avaient un corps plus semblable aux crocodiles. La disposition des nageoires est inhabituelle pour les animaux aquatiques en ce sens que les quatre membres ont probablement été utilisés pour propulser l'animal dans l'eau par des mouvements de haut en bas. La queue n'était probablement utilisée que pour aider au contrôle directionnel. Cela contraste avec les ichthyosaures et plus tard les mosasaures, dans lesquels la queue fournissait la propulsion principale[58].
Pour alimenter les nageoires, la ceinture scapulaire et le bassin avaient été considérablement modifiés, se développant en de larges plaques osseuses sous le corps, qui servaient de surface d'attache pour les grands groupes musculaires, capables de tirer les membres vers le bas. Au niveau de l'épaule, le coracoïde était devenue l'élément le plus volumineux recouvrant la majeure partie de la poitrine. L'omoplate était beaucoup plus petite, formant le bord avant extérieur du tronc. Au milieu, il a continué dans une clavicule et enfin un petit os interclaviculaire. Comme pour la plupart des tétrapodes, l'articulation de l'épaule était formée par l'omoplate et le coracoïde. Dans le bassin, la plaque osseuse était formée par l'ischion à l'arrière et le plus gros os pubien devant. L'ilion, qui chez les vertébrés terrestres porte le poids du membre postérieur, était devenu un petit élément à l'arrière, n'étant plus attaché ni à l'os pubien ni au fémur. L'articulation de la hanche était formée par l'ischion et l'os pubien. Les plaques pectorales et pelviennes étaient reliées par un plastron, une cage osseuse formée par les côtes ventrales appariées qui avaient chacune une section médiane et une section externe. Cet arrangement a immobilisé tout le tronc[58].
Pour devenir des palmes, les membres avaient considérablement changé. Les membres étaient très gros, chacun à peu près aussi long que le tronc. Les membres antérieurs et postérieurs se ressemblaient fortement. L'humérus du haut du bras et le fémur du haut de la jambe étaient devenus de gros os plats, dilatés à leurs extrémités externes. Les articulations du coude et du genou n'étaient plus fonctionnelles : l'avant-bras et le bas de la jambe ne pouvaient pas fléchir par rapport aux éléments des membres supérieurs, mais en formaient un prolongement plat. Tous les os externes étaient devenus des éléments de support plats des nageoires, étroitement liés les uns aux autres et à peine capables de tourner, de fléchir, de s'étendre ou de s'étendre. Cela était vrai du cubitus, du radius, des métacarpiens et des doigts, ainsi que du tibia, du péroné, des métatarses et des orteils. De plus, afin d'allonger les nageoires, le nombre de phalanges avait augmenté, jusqu'à dix-huit d'affilée, un phénomène appelé hyperphalangie. Les palmes n'étaient pas parfaitement plates, mais avaient un profil supérieur légèrement convexe, comme un profil aérodynamique, pour pouvoir « voler » dans l'eau[58].

Alors que les plésiosaures variaient peu dans la construction du tronc et pouvaient être qualifiés de « conservateurs » à cet égard, il existait des différences majeures entre les sous-groupes en ce qui concerne la forme du cou et du crâne. Les plésiosaures peuvent être divisés en deux types morphologiques principaux qui diffèrent par la taille de la tête et du cou. Les « plésiosauromorphes », comme les Cryptoclididae, les Elasmosauridae et les Plesiosauridae, avaient un long cou et une petite tête. Les « pliosauromorphes », comme les Pliosauridae et les Rhomaleosauridae, avaient un cou plus court avec une tête large et allongée. Les variations de longueur du cou n'étaient pas causées par un allongement des vertèbres cervicales individuelles, mais par l'augmentation du nombre de ces vertèbres cervicales. Elasmosaurus a soixante-douze vertèbres cervicales ; le record connu est détenu par l'élasmosauridé Albertonectes, avec soixante-seize cervicales[59]. Le grand nombre d'articulations a suggéré aux premiers chercheurs que le cou devait être très flexible ; en effet, une courbure du cou en forme de cygne était supposée possible - en islandais, les plésiosaures sont même appelés Svaneðlur, « lézards-cygnes ». Cependant, la recherche moderne a confirmé une conjecture antérieure de Williston selon laquelle les longues épines en forme de plaque au-dessus des vertèbres, le processus spinosi, limitaient fortement le mouvement vertical du cou. Bien que la courbure horizontale était moins restreinte, en général, le cou devait être plutôt raide et était certainement incapable d'être plié en serpentins. C'est encore plus vrai pour les « pliosauromophes » à cou court, qui n'avaient que onze vertèbres cervicales. Avec les formes précoces, les vertèbres du cou amphicoeles ou amphiplates portaient des côtes du cou à deux têtes ; les formes ultérieures avaient des côtes à une seule tête. Dans le reste de la colonne vertébrale, le nombre de vertèbres dorsales variait entre dix-neuf et trente-deux environ ; des vertèbres sacrées, entre deux et six, et des vertèbres caudales, entre vingt et un et trente-deux environ. Ces vertèbres possédaient encore les processus originaux hérités des ancêtres terrestres des Sauroptérygies et n'avaient pas été réduites à de simples disques ressemblant à des poissons, comme c'était le cas avec les vertèbres des ichtyosaures. Les vertèbres de la queue possédaient des os en chevron. Les vertèbres dorsales des plésiosaures sont facilement reconnaissables par deux grands foramens subcentralia, des ouvertures vasculaires appariées sur la face inférieure[58].
Le crâne des plésiosaures présentait l'état « euryapside », sans les fenêtres temporales inférieures, les ouvertures sur les côtés arrière inférieurs. Les fenêtres temporales supérieures formaient de grandes ouvertures sur les côtés du toit arrière du crâne, l'attache des muscles fermant les mâchoires inférieures. Généralement, les os pariétaux étaient très gros, avec une crête médiane, tandis que les os squamosaux formaient généralement un arc, excluant les pariétaux de l'occiput. Les orbites étaient larges, pointant en général obliquement vers le haut ; les pliosauridés avaient des yeux dirigés plus latéralement. Les yeux étaient soutenus par des anneaux scléraux dont la forme montre qu'ils étaient relativement plats, une adaptation à la plongée. Les narines internes placées antérieurement, les choanes, ont des sillons palatins pour canaliser l'eau, dont l'écoulement serait maintenu par une pression hydrodynamique sur les narines externes placées postérieurement, devant les orbites, pendant la nage. Selon une hypothèse, lors de son passage dans les conduits nasaux, l'eau aurait été « senti » par les épithéliums olfactifs[60] - [61]. Cependant, plus en arrière, une seconde paire d'ouvertures est présente en bouche ; une hypothèse ultérieure soutient que ce sont les véritables choanes et que la paire avant représentait en réalité des glandes salines appariées[62]. La distance entre les orbites et les narines était si limitée car les os du nez étaient fortement réduits, voire absents chez de nombreuses espèces. Les prémaxillaires touchaient directement les os frontaux ; chez les élasmosauridés, ils remontaient même jusqu'aux os pariétaux. Souvent, les os lacrymaux faisaient également défaut[45].

La forme et le nombre des dents étaient très variables. Certaines formes avaient des centaines de dents en forme d'aiguille. La plupart des espèces avaient des dents coniques plus grandes avec une section transversale ronde ou ovale. Ces dents étaient au nombre de quatre à six dans le prémaxillaire et d'environ quatorze à vingt-cinq dans le maxillaire ; le nombre dans les mâchoires inférieures équivalait à peu près à celui du crâne. Les dents étaient placées dans des empreintes dentaires, avaient un émail plissé verticalement et n'avaient pas de véritable tranchant ou carène. Chez certaines espèces, les dents de devant étaient notablement plus longues pour saisir les proies.
Tissus mous
Les restes de tissus mous de plésiosaures sont rares, mais parfois, en particulier dans les gisements de schiste, ils ont été partiellement préservés, par ex. montrant les contours du corps. Une première découverte à cet égard était l'holotype de Plesiosaurus conybeari (actuellement Attenborosaurus). D'après de telles découvertes, on sait que la peau était lisse, sans écailles apparentes mais avec de petites rides, que le bord de fuite des nageoires s'étendait considérablement derrière les os des membres[63] ; et que la queue portait une nageoire verticale, comme l'a rapporté Wilhelm Dames dans sa description de Plesiosaurus guilelmiimperatoris (actuellement Seeleyosaurus)[64]. La possibilité d'une douve de la queue a été confirmée par des études récentes sur la forme de l'épine neurale caudale de Pantosaurus, Cryptoclidus et Rhomaleosaurus zetlandicus[65] - [66] - [67]. Une étude de 2020 affirme que la nageoire caudale était de configuration horizontale[68].
Paléobiologie et comportement

Nourriture

Les cous extrêmement longs des « plésiosauromorphes » ont suscité des spéculations quant à leur fonction dès le moment où leur construction particulière est devenue apparente.
Conybeare avait proposé trois explications possibles. Le cou aurait pu servir à intercepter des poissons se déplaçant rapidement lors d'une poursuite. Alternativement, les plésiosaures auraient pu se reposer au fond de la mer, tandis que la tête était envoyée à la recherche de proies, ce qui semblait être confirmé par le fait que les yeux étaient dirigés relativement vers le haut. Enfin, Conybeare a suggéré la possibilité que les plésiosaures nageaient à la surface, laissant leur cou plonger vers le bas pour chercher de la nourriture à des niveaux inférieurs. Toutes ces interprétations supposaient que le cou était très flexible. L'idée moderne que le cou était, en fait, plutôt rigide, avec un mouvement vertical limité, a nécessité de nouvelles explications. Une hypothèse est que la longueur du cou permettait de surprendre les bancs de poissons, la tête arrivant avant que la vue ou l'onde de pression du tronc ne puisse les alerter. Les « plésiosauromorphes » chassaient visuellement, comme le montrent leurs grands yeux, et utilisaient peut-être un sens olfactif directionnel. Les Plésiosaures ont été découverts avec des fossiles de Belemnoidea (proches des seiches actuelles), et d'ammonites dans leurs estomacs. Leurs mâchoires étaient probablement suffisamment puissantes pour percer la coquille de leur proie[69]. Les poissons osseux (Osteichthyes), qui se sont encore diversifiés au cours du Jurassique, étaient probablement aussi des proies. Une hypothèse très différente prétend que les « plésiosauromorphes » étaient des mangeurs de fond. Les cous raides auraient servi à labourer les fonds marins, mangeant le benthos. Cela aurait été prouvé par les longs sillons présents dans les anciens fonds marins[70] - [71]. Un tel style de vie a été suggéré en 2017 pour Mortturneria[72]. Les « plésiosauromorphes » n'étaient pas bien adaptés à la capture de grosses proies se déplaçant rapidement, car leur long cou, bien qu'apparemment profilé, provoquait d'énormes frictions cutanées. Sankar Chatterjee a suggéré en 1989 que certains Cryptocleididae étaient des mangeurs de suspension, filtrant le plancton. Aristonectes, par ex. avait des centaines de dents, lui permettant de tamiser les petits crustacés de l'eau[73].
Les « pliosauromorphes » à cou court étaient les carnivores de sommet, ou superprédateurs, dans leurs réseaux trophiques respectifs[74]. C'étaient des prédateurs de poursuite[75] ou des prédateurs embusqués de proies de différentes tailles et des mangeurs opportunistes ; leurs dents pouvaient être utilisées pour percer des proies au corps mou, en particulier des poissons[76]. Leurs têtes et leurs dents étaient très grandes, adaptées pour saisir et déchirer les gros animaux. Leur morphologie permettait une vitesse de nage élevée. Eux aussi chassaient visuellement.
Les plésiosaures étaient eux-mêmes la proie d'autres carnivores, comme le montrent les marques de morsure laissées par un requin qui ont été découvertes sur une nageoire de plésiosaure fossilisée[77] et les restes fossilisés du contenu de l'estomac d'un mosasaure que l'on pense être les restes d'un plésiosaure[78].

Des squelettes ont également été découverts avec des gastrolithes, des pierres, dans leur estomac, mais il n'a pas été établi que ce soit pour aider à décomposer les aliments, en particulier les céphalopodes, dans un gésier musculaire, ou pour varier la flottabilité, ou les deux[79] - [80]. Cependant, le poids total des gastrolithes trouvés dans divers spécimens semble insuffisant pour modifier la flottabilité de ces grands reptiles[81]. Les premiers gastrolithes de plésiosaure, trouvés avec Mauisaurus gardneri, ont été rapportés par Harry Govier Seeley en 1877[82]. Le nombre de ces pierres par individu est souvent très important. En 1949, un fossile d'Alzadasaurus (spécimen SDSM 451, rebaptisé plus tard Styxosaurus) en montra 253[83]. La taille individuelle des pierres est souvent considérable. En 1991, un spécimen d'élasmosauridé, KUVP 129744, a été étudié, contenant un gastrolithe d'un diamètre de dix-sept centimètres et d'un poids de 1 300 grammes ; et une pierre un peu plus courte de 1 490 grammes. Au total, quarante-sept gastrolithes étaient présents, avec un poids combiné de 13 kilogrammes. La taille des pierres a été considérée comme une indication qu'elles n'ont pas été avalées par accident, mais délibérément, l'animal ayant peut-être parcouru de grandes distances à la recherche d'un type de roche approprié[84].
Mouvement de battement
La forme distinctive du corps à quatre nageoires a suscité de nombreuses spéculations sur le type d'utilisation des plésiosaures. Le seul groupe moderne à quatre nageoires est celui des tortues marines, qui n'utilisent que la paire avant pour la propulsion. Conybeare et Buckland avaient déjà comparé les nageoires avec des ailes d'oiseau. Cependant, une telle comparaison n'était pas très informative, car la mécanique du vol des oiseaux à cette période était mal comprise. Au milieu du XIXe siècle, on supposait généralement que les plésiosaures utilisaient un mouvement d'aviron. Les palmes auraient été déplacées vers l'avant dans une position horizontale, pour minimiser le frottement, puis tournées axialement vers une position verticale afin d'être tirées vers l'arrière, provoquant la plus grande force réactive possible. En fait, une telle méthode serait très inefficace : la course de récupération dans ce cas ne génère aucune poussée et la course arrière génère une énorme turbulence. Au début du XXe siècle, les principes nouvellement découverts du vol des oiseaux ont suggéré à plusieurs chercheurs que les plésiosaures, comme les tortues et les pingouins, effectuaient un mouvement de vol en nageant. C'était par ex. proposé par Eberhard Fraas en 1905[85], et en 1908 par Othenio Abel[86]. En vol, le mouvement de la nageoire est plus vertical, sa pointe décrivant un ovale ou "8". Idéalement, la palme est d'abord déplacée obliquement vers l'avant et vers le bas puis, après une légère rétraction et rotation, traverse cette trajectoire par le bas pour être tirée vers l'avant et vers le haut. Pendant les deux courses, vers le bas et vers le haut, selon le principe de Bernoulli, la poussée vers l'avant et vers le haut est générée par le profil supérieur incurvé de manière convexe de la nageoire, le bord avant légèrement incliné par rapport au débit d'eau, tandis que la turbulence est minime. Cependant, malgré les avantages évidents d'une telle méthode de nage, en 1924 la première étude systématique sur la musculature des plésiosaures par David Meredith Seares Watson a conclu qu'ils effectuaient néanmoins un mouvement d'aviron[87].
Au milieu du vingtième siècle, le « modèle de l'aviron » de Watson est resté l'hypothèse dominante concernant la nage des plésiosaures. En 1957, Lambert Beverly Halstead, utilisant à l'époque le nom de famille Tarlo, proposa une variante : les membres postérieurs auraient ramé dans le plan horizontal mais les membres antérieurs auraient ramé, déplacés vers le bas et vers l'arrière[88] - [89]. En 1975, le modèle traditionnel est remis en cause par Jane Ann Robinson, qui relance l'hypothèse du « vol ». Elle a fait valoir que les principaux groupes musculaires étaient placés de manière optimale pour un mouvement vertical des nageoires, pas pour tirer les membres horizontalement, et que la forme des articulations des épaules et des hanches aurait empêché la rotation verticale nécessaire pour ramer[90]. Dans un article ultérieur, Robinson a proposé que l'énergie cinétique générée par les forces exercées sur le tronc par les coups, aurait été stockée et libérée sous forme d'énergie élastique dans la cage thoracique, permettant un système de propulsion particulièrement efficace et dynamique[91].
Dans le modèle de Robinson, à la fois la descente et la montée auraient été puissantes. En 1982, elle a été critiquée par Samuel Tarsitano, Eberhard Frey et Jürgen Riess, qui ont affirmé que, alors que les muscles sous les plaques de l'épaule et du bassin étaient clairement suffisamment puissants pour tirer les membres vers le bas, des groupes musculaires comparables sur le dessus de ces les plaques pour élever les membres manquaient tout simplement, et, si elles avaient été présentes, n'auraient pas pu être employées avec force, leur renflement risquant de blesser les organes internes. Ils ont proposé un modèle de vol plus limité dans lequel une course descendante puissante était combinée à une récupération largement non alimentée, la nageoire revenant à sa position d'origine par l'élan du corps se déplaçant vers l'avant et coulant temporairement[92] - [93]. Ce modèle volant modifié est devenu une interprétation populaire. Moins d'attention a été accordée à une hypothèse alternative de Stephen Godfrey en 1984, qui proposait que les membres antérieurs et postérieurs effectuaient un mouvement de pagaie profond vers l'arrière combiné à un mouvement de récupération motorisé vers l'avant, ressemblant au mouvement effectué par les membres antérieurs des lions de mer[94].
En 2010, Frank Sanders et Kenneth Carpenter ont publié une étude concluant que le modèle de Robinson était correct. Frey & Riess se seraient trompés en affirmant que les plaques de l'épaule et du bassin n'avaient pas de muscles attachés à leurs côtés supérieurs. Alors que ces groupes musculaires n'étaient probablement pas très puissants, cela aurait pu être facilement compensé par les gros muscles du dos, en particulier le grand dorsal, qui aurait été bien développé compte tenu des hautes épines de la colonne vertébrale. De plus, la construction plate des articulations de l'épaule et de la hanche indiquait fortement que le mouvement principal était vertical et non horizontal[95].
Démarche
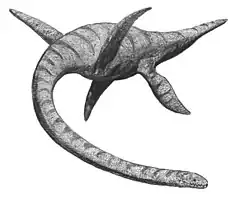
Comme tous les tétrapodes dotés de membres, les plésiosaures devaient avoir une certaine démarche, un modèle de mouvement coordonné des nageoires, dans ce cas, . Parmi le nombre infini de possibilités, dans la pratique, l'attention a été largement dirigée vers la question de savoir si la paire avant et la paire arrière se déplaçaient simultanément, de sorte que les quatre nageoires étaient engagées au même moment, ou selon un schéma alternatif, chaque paire étant employée à son tour. Frey & Riess en 1991 ont proposé un modèle alternatif, qui aurait eu l'avantage d'une propulsion plus continue[96]. En 2000, Theagarten Lingham-Soliar a éludé la question en concluant que, comme les tortues marines, les plésiosaures n'utilisaient la paire avant que pour un coup motorisé. La paire de derrière aurait été simplement utilisée pour la direction. Lingham-Soliar a déduit cela de la forme de l'articulation de la hanche, qui n'aurait permis qu'un mouvement vertical limité. De plus, une séparation de la fonction de propulsion et de direction aurait facilité la coordination générale du corps et évité un tangage trop extrême. Il a rejeté l'hypothèse de Robinson selon laquelle l'énergie élastique était stockée dans la cage thoracique, considérant que les côtes étaient trop rigides pour cela[97].
L'interprétation de Frey & Riess est devenue dominante, mais a été contestée en 2004 par Sanders, qui a montré expérimentalement que, alors qu'un mouvement alternatif aurait pu provoquer un tangage excessif, un mouvement simultané n'aurait provoqué qu'un léger tangage, ce qui aurait pu être facilement contrôlé par les nageoires postérieures. Parmi les autres mouvements axiaux, le roulis aurait pu être contrôlé en engageant alternativement les nageoires du côté droit ou gauche, et le lacet par le long cou ou une nageoire caudale verticale. Sanders ne croyait pas que la paire postérieure n'était pas utilisée pour la propulsion, concluant que les limitations imposées par l'articulation de la hanche étaient très relatives[98]. En 2010, Sanders & Carpenter concluaient qu'avec une allure alternée, les turbulences provoquées par le couple avant auraient entravé une action efficace du couple arrière. De plus, une longue phase de glisse après un engagement simultané aurait été très économe en énergie[95]. Il est également possible que l'allure soit facultative et qu'elle ait été adaptée aux circonstances. Lors d'une poursuite rapide et régulière, un mouvement alternatif aurait été utile ; dans une embuscade, un coup simultané aurait permis une vitesse de pointe. Lors de la recherche de proies sur une plus longue distance, une combinaison d'un mouvement simultané avec un vol plané aurait coûté le moins d'énergie[99]. En 2017, une étude de Luke Muscutt, utilisant un modèle de robot, a conclu que les nageoires arrière étaient activement utilisées, permettant une augmentation de 60 % de la force de propulsion et une augmentation de 40 % de l'efficacité. Le coup aurait été le plus puissant en utilisant une allure légèrement alternée, les nageoires arrière s'engageant juste après les nageoires avant, pour profiter de leur sillage. Cependant, il n'y aurait pas eu une seule phase optimale pour toutes les conditions, la démarche ayant probablement été modifiée en fonction de la situation[100].
Vitesse

En général, il est difficile de déterminer la vitesse maximale des créatures marines éteintes. Pour les plésiosaures, cela est rendu plus difficile par le manque de consensus sur leur course de nageoires et leur démarche. Il n'y a pas de calculs exacts de leur nombre de Reynolds. Les empreintes fossiles montrent que la peau était relativement lisse, non écaillée, ce qui peut avoir réduit la traînée de forme[95]. De petites rides sont présentes dans la peau qui peuvent avoir empêché la séparation du flux laminaire dans la couche limite et ainsi réduit le frottement cutané.
La vitesse soutenue peut être estimée en calculant la traînée d'un modèle simplifié du corps, qui peut être approché par un sphéroïde allongé, et le niveau durable de production d'énergie par les muscles. Une première étude de ce problème a été publiée par Judy Massare en 1988[101]. Même en supposant une faible efficacité hydrodynamique de 0,65, le modèle de Massare semblait indiquer que les plésiosaures, s'ils étaient à sang chaud, auraient navigué à une vitesse de quatre mètres par seconde, soit environ quatorze kilomètres par heure, dépassant considérablement les vitesses connues des dauphins et les baleines existants[102]. Cependant, en 2002, Ryosuke Motani a montré que les formules utilisées par Massare étaient erronées. Un nouveau calcul, utilisant des formules corrigées, a abouti à une vitesse d'un demi-mètre par seconde (1,8 km/h) pour un plésiosaure à sang froid et d'un mètre et demi par seconde (5,4 km/h) pour un plésiosaure endothermique. Même l'estimation la plus élevée est environ un tiers inférieure à la vitesse des cétacés existants[103].
Massare a également essayé de comparer les vitesses des plésiosaures avec celles des deux autres principaux groupes de reptiles marins, les Ichthyosauria et les Mosasauridae. Elle a conclu que les plésiosaures étaient environ vingt pour cent plus lents que les ichtyosaures avancés, qui utilisaient un mouvement tunneliforme très efficace, oscillant juste la queue, mais cinq pour cent plus rapide que les mosasauridés, qui étaient supposés nager avec un mouvement anguilliforme inefficace, semblable à une anguille, de la corps[102].
Les nombreuses espèces de plésiosaures peuvent avoir différé considérablement dans leurs vitesses de nage, reflétant les différentes formes corporelles présentes dans le groupe. Alors que les « pliosauromorphes » à cou court (par exemple, le Liopleurodon) étaient peut-être des nageurs rapides, les « plésiosauromorphes » à long cou étaient davantage conçus pour la maniabilité que pour la vitesse, ralentis par une forte friction cutanée, mais capables d'un mouvement de roulement rapide. Certaines formes à long cou, comme les Elasmosauridae, ont également des nageoires trapues relativement courtes avec un faible rapport hauteur/largeur, ce qui réduit encore la vitesse mais améliore le roulis[104].
Plongée
Peu de données sont disponibles qui montrent exactement à quelle profondeur les plésiosaures ont plongé. Qu'ils aient plongé à une profondeur considérable est prouvé par des traces d'accident de décompression. Les têtes des humérus et des fémurs avec de nombreux fossiles montrent une nécrose du tissu osseux, causée par une remontée trop rapide après une plongée profonde. Cependant, cela ne permet pas de déduire une profondeur exacte car les dégâts pourraient avoir été causés par quelques plongées très profondes, ou bien par un grand nombre de descentes relativement peu profondes. Les vertèbres ne présentent pas de tels dommages : elles étaient probablement protégées par une irrigation sanguine supérieure, rendue possible par les artères pénétrant dans l'os par les deux foramens subcentralia, de grandes ouvertures dans leurs faces inférieures[105].
La descente aurait été aidée par une poussée d'Archimède négative, c'est-à-dire plus dense que l'eau. Bien sûr, cela aurait eu l'inconvénient de gêner la remontée. Les jeunes plésiosaures présentent une pachyostose, une densité extrême du tissu osseux, qui pourrait avoir augmenté le poids relatif. Les individus adultes ont un os plus spongieux. Les gastrolithes ont été suggérés comme méthode pour augmenter le poids[106] ou même comme moyen d'atteindre une flottabilité neutre, en les avalant ou en les crachant à nouveau au besoin[107]. Ils pourraient également avoir été utilisés pour augmenter la stabilité[108].
Les yeux relativement grands des Cryptocleididae ont été considérés comme une adaptation à la plongée profonde[109].
Rôle de queue
Une étude de 2020 a postulé que les sauroptérygiens s'appuyaient sur des coups de queue verticaux un peu comme les cétacés. Chez les plésiosaures, le tronc était rigide, donc cette action était plus limitée et en conjonction avec les nageoires[68].
Métabolisme
Traditionnellement, on supposait que les groupes de reptiles éteints étaient à sang froid comme les reptiles modernes. De nouvelles recherches au cours des dernières décennies ont conduit à la conclusion que certains groupes, tels que les dinosaures théropodes et les ptérosaures, étaient très probablement à sang chaud. Il est difficile de déterminer si les plésiosaures étaient également à sang chaud. L'une des indications d'un métabolisme élevé est la présence d'os fibrolamellaire à croissance rapide. Cependant, la pachyostose chez les individus juvéniles rend difficile d'établir si les plésiosaures possédaient un tel os. Cependant, il a été possible de vérifier sa présence avec des membres plus basaux du groupe plus inclusif auquel appartenaient les plésiosaures, les sauropterygia. Une étude en 2010 a conclu que l'os fibrolamellaire était à l'origine présent chez les sauroptérygiens[110]. Une publication ultérieure en 2013 a révélé que les Nothosauridae n'avaient pas ce type de matrice osseuse mais que les Pistosauria basaux en possédait, signe d'un métabolisme plus élevé[111]. Il est donc plus parcimonieux de supposer que les pistosauriens plus dérivés, les plésiosaures, avaient également un métabolisme plus rapide. Un article publié en 2018 a affirmé que les plésiosaures avaient des taux métaboliques au repos (RMR) dans l'aire de répartition des oiseaux sur la base d'une modélisation ostéohistologique quantitative[112]. Cependant, ces résultats sont problématiques au vu des principes généraux de la physiologie des vertébrés (voir la loi de Kleiber) ; les preuves des études isotopiques de l'émail des dents des plésiosaures suggèrent en effet une endothermie à des RMR inférieurs, avec des températures corporelles inférées d'env. 26 °C[113].
Reproduction

Comme les reptiles en général sont ovipares, jusqu'à la fin du vingtième siècle, il était possible que de plus petits plésiosaures aient pu ramper sur une plage pour pondre des œufs, comme les tortues modernes. Leurs membres forts et leur ventre plat semblaient avoir rendu cela possible. Cette méthode a été, par exemple, défendue par Halstead. Cependant, comme ces membres n'avaient plus d'articulations fonctionnelles du coude ou du genou et que le dessous, par sa planéité même, aurait généré beaucoup de frictions, déjà au XIXe siècle, on avait émis l'hypothèse que les plésiosaures avaient été vivipares. D'ailleurs, il était difficile de concevoir comment les plus grandes espèces, aussi grosses que des baleines, auraient pu survivre à un échouage. Les découvertes de fossiles d'embryons d'ichtyosaures ont montré qu'au moins un groupe de reptiles marins avait mis au monde des petits vivants. Le premier à affirmer que des embryons similaires avaient été trouvés chez des plésiosaures fut Harry Govier Seeley, qui déclara en 1887 avoir acquis un nodule avec quatre à huit minuscules squelettes[114]. En 1896, il décrit cette découverte plus en détail[115]. S'ils étaient authentiques, les embryons de plésiosaures auraient été très petits, comme ceux des ichtyosaures. Cependant, en 1982, Richard Anthony Thulborn montra que Seeley avait été trompé par un fossile « soigné » d'un nid d'écrevisses[116].
Un spécimen réel de plésiosaure trouvé en 1987 a finalement prouvé que les plésiosaures donnaient naissance à des jeunes vivants[117] : ce fossile d'un Polycotylus latippinus gravide montre que ces animaux ont donné naissance à un seul grand juvénile et ont probablement investi les soins parentaux dans leur progéniture, semblable à moderne baleines. Le jeune mesurait 1,5 mètre de long et était donc grand par rapport à sa mère de cinq mètres de long, indiquant une stratégie K en matière de reproduction[118]. On sait peu de choses sur les taux de croissance ou un possible dimorphisme sexuel.
Comportement social et intelligence
De la prise en charge parentale indiquée par la grande taille des jeunes, on peut déduire que le comportement social en général était relativement complexe[117]. On ne sait pas si les plésiosaures chassaient en meute. La taille relative de leur cerveau semble être typique des reptiles. Parmi les sens, la vue et l'odorat étaient importants, l'ouïe moins ; les élasmosauridés ont complètement perdu l'étrier. Il a été suggéré qu'avec certains groupes, le crâne abritait des organes électro-sensibles[119] - [120].
Paléopathologie
Certains fossiles de plésiosaures présentent des pathologies, résultat de maladie ou de vieillesse. En 2012, une mandibule de Pliosaurus a été décrite avec une articulation de la mâchoire clairement atteinte d'arthrite, signe typique de la sénescence[121].
Dans la culture contemporaine
_trailer_-_Plesiosaurus_1.png.webp)
Il a été suggéré que les légendes des serpents de mer et les observations modernes de supposés monstres dans les lacs ou la mer pourraient s'expliquer par la survie des plésiosaures dans les temps modernes. Cette proposition cryptozoologique a été rejetée par la communauté scientifique dans son ensemble, qui la considère comme basée sur la fantaisie et la pseudoscience. Il a été démontré que les prétendues carcasses de plésiosaures étaient des cadavres partiellement décomposés de requins pèlerins[122] - [123] - [124].
Alors que le monstre du Loch Ness est souvent décrit comme ressemblant à un plésiosaure, il est également souvent décrit comme ayant une apparence complètement différente. Un certain nombre de raisons ont été avancées pour qu'il soit peu probable qu'il s'agisse d'un plésiosaure. Ils incluent l'hypothèse que l'eau du loch est trop froide pour qu'un reptile présumé à sang froid puisse survivre facilement, l'hypothèse que les animaux à respiration aérienne seraient faciles à voir chaque fois qu'ils apparaissent à la surface pour respirer[125], le fait que le loch est trop petit et ne contient pas suffisamment de nourriture pour pouvoir nourrir une colonie de reproduction de grands animaux, et enfin le fait que le lac s'est formé il y a seulement 10 000 ans à la fin de la dernière période glaciaire, et la dernière apparition de fossiles de plésiosaures remonte à plus de 66 millions d'années[126]. Les explications fréquentes des observations incluent des vagues, des objets inanimés flottants, des tours de lumière, des animaux connus nageant et des farces[127]. Néanmoins, dans l'imaginaire populaire, les plésiosaures ont fini par être identifiés avec le monstre du Loch Ness. Cela a eu l'avantage de mieux faire connaître le groupe au grand public, mais l'inconvénient que les gens ont du mal à prendre le sujet au sérieux, obligeant les paléontologues à expliquer à maintes reprises que les plésiosaures ont réellement existé et ne sont pas simplement des créatures de mythe ou de fantaisie[128].
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- T. Lingham-Soliar, 1995: in Phil. Trans. Roy. Soc. Lond. 347: 155-180
- D. Cicimurri et M. Everhart, 2001: in Trans. Kansas. Acad. Sci. 104: 129-143
- F. R. O'Keefe, 2001: A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia) ; Acta Zool. Fennica 213: 1-63
- T. White, 1935: in Occasional Papers Boston Soc. Nat. Hist. 8: 219-228
- O. Hampe, 1992: Courier Forsch.-Inst. Senckenberg 145: 1-32
- R. Ellis, 2003: Sea Dragons' (Kansas University Press)
- 1997: in Reports of the National Center for Science Education, 17.3 (May/June 1997) p. 16–28.
- M.J. Everhart, 2005. "Where the Elasmosaurs roamed", Chapter 7 in "Oceans of Kansas: A Natural History of the Western Interior Sea," Indiana University Press, Bloomington, 322 p.
- M.J. Everhart, 2005. "Gastroliths associated with plesiosaur remains in the Sharon Springs Member (Late Cretaceous) of the Pierre Shale, Western Kansas" (on-line, updated from article in Kansas Acad. Sci. Trans. 103(1-2):58-69)
Voir aussi
Liens externes
- Ressources relatives au vivant :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- (en) Plesiosauria sur The Plesiosaur Directory
- (en) BBC NEWS | Science/Nature | Plesiosaur bottom-feeding shown
Notes et références
Notes
Références taxonomiques
- (en) Référence Paleobiology Database : Plesiosauria Blainville 1835 (consulté le )
Références
- (en) « The Plesiosaur Directory » [archive du ] (consulté le ).
- (en) M. Evans, Dinosaurs and Other Extinct Saurians: A Historical Perspective, Geological Society of London, , 5–31 p. (ISBN 978-1-86239-311-0).
- Richard Verstegan, 1605, A restitution of decayed intelligence or Nationum Origo, R. Bruney, Antwerpen
- Lhuyd, E., 1699, Lithophylacii Brittannici Ichnographia, sive Lapidum aliorumque Fossilium Brittanicorum singulari figurà insignium, Londen
- (en) W Stukeley, « An account of the impression of the almost entire sceleton of a large animal in a very hard stone, lately presented the Royal Society, from Nottinghamshire », Philosophical Transactions, vol. 30, no 360, , p. 963–968 (DOI 10.1098/rstl.1717.0053
 ).
). - Nicholls, J., 1795, The History and Antiquities of the County of Leicestershire. Volume I, John Nicholls, Londen
- (en) W.D. Conybeare, « Additional notices on the fossil genera Ichthyosaurus and Plesiosaurus », Transactions of the Geological Society of London, vol. 2, , p. 103–123 (DOI 10.1144/transgslb.1.1.103, S2CID 129545314, lire en ligne).
- (en) H.T. De la Beche et W.D. Conybeare, « Notice of the discovery of a new animal, forming a link between the Ichthyosaurus and crocodile, together with general remarks on the osteology of Ichthyosaurus », Transactions of the Geological Society of London, vol. 5, , p. 559–594.
- « Plesiosaur_Names », sur oceansofkansas.com.
- (en) W.D. Conybeare, « On the discovery of an almost perfect skeleton of the Plesiosaurus », Transactions of the Geological Society of London, vol. 2, , p. 382–389.
- (en) R.B.J. Benson, M. Evans, A.S. Smith, J. Sassoon, S. Moore-Faye, H.F. Ketchum et R. Forrest, « A giant pliosaurid skull from the Late Jurassic of England », PLOS ONE, vol. 8, no 5, , e65989 (PMID 23741520, PMCID 3669260, DOI 10.1371/journal.pone.0065989 , Bibcode 2013PLoSO...865989B).
- (en) T. H. Hawkins, « Memoirs on Ichthyosauri and Plesiosauri; Extinct monsters of the ancient Earth » [archive du ], Relfe and Fletcher, .
- (en) A. Peterson, « Terrible lizards and the wrath of God: How 19th century Christianity and Romanticism affected visual representations of dinosaurs and our perceptions of the ancient world », Stanford Undergraduate Research Journal, (lire en ligne).
- (en) T. H. Hawkins, The Book of the Great Sea-dragons, Ichthyosauri and Plesiosauri, Gedolim Taninum of Moses. Extinct Monsters of the Ancient Earth, W. Pickering, London, , 1–27 p..
- Christopher McGowan, 2001, The Dragon Seekers, Cambridge, Massachusetts, Perseus Publishing
- (en) R Owen, « Description of some remains of a gigantic crocodilian saurian, probably marine, from the Lower Greensand at Hythe and of teeth from the same formation at Maidstone, referable to the genus Polyptychodon », Proceedings of the Geologists' Association, vol. 3, , p. 449–452 (lire en ligne).
- (en) John Edward Gray, « A Synopsis of the Genera of Reptiles and Amphibia, with a Description of some new Species », Annals of Philosophy (British Museum), vol. 10, , p. 193–217.
- H. M. D. de Blainville, « Description de quelques espèces de reptiles de la Californie, précédée de l'analyse d'une système générale d'Erpetologie et d'Amphibiologie », Nouvelles Archives du Muséum d'Histoire Naturelle, vol. 4, , p. 233–296.
- (en) E.D. Cope, « [A resolution thanking Dr. Theophilus Turner for his donation of the skeleton of Elasmosaurus platyurus]. », Proc. Acad. Nat. Sci. Phila., vol. 20, , p. 314.
- (en) E.D. Cope, « Remarks on a new enaliosaurian, Elasmosaurus platyurus », Proceedings of the Academy of Natural Sciences of Philadelphia, vol. 20, , p. 92–93.
- (en) E.D. Cope, « On the reptilian orders Pythonomorpha and Streptosauria », Proceedings of the Boston Society of Natural History, vol. XII, , p. 250–266.
- (en) E.D. Cope, « On a new large enaliosaur », American Journal of Science Series, vol. 46, no 137, , p. 263–264.
- (en) E. D. Cope, Synopsis of the Extinct Batrachia and Reptilia of North America, Part I, vol. 14, Transactions of the American Philosophical Society, coll. « New Series », , 1–235 p., « Sauropterygia ».
- (en) J Leidy, « On the Elasmosaurus platyurus of Cope », American Journal of Science Series, vol. 49, no 147, , p. 392.
- (en) E.D. Cope, « Synopsis of the extinct Batrachia and Reptilia of North America », Transactions of the American Philosophical Society, vol. 14, no 1, , p. 1–252 (DOI 10.2307/1005355, JSTOR 1005355, lire en ligne).
- E.D. Cope, « On Elasmosaurus platyurus Cope », American Journal of Science Series, vol. 50, no 148, , p. 140–141.
- Ellis (2003), p. 129
- Williston, S.W., 1914, Water Reptiles of the Past and Present. Chicago University Press. Chicago, Illinois. 251 p.
- (en) J. P. Davidson, « Misunderstood Marine Reptiles: Late Nineteenth-Century Artistic Reconstructions of Prehistoric Marine Life », Transactions of the Kansas Academy of Science, vol. 118, nos 1–2, , p. 53–67 (DOI 10.1660/062.118.0107, S2CID 83904449).
- Smith, A.S., 2003, Cladistic analysis of the Plesiosauria (Reptilia: Sauropterygia). Masters thesis in palaeobiology, University of Bristol, 91 p.
- (en) Rodrigo A. Otero, Mario Suárez et Jacobus P. Le Roux, « First record of Elasmosaurid Plesiosaurs (Sauropterygia: Plesiosauria) in upper levels of the Dorotea Formation, Late Cretaceous (Maastrichtian), Puerto Natales, Chilean Patagonia », Andean Geology, vol. 36, no 2, , p. 342–350 (DOI 10.4067/s0718-71062009000200008 , lire en ligne).
- (en) Richard Forrest, « Liopleurodon » [archive du ], The Plesiosaur Site (consulté le ).
- (en) Richard Forrest, « The 'Monster of Aramberri' » [archive du ], The Plesiosaur Site (consulté le ).
- (en) Nigel Larkin, O'Connor, Sonia et Parsons, Dennis, « The virtual and physical preparation of the Collard plesiosaur from Bridgwater Bay, Somerset, UK », Geological Curator, vol. 9, no 3, , p. 107.
- (en) Richard Forrest, « The Collard Plesiosaur » [archive du ] (consulté le ).
- (en) Nigel Larkin, « Preparing and conserving an important six-foot long Plesiosaur skeleton for Somerset Museum » (consulté le ).
- Tamaki Sato, « A new Polycotylid Plesiosaur (Reptilia: Sauropterygia) from the Upper Cretaceous Bearpaw Formation in Saskatchewan, Canada. », Journal of Paleontology, vol. 79: 969-980, .
- « Hallazgo de un ejemplar completo de plesiosaurio joven » [archive du ] (consulté le ) (In Spanish)
- (en) H. Ledford, « Rare reptile fossil found in Antarctica », Nature News, , news061211–4 (DOI 10.1038/news061211-4, S2CID 85361720, lire en ligne).
- (en) « PREDATOR X - Naturhistorisk Museum » [archive du ], .
- (en) Katherine Hignett, « Plesiosaur: Ancient Sea Monster Discovered in Antarctica », Newsweek, (lire en ligne, consulté le ).
- (de) O. Rieppel, Sauropterygia I., vol. 12A, Verlag Dr. Friedrich Pfeil, coll. « Handbuch der Paläoherpetologie », , 1–134 p..
- (en) Y-N. Cheng, X-C. Wu et Q. Ji, « Chinese marine reptiles gave live birth to young », Nature, vol. 432, no 7015, , p. 383–386 (PMID 15549103, DOI 10.1038/nature03050, Bibcode 2004Natur.432..383C, S2CID 4391810).
- (en) G.W. Storrs, « Function and phylogeny in sauropterygian (Diapsida) evolution », American Journal of Science, vol. 293A, , p. 63–90 (DOI 10.2475/ajs.293.A.63, Bibcode 1993AmJS..293...63S).
- Rieppel, O., 1997, "Introduction to Sauropterygia", In: Callaway, J. M. & Nicholls, E. L. (eds.), Ancient marine reptiles p. 107–119. Academic Press, San Diego, California
- (en) F.R. O'Keefe, « The evolution of plesiosaur and pliosaur morphotypes in the Plesiosauria (Reptilia: Sauropterygia) », Paleobiology, vol. 28, , p. 101–112 (DOI 10.1666/0094-8373(2002)028<0101:teopap>2.0.co;2).
- (en) Roger B. J. Benson, Mark Evans et Patrick S. Druckenmiller, « High Diversity, Low Disparity and Small Body Size in Plesiosaurs (Reptilia, Sauropterygia) from the Triassic–Jurassic Boundary », PLOS ONE, vol. 7, no 3, , e31838 (PMID 22438869, PMCID 3306369, DOI 10.1371/journal.pone.0031838 , Bibcode 2012PLoSO...731838B).
- (en) H.F. Ketchum et R.B.J. Benson, « Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses », Biological Reviews of the Cambridge Philosophical Society, vol. 85, no 2, , p. 361–392 (PMID 20002391, DOI 10.1111/j.1469-185X.2009.00107.x, S2CID 12193439).
- (en) R.T. Bakker et Kauffman, E.G., Evolution of the Western Interior Basin, Geological Association of Canada, , 641–664 p., « Plesiosaur Extinction Cycles — Events that Mark the Beginning, Middle and End of the Cretaceous ».
- (en) P. S. Druckenmiller et A. P. Russell, « A phylogeny of Plesiosauria (Sauropterygia) and its bearing on the systematic status of Leptocleidus Andrews, 1922 », Zootaxa, vol. 1863, , p. 1–120 (DOI 10.11646/zootaxa.1863.1.1).
- (en) R. B. J. Benson et P. S. Druckenmiller, « Faunal turnover of marine tetrapods during the Jurassic-Cretaceous transition », Biological Reviews, vol. 89, no 1, , p. 1–23 (PMID 23581455, DOI 10.1111/brv.12038, S2CID 19710180).
- R. B. J. Benson et P. S. Druckenmiller, « Faunal turnover of marine tetrapods during the Jurassic-Cretaceous transition », Biological Reviews, vol. 89, no 1, , p. 1–23 (PMID 23581455, DOI 10.1111/brv.12038, S2CID 19710180).
- (en) L.B.H. Tarlo, « Stretosaurus gen nov., a giant pliosaur from the Kimmeridge Clay », Palaeontology, vol. 2, no 2, , p. 39–55.
- McHenry, Colin Richard (2009). "Devourer of Gods: the palaeoecology of the Cretaceous pliosaur Kronosaurus queenslandicus" (PDF): 1–460
- (en) R. B. J. Benson, M. Evans, A. S. Smith, J. Sassoon, S. Moore-Faye, H. F. Ketchum et R. Forrest, « A Giant Pliosaurid Skull from the Late Jurassic of England », PLOS ONE, vol. 8, no 5, , e65989 (PMID 23741520, PMCID 3669260, DOI 10.1371/journal.pone.0065989 , Bibcode 2013PLoSO...865989B).
- Caldwell, Michael W; 1997b. Modified perichondral ossification and the evolution of paddle-like limbs in Ichthyosaurs and Plesiosaurs; Journal of Vertebrate Paleontology 17 (3); 534-547
- Storrs, Glenn W.; 1990. Phylogenetic Relationships of Pachypleurosaurian and Nothosauriform Reptiles (Diapsida: Sauropterygia); Journal of Vertebrate Paleontology; 10 (Supplement to Number 3)
- (en) Adam Stuart Smith, « Fossils explained 54: Plesiosaurs », Geology Today, vol. 24, no 2, , p. 71–75 (DOI 10.1111/j.1365-2451.2008.00659.x).
- (en) Tai Kubo, Mark T. Mitchell et Donald M. Henderson, « Albertonectes vanderveldei, a new elasmosaur (Reptilia, Sauropterygia) from the Upper Cretaceous of Alberta », Journal of Vertebrate Paleontology, vol. 32, no 3, , p. 557–572 (DOI 10.1080/02724634.2012.658124, S2CID 129500470).
- (en) A.R.I. Cruickshank, P.G. Small et M.A. Taylor, « Dorsal nostrils and hydrodynamically driven underwater olfaction in plesiosaurs », Nature, vol. 352, no 6330, , p. 62–64 (DOI 10.1038/352062a0, Bibcode 1991Natur.352...62C, S2CID 4353612).
- D. S. Brown et A. R. I. Cruickshank, « The skull of the Callovian plesiosaur Cryptoclidus eurymerus and the sauropterygian cheek », Palaeontology, vol. 37, no 4, , p. 941–953.
- (en) M C. Buchy, E. Frey et S.W. Salisbury, « The internal cranial anatomy of the Plesiosauria (Reptilia, Sauropterygia): evidence for a functional secondary palate », Lethaia, vol. 39, no 4, , p. 289–303 (DOI 10.1080/00241160600847488).
- (de) F. von Huene, « Ein neuer Plesiosaurier aus dem oberen Lias Württembergs », Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg, vol. 79, , p. 1–21.
- (de) W Dames, « Die Plesiosaurier der Süddeutschen Liasformation », Abhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin, vol. 1895, , p. 1–81.
- Wilhelm, B.C., 2010, Novel anatomy of cryptoclidid plesiosaurs with comments on axial locomotion. Ph.D thesis, Marshall University, Huntington, WV. USA
- (en) B.C. Wilhelm et F. O'Keefe, « A new partial skeleton of Pantosaurus striatus, a cryptocleidoid Plesiosaur from the Upper Jurassic Sundance Formation of Wyoming », Journal of Vertebrate Paleontology, vol. 30, no 6, , p. 1736–1742 (DOI 10.1080/02724634.2010.521217, S2CID 36408899, lire en ligne).
- (en) Adam S. Smith, « Morphology of the caudal vertebrae in Rhomaleosaurus zetlandicus and a review of the evidence for a tail fin in Plesiosauria », Paludicola, vol. 9, no 3, , p. 144–158.
- (en) A. G. Sennikov, « Peculiarities of the Structure and Locomotor Function of the Tail in Sauropterygia », Biology Bulletin, vol. 46, no 7, , p. 751–762 (DOI 10.1134/S1062359019070100, S2CID 211217453).
- (en) C.R. McHenry, A.G. Cook et S. Wroe, « Bottom-feeding plesiosaurs », Science, vol. 310, no 5745, , p. 75 (PMID 16210529, DOI 10.1126/science.1117241, S2CID 28832109).
- « Plesiosaur bottom-feeding shown », BBC News, (lire en ligne, consulté le ).
- (de) J Geister, « Lebensspuren von Meersauriern und ihren Beutetieren im mittleren Jura (Callovien) von Liesberg, Schweiz », Facies, vol. 39, no 1, , p. 105–124 (DOI 10.1007/bf02537013, S2CID 127249009).
- (en) F. O'Keefe, R. Otero, S. Soto-Acuña, J. O'Gorman, S. Godfrey et S. Chatterjee, « Cranial anatomy of Morturneria seymourensis from Antarctica, and the evolution of filter feeding in plesiosaurs of the Austral Late Cretaceous », Journal of Vertebrate Paleontology, vol. 37, no 4, , e1347570 (DOI 10.1080/02724634.2017.1347570, S2CID 91144814).
- Chatterjee, S. and Small, B.J., 1989, "New plesiosaurs from the Upper Cretaceous of Antarctica", In: Crame, J. (ed) Origins and Evolution of Antarctic Biota, p. 197-215, Geological Society Publishing House, London
- (en) « The Plesiosaur Directory » (consulté le ).
- (en) J.A. Massare, « Ancient mariners », Natural History, vol. 101, , p. 48–53.
- (en) J A Massare, « Tooth morphology and prey preference of Mesozoic marine reptiles », J. Vertebr. Paleontol., vol. 7, no 2, , p. 121–137 (DOI 10.1080/02724634.1987.10011647).
- (en) M. J. Everhart, « Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the lamniform shark, Cretoxyrhina mantelli », Vertebrate Paleontology, vol. 2, no 2, , p. 14–24.
- (en) M. J. Everhart, « Plesiosaurs as the food of mosasaurs; new data on the stomach contents of a Tylosaurus proriger (Squamata; Mosasauridae) from the Niobrara Formation of western Kansas », The Mosasaur, vol. 7, , p. 41–46.
- Williston, Samuel Wendel; 1904. The stomach stones of the plesiosaurs Science 20; 565
- (en) M. J. Everhart, « Gastroliths associated with plesiosaur remains in the Sharon Springs Member of the Pierre Shale (Late Cretaceous), western Kansas », Kansas Acad. Sci. Trans, vol. 103, nos 1–2, , p. 58–69 (DOI 10.2307/3627940, JSTOR 3627940).
- (es) A Cerda et L Salgado, « Gastrolitos en un plesiosaurio (Sauropterygia) de la Formación Allen (Campaniano-Maastrichtiano), provincia de Río Negro, Patagonia, Argentina », Ameghiniana, vol. 45, , p. 529–536.
- (en) H.G. Seeley, « On Mauisaurus gardneri Seeley, an elasmosaurian from the base of the Gault of Folkestone », Quarterly Journal of the Geological Society of London, vol. 33, nos 1–4, , p. 541–546 (DOI 10.1144/gsl.jgs.1877.033.01-04.32, S2CID 128691833, lire en ligne).
- (en) S.P. Welles et J.D. Bump, « Alzadasaurus pembertoni, a new elasmosaur from the Upper Cretaceous of South Dakota », Journal of Paleontology, vol. 23, no 5, , p. 521–535.
- (en) M.J. Everhart, « Gastroliths associated with plesiosaur remains in the Sharon Springs Member of the Pierre Shale (late Cretaceous), Western Kansas », Kansas Academy of Sciences Transactions, vol. 103, nos 1–2, , p. 58–69.
- (de) E Fraas, « Reptilien und Säugetiere in ihren Anpassungserscheinungen an das marine Leben », Jahresheften des Vereins für vaterländische Naturkunde in Württemberg, vol. 29, , p. 347–386.
- (de) O Abel, « Die Anpassungsformen der Wirbeltiere an das Meeresleben », Schriften des Vereines zur Verbreitung Naturwissenschaftlicher Kenntnisse in Wien, vol. 48, no 14, , p. 395–422.
- (en) D.M.S. Watson, « The elasmosaurid shoulder-girdle and fore-limb », Proceedings of the Zoological Society of London, vol. 1924, no 2, , p. 885–917.
- (en) L.B. Tarlo, « The scapula of Pliosaurus macromerus Phillips », Palaeontology, vol. 1, , p. 193–199.
- (en) L.B. Halstead, « Plesiosaur locomotion », Journal of the Geological Society, vol. 146, no 1, , p. 37–40 (DOI 10.1144/gsjgs.146.1.0037, Bibcode 1989JGSoc.146...37H, S2CID 219541473).
- (en) J.A. Robinson, « The locomotion of plesiosaurs », Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, vol. 149, no 3, , p. 286–332.
- (en) J.A. Robinson, « Intercorporal force transmission in plesiosaurs », Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, vol. 153, no 1, , p. 88–128.
- (en) S. Tarsitano et J. Riess, « Plesiosaur locomotion — underwater flight versus rowing », Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, vol. 164, nos 1–2, , p. 193–194 (DOI 10.1127/njgpa/164/1982/188).
- (en) E. Frey et J. Reiss, « Considerations concerning plesiosaur locomotion », Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, vol. 164, nos 1–2, , p. 188–192 (DOI 10.1127/njgpa/164/1982/193).
- (en) Stephen J. Godfrey, « Plesiosaur subaqueous locomotion: a reappraisal », Neues Jahrbuch für Geologie und Paläontologie, vol. 1984, no 11, , p. 661–672 (DOI 10.1127/njgpm/1984/1984/661).
- (en) F. Sanders, K. Carpenter, B. Reed et J. Reed, « Plesiosaur swimming reconstructed from skeletal analysis and experimental results », Transactions of the Kansas Academy of Science, vol. 113, nos 1/2, , p. 1–34 (DOI 10.1660/062.113.0201, S2CID 86491931).
- Riess, J. and E. Frey, 1991. "The evolution of underwater flight and the locomotion of plesiosaurs", In: J.M.V. Rayner and R.J. Wootton (eds.) Biomechanics in Evolution, Cambridge, England: Cambridge University Press, p. 131-144
- (en) T. Lingham-Soliar, « Plesiosaur locomotion: Is the four-wing problem real or merely an atheoretical exercise? », Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, vol. 217, , p. 45–87 (DOI 10.1127/njgpa/217/2000/45).
- (en) F. Sanders, K. Carpenter, B. Reed et J. Reed, « Plesiosaur swimming reconstructed from skeletal analysis and experimental results », Journal of Vertebrate Paleontology, vol. 24, , p. 108A–109A (DOI 10.1080/02724634.2004.10010643, S2CID 220415208).
- (en) J. H. Long, J. Schumaker, N. Livingston et M. Kemp, « Four flippers or two? Tetrapodal swimming with an aquatic robot », Bioinspiration & Biomimetics, vol. 1, no 1, , p. 20–29 (PMID 17671301, DOI 10.1088/1748-3182/1/1/003, Bibcode 2006BiBi....1...20L).
- (en) Luke E. Muscutt, Gareth Dyke, Gabriel D. Weymouth, Darren Naish, Colin Palmer et Bharathram Ganapathisubramani, « The four-flipper swimming method of plesiosaurs enabled efficient and effective locomotion », Proceedings of the Royal Society B, vol. 284, no 1861, , p. 20170951 (PMID 28855360, PMCID 5577481, DOI 10.1098/rspb.2017.0951).
- (en) J.A. Massare, « Swimming capabilities of Mesozoic marine reptiles: implications for methods of predation », Paleobiology, vol. 14, no 2, , p. 187–205 (DOI 10.1017/s009483730001191x).
- Massare, J. A., 1994, "Swimming capabilities of Mesozoic marine reptiles: a review", In: L. Maddock et al. (eds.) Mechanics and Physiology of Animal Swimming, Cambridge, England: Cambridge University Press p. 133-149
- (en) R Motani, « Swimming speed estimation of extinct marine reptiles: energetic approach revisited », Paleobiology, vol. 28, no 2, , p. 251–262 (DOI 10.1666/0094-8373(2002)028<0251:sseoem>2.0.co;2).
- (en) F.R. O'Keefe, « Ecomorphology of plesiosaur flipper geometry », Journal of Evolutionary Biology, vol. 14, no 6, , p. 987–991 (DOI 10.1046/j.1420-9101.2001.00347.x, S2CID 53642687, CiteSeerx 10.1.1.579.4702, lire en ligne).
- (en) B.M. Rothschild et G.W. Storrs, « Decompression syndrome in plesiosaurs (Sauropterygia: Reptilia) », Journal of Vertebrate Paleontology, vol. 23, no 2, , p. 324–328 (DOI 10.1671/0272-4634(2003)023[0324:dsipsr]2.0.co;2).
- (en) M.A. Taylor, « Plesiosaurs — rigging and ballasting », Nature, vol. 290, no 5808, , p. 628–629 (DOI 10.1038/290628a0 , Bibcode 1981Natur.290..628T, S2CID 10700992).
- Taylor, M.A., 1993, "Stomach stones for feeding or buoyancy? The occurrence and function of gastroliths in marine tetrapods", Philosophical Transactions of the Royal Society of London B 341: 163–175
- (en) D.M. Henderson, « Floating point: a computational study of buoyancy, equilibrium, and gastroliths in plesiosaurs », Lethaia, vol. 39, no 3, , p. 227–244 (DOI 10.1080/00241160600799846).
- A. Yu. Berezin 2019 "Morphofunctional features of the plesiosaur Abyssosaurus nataliae (Plesiosauroidea: Plesiosauria) in connection with adaptations to a deep-water lifestyle." Ministry of National Resources and Ecology of the Russian Federation [in Russian].
- (en) N Klein, « Long Bone Histology of Sauropterygia from the Lower Muschelkalk of the Germanic Basin Provides Unexpected Implications for Phylogeny », PLOS ONE, vol. 5, no 7, , e11613 (PMID 20657768, PMCID 2908119, DOI 10.1371/journal.pone.0011613 , Bibcode 2010PLoSO...511613K).
- (en) Anna Krahl, Nicole Klein et P Martin Sander, « Evolutionary implications of the divergent long bone histologies of Nothosaurus and Pistosaurus (Sauropterygia, Triassic) », BMC Evolutionary Biology, vol. 13, , p. 123 (PMID 23773234, PMCID 3694513, DOI 10.1186/1471-2148-13-123).
- (en) Corinna V. Fleischle, Tanja Wintrich et P. Martin Sander, « Quantitative histological models suggest endothermy in plesiosaurs », PeerJ, vol. 6, , e4955 (ISSN 2167-8359, PMID 29892509, PMCID 5994164, DOI 10.7717/peerj.4955).
- (en) Aurélien Bernard, Christophe Lécuyer, Peggy Vincent, Romain Amiot, Nathalie Bardet, Eric Buffetaut, Gilles Cuny, François Fourel, François Martineau, Jean-Michel Mazin et Abel Prieur, « Regulation of Body Temperature by Some Mesozoic Marine Reptiles », Science, vol. 328, no 5984, , p. 1379–1382 (ISSN 1095-9203, PMID 20538946, DOI 10.1126/science.1187443, Bibcode 2010Sci...328.1379B, S2CID 206525584).
- (en) H.G. Seeley, « On the Mode of Development of the Young in Plesiosaurus », Report of the British Association for the Advancement of Science; Held at Manchester September, vol. 1887, , p. 697–698.
- Seeley, H. G.; 1896; "On a pyritous concretion from the Lias of Whitby which appears to show the external form of the body of embryos of a species of Plesiosaurus", Annual Report of Yorkshire philosophical Society p. 20-29
- (en) R.A. Thulborn, « Liassic plesiosaur embryos reinterpreted as shrimp burrows », Palaeontology, vol. 25, , p. 351–359.
- (en) F.R. O'Keefe et L.M. Chiappe, « Viviparity and K-Selected Life History in a Mesozoic Marine Plesiosaur (Reptilia, Sauropterygia) », Science, vol. 333, no 6044, , p. 870–873 (PMID 21836013, DOI 10.1126/science.1205689, Bibcode 2011Sci...333..870O, S2CID 36165835).
- (en) Jennifer Welsh, « Pregnant Fossil Suggests Ancient 'Sea Monsters' Birthed Live Young », LiveScience, (consulté le ).
- (en) J.P. O'Gorman et Z. Gasparini, « Revision of Sulcusuchus erraini (Sauropterygia, Polycotylidae) from the Upper Cretaceous of Patagonia, Argentina », Alcheringa, vol. 37, no 2, , p. 161–174 (DOI 10.1080/03115518.2013.736788, S2CID 131429825).
- (en) D. Foffa, J. Sassoon, A.R. Cuff, M.N. Mavrogordato et M.J. Benton, « Complex rostral neurovascular system in a giant pliosaur », Naturwissenschaften, vol. 101, no 5, , p. 453–456 (PMID 24756202, DOI 10.1007/s00114-014-1173-3, Bibcode 2014NW....101..453F, S2CID 7406418).
- (en) J. Sassoon, L.F. Noe et M.J. Benton, « Cranial anatomy, taxonomic implications and palaeopathology of an Upper Jurassic pliosaur (Reptilia: Sauropterygia) from Westbury, Wiltshire, UK », Palaeontology, vol. 55, no 4, , p. 743–773 (DOI 10.1111/j.1475-4983.2012.01151.x ).
- Anonymous (AP Report). Japanese scientist says that sea creature could be related to a shark species. The New York Times, 26 July 1977.
- (en) « Sea-Monster or Shark: An Alleged Plesiosaur Carcass », sur paleo.cc.
- Kimura S, Fujii K, and others. The morphology and chemical composition of the horny fiber from an unidentified creature captured off the coast of New Zealand. In CPC 1978, p. 67–74.
- The Great Sea Serpent, Antoon Cornelis Oudemans, 2009, Cosimo Inc (ISBN 978-1-60520-332-4) p. 321.
- (en) « Life », sur New Scientist.
- (en) « Crawley Creatures » [archive du ], sur crawley-creatures.com (consulté le ).
- Ellis (2003), p. 1–3.