Peloneustes
Peloneustes philarchus

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Sauropsida |
| Super-ordre | † Sauropterygia |
| Ordre | † Plesiosauria |
| Famille | † Pliosauridae |
| Clade | † Thalassophonea |
- Plesiosaurus philarchus Seeley, 1869
- Thaumatosaurus philarchus Lydekker, 1888
- Pliosaurus philarchus Seeley, 1892
- P. philarchus var. spathyrhynchus Linder, 1913
Peloneustes (littéralement « nageur dans la boue ») est un genre éteint de pliosaures ayant vécu durant le Jurassique moyen (Callovien) dans ce qui est aujourd'hui le Royaume-Uni. Tous les fossiles actuellement attribués au taxon proviennent du membre de Peterborough, dans la formation d'Oxford Clay, située en Angleterre. Il fut décrit à l'origine comme une espèce de Plesiosaurus par le paléontologue Harry Govier Seeley en 1869, avant de se voir attribuer son propre genre par le naturaliste Richard Lydekker en 1889. Alors que de nombreuses espèces ont été attribuées à Peloneustes, Peloneustes philarchus est actuellement la seule considérée comme valide, les autres ayant été déplacées vers des genres différents, considérés comme douteux ou synonymes de P. philarchus. Une partie du matériel anciennement attribué à une deuxième espèce nommée Peloneustes evansi, a depuis été réattribué à "Pliosaurus" andrewsi, un taxon lui-même incertain. Peloneustes est connu à partir de nombreux spécimens fossiles, incluant du matériel très complet.
Avec une taille maximale estimée entre 3,5 et 4 m de long, Peloneustes n'est pas un grand représentant des pliosauridés. Ce reptile marin a un grand crâne de forme triangulaire qui occupe environ un cinquième de la longueur du corps. L'avant du crâne est allongé en un rostre étroit, constituant le museau. La symphyse mandibulaire, là où les extrémités situées sur l’avant de chaque côté de la mandibule fusionnent, est allongée chez Peloneustes et contribue à renforcer la mâchoire. Une crête surélevée est située entre les rangées de dents au niveau de la symphyse mandibulaire. Les dents de Peloneustes sont coniques et ont des sections transversales circulaires, portant des arêtes verticales sur tous les côtés. Les dents situées à l’avant sont plus grandes que les dents à l'arrière. Avec seulement 19 à 21 vertèbres cervicales, Peloneustes a un cou relativement court par rapport aux autres plésiosaures. Les membres de Peloneustes sont en forme de nageoires, les palettes natatoires arrières étant plus grandes que celles situées en avant.
Peloneustes est considéré à la fois comme un proche parent de Pliosaurus ou comme un pliosauridé plus basal au sein du clade des Thalassophonea, cette dernière interprétation trouvant le plus de soutien parmi les paléontologues. Comme les autres plésiosaures, Peloneustes fut bien adapté à la vie aquatique, utilisant ses palettes natatoires pour une méthode de nage connue sous le nom de « vol sous-marin ». Les crânes des pliosauridés sont renforcés pour mieux résister aux contraintes de leur alimentation. Le museau long et étroit de Peloneustes aurait pu être balancé rapidement sous l'eau pour attraper des poissons, qu'il aurait percé grâce à ses nombreuses dents acérées. D'après les archives fossiles, Peloneustes habitait une mer intérieure d'environ 30 à 50 m de profondeur. Il partageait son habitat avec une variété d'autres animaux, notamment des invertébrés, des poissons, des thalattosuchiens, des ichtyosaures et d'autres plésiosaures. Au moins cinq autres pliosauridés sont connus du membre de Peterborough, mais leur anatomie est assez variée, ce qui indique qu'ils se seraient alimentés à partir de différentes sources de nourriture, évitant ainsi la concurrence.
Historique des recherches
Premières découvertes
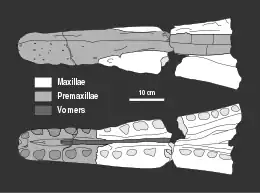
Les strates du membre de Peterborough de la formation d'Oxford Clay sont depuis longtemps exploitées pour la fabrication de briques. Depuis la fin du XIXe siècle, lorsque les opérations d'excavation commencent, des fossiles de nombreux animaux marins sont extraits des roches[1]. Parmi ceux-ci se trouve le spécimen qui allait devenir l'holotype de Peloneustes philarchus, découvert par le géologue Henry Porter dans une argilière située près de Peterborough, en Angleterre. Le spécimen comprend une mandibule, la partie avant de la mâchoire supérieure, diverses vertèbres de tout le corps, des éléments de la ceinture scapulaire et du bassin, des humérus, des fémurs et divers autres os des membres[2]. En 1866, le géologue Adam Sedgwick achète le spécimen pour le musée de l'université de Cambridge[1], qui par la suite le catalogue sous le numéro CAMSM J.46913 et le stocke dans la salle de cours de l'université au sein du cabinet D[1] - [2]. Le paléontologue Harry Govier Seeley décrit le spécimen en 1869 comme une nouvelle espèce du genre Plesiosaurus, sous le nom de Plesiosaurus philarchus[2]. L'épithète spécifique philarchus vient du grec ancien φιλία / philía « aimant » et ἄρχω / árkhō « commander » ou « régir ». Bien qu'aucune description du sens de cet étymologie n'ait été donnée par Seeley, il est suggéré que ce nom signifie « aimant le pouvoir », probablement en raison de son crâne large et imposant[3]. Dans son ouvrage, Seeley ne décrit pas ce spécimen en détail, donnant principalement une liste du matériel connu[2]. Alors que des publications ultérieures décrivaient plus en détail ces restes, CAMSM J.46913 reste alors mal décrit[1].
Les frères Alfred (en) et Charles Leeds collectaient des fossiles d'Oxford Clay depuis environ 1867, encouragés par le géologue John Phillips de l'université d'Oxford, assemblant ce qui est devenu la collection Leeds. Alors que Charles est finalement parti, Alfred, qui a collecté la majorité des spécimens, continue à collecter des fossiles jusqu'en 1917. Finalement, après une visite d'Henry Woodward du musée d'histoire naturelle de Londres à la collection de Leeds à Eyebury en 1885, le musée achète environ 5 tonnes de fossiles en 1890. Cela apporte une plus grande renommée à la collection de Leeds, et il vendra plus tard des spécimens à différents musées à travers l'Europe, et même certains aux États-Unis[4]. Le matériel soigneusement préparé est généralement en bon état, bien qu'il ait été assez souvent écrasé et brisé par les processus géologiques, en particulier le matériel crânien[5]:V-VI.
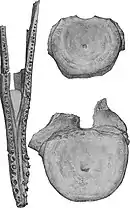
Le naturaliste Richard Lydekker a été informé de la présence d'un squelette de plésiosaure au musée d'histoire naturelle de Londres par le géologue George Charles Crick (en), qui y travaillait alors sur le spécimen, catalogué NHMUK R1253[1], découvert dans la formation d'Oxford Clay à Green End, Kempston, près de Bedford. Alors que Lydekker émet l'hypothèse que le squelette pourrait être complet, il a toutefois été endommagé lors de l'excavation. Les ceintures des membres avaient été fortement fragmentées lorsque le spécimen est arrivé au musée, mais un ouvrier du nom de Lingard du département de géologie réussit à en restaurer une grande partie. En plus des ceintures des membres, le spécimen se compose également d'une mandibule partielle, de dents, de plusieurs vertèbres (bien qu'aucune du cou) et d'une grande partie des membres. Lydekker identifie ce spécimen comme un individu de Plesiosaurus philarchus et en publie une description en 1889. Après avoir étudié ce spécimen et d'autres de la collection Leeds, il en conclu que les plésiosaures au cou raccourci et à la tête large ne pouvaient pas être classés comme des espèces de Plesiosaurus, ce qui signifie que "P." philarchus appartient à un genre différent. Il l'attribua initialement à Thaumatosaurus en 1888[6], mais décida plus tard qu'il est suffisamment distinct pour justifier son propre genre, qu'il nomme Peloneustes dans sa publication de 1889[7]. Le nom générique Peloneustes vient du grec ancien πηλός / pēlós signifiant « boue » ou « argile », en référence à la formation d'Oxford Clay, et νηκτός / nêktos, voulant dire « nageur »[3]. Cependant, Seeley regroupe Peloneustes dans Pliosaurus en 1892, affirmant que les deux n'étaient pas suffisamment différents pour justifier des genres séparés[8]. Seeley et Lydekker n'ont pas pu s'entendre dans quel taxon fallait classer P. philarchus, ce qui représentait une partie d'une querelle entre les deux scientifiques. Cependant, Peloneustes est depuis devenu le nom accepté[6].
La collection Leeds contenait plusieurs spécimens de Peloneustes[9]:63-70. En 1895, le paléontologue Charles William Andrews décrit l'anatomie du crâne de Peloneustes sur la base de quatre crânes partiels de la collection Leeds[10]. En 1907, le géologue Frédéric Jaccard publie une description concernant deux spécimens de Peloneustes d'Oxford Clay près de Peterborough, conservés au musée cantonal de géologie de Lausanne, en Suisse. Le plus complet des deux spécimens comprend un crâne complet préservant les deux mâchoires ; plusieurs dents isolées ; 13 vertèbres cervicales, 5 vertèbres pectorales et 7 vertèbres caudales ; des côtes ; les deux omoplates, une coracoïde ; une interclavicule partielle ; un bassin complet à l'exception d'un ischion ; et les quatre membres, qui sont presque complets. L'autre spécimen conserve 33 vertèbres et quelques côtes associées. Étant donné que le spécimen décrit par Lydekker avait besoin d'être restauré et que les informations manquantes avaient été complétées par des données d'autres spécimens dans sa publication, Jaccard juge pertinent de publier une description contenant des photographies du spécimen le plus complet à Lausanne pour mieux illustrer l'anatomie de Peloneustes[11].
En 1913, le naturaliste Hermann Linder décrit plusieurs spécimens de P. philarchus hébergés à l'Institut für Geowissenschaften, à l'université de Tübingen et au musée national d'histoire naturelle de Stuttgart, en Allemagne. Ces spécimens proviennent également de la collection Leeds[1]. Parmi les spécimens qu'il décrit de l'ancienne institution, il y a un squelette monté presque complet, dépourvu de deux vertèbres cervicales, de quelques vertèbres caudales à l'extrémité de la queue et de quelques phalanges distales. Seule la partie arrière du crâne est en bon état, ainsi qu’une mandibule qui est en grande partie intacte. Un autre des spécimens décrits par Linder est un crâne bien conservé (catalogué GPIT RE/3409)[1], également de l'université de Tübingen, préservant un anneau sclérotique, étant la quatrième fois que ces os sont signalés chez un plésiosaure[12].
.png.webp)
Andrews décrit plus tard les spécimens de reptiles marins de la collection Leeds qui se trouvent au musée d'histoire naturelle de Londres, publiant deux volumes, l'un en 1910 et l'autre en 1913. L'anatomie des spécimens de Peloneustes est décrite dans le deuxième volume, basée principalement sur les crânes bien conservés NHMUK R2679, NHMUK R3808 et NHMUK R3318, un squelette presque complet. Le spécimen NHMUK R3318 est si bien conservé qu'il a pu être réarticulé et monté, bien que les parties manquantes du bassin et des membres aient dû être remplies. Le squelette monté a été exposé dans la galerie des reptiles fossiles du musée[5]:IX - [9]:35, 63. Andrews décrit cette monture en 1910, remarquant qu'il s'agissait de la première monture squelettique d'un pliosauridé, fournissant ainsi des informations importantes sur l'anatomie globale du groupe[13].
En 1960, le paléontologue Lambert Beverly Tarlo publie une revue des espèces de pliosauridés qui ont été signalées dans les archives fossiles du Jurassique supérieur. De nombreuses espèces de pliosauridés avaient été nommées sur la base de fragments isolés, créant une confusion. Tarlo constate également que des descriptions inexactes du matériel et des paléontologues ignorant le travail d'autres ne faisaient qu'accentuer cette confusion. Sur les 36 espèces qu'il examine, il n'en trouve que neuf valides, dont P. philarchus[6]. En 2011, les paléontologues Hilary Ketchum et Roger Benson décrivent l'anatomie du crâne de Peloneustes. Depuis les précédentes études anatomiques d'Andrews et Linder, d'autres spécimens ont été trouvés, dont NHMUK R4058, un crâne très bien conservé, donnant une meilleure idée de la forme du crâne[1].
Autres espèces attribuées
De nombreuses autres espèces ont été attribuées à Peloneustes tout au long de son histoire, mais elles ont toutes été réaffectées à différents genres ou considérées comme invalides[1]. Dans la même publication dans laquelle il nomme P. philarchus, Seeley nomme également une autre espèce de Plesiosaurus, P. sterrodeirus, sur la base de sept spécimens du Woodwardian Museum constitués de matériel crânien et vertébral[2]. Lorsque Lydekker érige le genre Peloneustes pour P. philarchus, il reclasse également "Plesiosaurus" sterrodeirus et "Pleiosaurus" aequalis (une espèce nommée par John Phillips en 1871)[14]:365 - [6] comme des représentants de ce genre[7]. Dans son examen de 1960 de la taxonomie des pliosauridés, Tarlo considère P. aequalis comme invalide, car il est basé sur des propodiums, des os du membre supérieur, qui ne peuvent pas être utilisés pour différencier les différentes espèces de pliosauridés. Il considère que Peloneustes sterrodeirus appartient plutôt à Pliosaurus, peut-être au sein de P. brachydeirus[6].
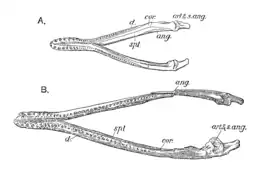
Une autre des espèces décrites par Seeley en 1869 est Pliosaurus evansi, basée sur des spécimens du Woodwardian Museum[2]. Ceux-ci se composent de vertèbres cervicales et dorsales, de côtes et d'une coracoïde. En raison du fait qu'il s'agit d'une espèce plus petite de Pliosaurus et de sa similitude avec Peloneustes philarchus, Lydekker le réaffecte au genre Peloneustes en 1890, notant qu'il est plus grand que Peloneustes philarchus[15]. Il pense également qu'une grande mandibule et une palette natatoire attribuées à Pleiosaurus? grandis par Phillips en 1871[14]:318 appartiennent plutôt à cette espèce[6]. En 1913, Andrews attribue un squelette partiel d'un autre grand pliosaure trouvé par Leeds à Peloneustes evansi, notant que si la mandibule et les vertèbres sont similaires à d'autres spécimens de cette espèce nommée, elles restent assez différentes de celles de Peloneustes philarchus. Par conséquent, Andrews considère qu'il est possible que P. evansi appartienne réellement à un genre distinct qui aurait été morphologiquement intermédiaire entre Peloneustes et Pliosaurus[9]:72. Dans sa revue de 1960 sur les pliosauridés, Tarlo synonymise Peloneustes evansi avec Peloneustes philarchus en raison de l'identité de leurs vertèbres cervicales (à l'exception d'une différence de taille). Il considère les plus gros spécimens de Peloneustes evansi comme distincts et les attribue à une nouvelle espèce de Pliosaurus, P. andrewsi (bien que cette espèce ne soit actuellement plus considérée comme appartenant à Pliosaurus)[6] - [16]. Ketchum et Benson ne sont pas d'accord avec cette synonymie et estiment en 2011 que l'holotype de Peloneustes evansi manque de traits distinctifs, restant alors un nomen dubium et donc un pliosauridé indéterminé[1].
En 1905, le paléontologue E. Koken décrit ce qu'il identifie comme une autre espèce de Peloneustes, P. kanzleri, datant du Crétacé d'Allemagne[1]. En 1960, Tarlo réidentifie cette espèce comme un élasmosauridé[6]. En 1913, Linder créé une sous-espèce de Peloneustes, P. philarchus var. spathyrhynchus, en le différenciant par sa symphyse mandibulaire spatulée (là où les deux côtés de la mandibule se rencontrent et fusionnent)[12]. Tarlo le considère comme un synonyme de Peloneustes philarchus en 1960[6], constatant que la symphyse mandibulaire de Peloneustes est proportionnellement plus large chez les spécimens plus grands, ce qui rend ce trait plus susceptible d'être dû à une variation intraspécifique. Les fossiles, étants broyés par les processus géologiques, rendent toutefois difficile la mesure précise de ces proportions[1]. En 1948, le paléontologue Nestor Novozhilov nomme une nouvelle espèce de Peloneustes, P. irgisensis, basée sur le spécimen PIN 426, un squelette partiel composé d'un grand crâne incomplet, de vertèbres et d'un membre postérieur partiel, avec le contenu de l'estomac préservé. Le spécimen a été déterré dans le bassin inférieur de la Volga en Russie[17] - [18]. Dans sa revue de 1960, Tarlo considère cette espèce comme étant trop différente de Peloneustes philarchus pour appartenir à ce genre, le plaçant provisoirement dans Pliosaurus. Il émet l'hypothèse que Novozhilov avait pensé à tort que Peloneustes était le seul pliosauridé à long museau, d'où l'affectation initiale[6]. En 1964, Novozhilov érige un nouveau genre pour cette espèce, Strongylokrotaphus, mais d'autres études concordant avec celles de Tarlo réaffectent l'espèce à Pliosaurus, voire peut-être même comme un synonyme de Pliosaurus rossicus. À ce moment-là, le spécimen PIN 426 avait subi de lourds dégâts de pyrite[18] - [16].

En 1998, le paléontologue Frank Robin O'Keefe propose qu'un spécimen de pliosauridé datant du Jurassique inférieur des argiles à Posidonies d'Allemagne pourrait représenter une nouvelle espèce de Peloneustes. En 2001, il le considère comme appartenant à un genre à part, le nommant Hauffiosaurus zanoni[1] - [19]. Les paléontologues Zulma Gasparini et Manuel A. Iturralde-Vinent attribuent un pliosauridé d'une formation datant du Jurassique supérieur de Cuba à Peloneustes sp. en 2006[20]. En 2009, Gasparini le redécrit sous le nom Gallardosaurus iturraldei[21]. En 2011, Ketchum et Benson considèrent que Peloneustes ne contient actuellement qu'une seule espèce valide, P. philarchus. Ils reconnaissent 21 spécimens précis de Peloneustes philarchus, tous provenant du membre de Peterborough de la formation d'Oxford Clay. Ils considèrent que certains spécimens du membre de Peterborough et de Marquise en France, précédemment affectés à Peloneustes, appartiennent à différents pliosauridés actuellement non nommés[1].
Description
Description simplifié

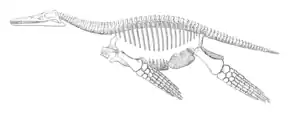
Peloneustes est un représentant de taille petite à moyenne des Pliosauridae[9]:34 - [22]:12. Le spécimen NHMUK R3318, le squelette monté du musée d'histoire naturelle de Londres, mesure 3,5 m de long[13], tandis que le squelette monté de l'Institut für Geowissenschaften de l'université de Tübingen mesure 4,05 m de long[12]. Les plésiosaures peuvent généralement être décrits comme étant du morphotype « plésiosauromorphe », à petite tête et à long cou, ou du morphotype « pliosauromorphe », à grosse tête et à cou court[23]. Peloneustes est un représentant de ce dernier morphotype[23], avec son crâne représentant un peu moins d'un cinquième de la longueur totale de l'animal. Peloneustes, comme tous les plésiosaures, a une queue courte, un tronc massif et tous ses membres modifiés en forme de grandes nageoires[22]:13.
Crâne
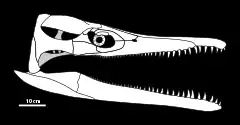
Alors que l'holotype de Peloneustes n'a pas la partie arrière de son crâne, de nombreux spécimens supplémentaires bien conservés, dont un n'étant pas écrasé de haut en bas, sont attribués au genre. Ces crânes varient en taille, mesurant 60 à 78,5 cm de long. Le crâne de Peloneustes est allongé et incliné vers le haut au niveau de son extrémité arrière[1]. Vu de dessus, le crâne a la forme d'un triangle isocèle[9]:35, avec l'arrière du crâne élargie et l'avant allongé en un rostre étroit. La partie la plus en arrière du crâne a des côtés à peu près parallèles, contrairement aux régions avant effilées. Les narines externes sont petites et situées vers le milieu du crâne. Les orbites en forme de rein sont tournées vers l'avant et vers l'extérieur et sont situées sur la moitié arrière du crâne. Les anneaux sclérotiques sont composés d'au moins 16 osselets individuels, un nombre inhabituellement élevé pour un reptile. Les fosses temporales sont élargies et elliptiques, et situées sur le quart le plus en arrière du crâne[1].
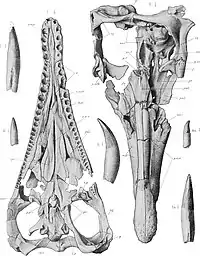
De manière caractéristique, les prémaxillaires de Peloneustes portent six dents chacun, et les diastèmes de la mâchoire supérieure sont étroits. Alors qu'il a été affirmé que Peloneustes aurait eu des os nasaux, des spécimens bien conservés indiquent que ce n'est pas le cas. Les frontaux de Peloneustes contactent à la fois les orbites et les narines externes, un trait distinctif de Peloneustes. Il y a eu une certaine incertitude quant à savoir si Peloneustes avait ou non des os lacrymaux en raison de la mauvaise conservation de certains fossiles. Cependant, des spécimens bien conservés indiquent que les os lacrymaux sont des os distincts comme chez les autres pliosauridés, par opposition aux extensions des os jugales. Le palais de Peloneustes est plat et porte de nombreuses ouvertures, incluant les narines internes. Ces ouvertures entrent en contact par des os palatins, une configuration utilisée pour identifier ce genre. Le parasphénoïde, un os qui forme la partie avant inférieure du neurocrâne, porte un long processus cultriforme qui est visible lorsque le palais est vu de dessous, une autre caractéristique distinctive de Peloneustes. L'os occipital de Peloneustes est ouvert, portant de grandes fosses[1].
Peloneustes est connu pour de nombreuses mandibules, dont certaines sont bien conservées. La plus longue d'entre elles mesure 87,7 cm. La symphyse mandibulaire est allongée, représentant environ un tiers de la longueur totale de la mandibule. Derrière la symphyse, les deux côtés de la mandibule divergent avant de se recourber délicatement vers l'intérieur et près de l'extrémité postérieure. Chaque os dentaire, l'os portant les dents dans la mandibule chez les diapsides, a entre 36 et 44 dents, dont 13 à 15 sont situées sur la symphyse. Les deuxième à septième alvéoles dentaires sont plus grandes que celles situées plus en arrière, et la symphyse mandibulaire est plus large autour des cinquième et sixième. En plus des caractéristiques des dents de la mandibule, Peloneustes peut également être identifié par ses processus coronoïdes, qui contribuent à la symphyse mandibulaire. Entre les rangées de dents, la symphyse mandibulaire porte une crête surélevée où les os dentaires se rencontrent. Il s'agit d'une caractéristique unique de Peloneustes, que l'on ne retrouve chez aucun autre plésiosaure. La glène mandibulaire, la prise de l'articulation de la mâchoire, est large, en forme de rein et inclinée vers le haut et vers l'intérieur[1].

Les dents de Peloneustes ont des sections transversales circulaires, comme on le voit chez d'autres pliosauridés de son époque[6]. Les dents ont la forme de cônes recourbés. L'émail des couronnes porte des crêtes verticales régulièrement espacées de longueur variable sur tous les côtés. Ces crêtes sont plus concentrées sur le bord concave des dents. La plupart des crêtes s'étendent de la moitié aux deux tiers de la hauteur totale de la couronne, peu atteignant réellement le foramen apical de la dent[1]. La dentition de Peloneustes est hétérodonte, c'est-à-dire que les dents sont de formes différentes. Les plus grandes dents sont caniniformes et sont situées à l'avant des mâchoires, tandis que les petites dents sont plus nettement recourbées[1], robustes, et situées plus en arrière[24].
Squelette postcrânien
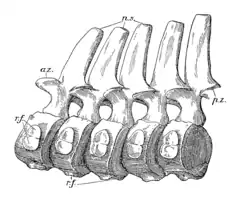
En 1913, sur la base de spécimens de la collection Leeds, Andrews rapporte que Peloneustes aurait eu 21 à 22 vertèbres cervicales, 2 à 3 vertèbres pectorales et environ 20 vertèbres dorsales, le nombre exact de vertèbres sacrées et caudales étant inconnu[9]:47, 52. Cependant, la même année, Linder signale 19 vertèbres cervicales, 5 pectorales, 20 dorsales, 2 sacrées et au moins 17 vertèbres caudales à Peloneustes, sur la base d'un spécimen de l'Institut für Geowissenschaften, à l'université de Tübingen[12] - [1]. Les deux premières vertèbres cervicales, l'atlas et l'axis, sont fusionnées chez les adultes, mais chez les juvéniles, elles sont présentes sous forme de plusieurs éléments non fusionnés[9]:47. L'intercentrum (partie du corps vertébral) de l'axis est à peu près rectangulaire, s'étendant sous le centrum (corps vertébral) de l'atlas[1]. Les vertèbres cervicales portent de hautes épines neurales qui sont comprimées d'un côté à l'autre[1] - [9]:50. Les centra cervicaux sont environ deux fois moins longs que larges. Ils portent des surfaces articulaires fortement concaves, avec un rebord proéminent autour du bord inférieur des vertèbres situées vers l'avant de la série. Chaque centrum cervical a une quille solide le long de la ligne médiane de sa face inférieure[6]. La plupart des côtes cervicales portent deux têtes séparées par une encoche[9]:53.
Les vertèbres pectorales portent des articulations pour leurs côtes respectives, partiellement sur leurs arcs central et neural. Après ces vertèbres se trouvent les vertèbres dorsales, qui sont plus allongées que les vertèbres cervicales et ont des épines neurales plus courtes. Les vertèbres sacrées et caudales ont toutes deux des centra moins allongés qui sont plus larges que hauts. De nombreuses côtes de la hanche et de la base de la queue portent des extrémités externes élargies qui semblent s'articuler les unes avec les autres. Andrews émet l'hypothèse en 2013 que cette configuration aurait raidi la queue, peut-être pour soutenir les gros membres postérieurs. Les dernières vertèbres caudales diminuent fortement de taille et auraient supporté des chevrons proportionnellement plus grands que les vertèbres caudales situées plus en avant. Andrews émet également l'hypothèse que cette morphologie était peut-être présente pour soutenir une petite structure en forme de nageoire caudale[9]:52-53. D'autres plésiosaures auraient eux des nageoires caudales, avec des impressions d'une telle structure peut-être connues chez une espèce[25].
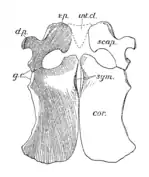
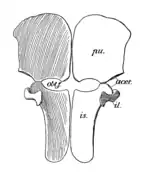
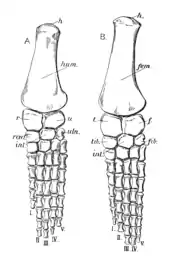
La ceinture scapulaire de Peloneustes est grande, mais pas aussi lourdement construite que chez certains autres plésiosaures. Les coracoïdes sont les plus gros os de la ceinture scapulaire et ont la forme d'une plaque. L'articulation de l'épaule est formée à la fois par l'omoplate et le coracoïde, les deux os formant un angle de 70° l'un avec l'autre. Les omoplates sont de forme typique pour un pliosauridé et triradié, portant trois saillies proéminentes. La branche dorsale est dirigée vers l'extérieur, vers le haut et vers l'arrière[9]:55 - [6]. Le dessous de chaque omoplate porte une crête dirigée vers le bord avant de sa branche ventrale[6]. Les branches ventrales des deux omoplates sont séparées l'une de l'autre par un os triangulaire appelé interclavicule. Comme on le voit chez d'autres pliosaures, le bassin de Peloneustes porte des ischions et des os pubiens larges et plats. Le troisième os pelvien, l'ilium, est plus petit et allongé, s'articulant avec l'ischion. L'extrémité supérieure de l'ilium présente une grande variation au sein de P. philarchus, avec deux formes connues, l'une avec un bord supérieur arrondi, l'autre avec un bord supérieur plat et une forme plus anguleuse[9]:55-56, 58-60.
Les membres postérieurs de Peloneustes sont plus longs que ses membres antérieurs, le fémur étant plus long que l'humérus, bien que l'humérus soit le plus robuste des deux os[9]:57, 60. Le radius est approximativement aussi large que long, contrairement à l'ulna, qui est plus large que long[6]. Le radius est le plus grand de ces deux os[9]:58. Le tibia est plus gros que le péroné et est plus long que large, tandis que le péroné est plus large que long chez certains spécimens[6]. Les métacarpiens, les métatarsiens et les phalanges manuelles proximales sont aplatis. La plupart des phalanges des deux membres ont des sections transversales arrondies et toutes ont des constrictions proéminentes au milieu. Le nombre de phalanges dans chaque doigt est inconnu dans les membres antérieurs et postérieurs[9]:58, 62.
Classification

Seeley décrit initialement Peloneustes comme une espèce de Plesiosaurus[2], ce qui fut une pratique assez courante, la portée des genres étant similaire à ce qui est actuellement utilisé pour les familles lors de cet époque[22]:7. En 1874, Seeley nomme une nouvelle famille de plésiosaures, Pliosauridae, pour contenir des représentants similaires à Pliosaurus[26]. En 1890, Lydekker place Peloneustes dans cette famille[15], auquel il a toujours été attribué depuis[8] - [9]:1 - [6] - [1]. La relation exacte entre les pliosauridés et les autres plésiosaures est incertaine. En 1940, le paléontologue Theodore E. White considère les pliosauridés comme des proches parents des Elasmosauridae sur la base de l'anatomie des épaules[27]. Cependant, le paléontologue Samuel Paul Welles, pense que les pliosauridés ressemblent davantage aux Polycotylidae, car les deux taxons ont tous de grands crânes et des cous courts, entre autres caractéristiques. Il regroupe ces deux familles dans la super-famille des Pliosauroidea, avec d'autres plésiosaures formant la super-famille des Plesiosauroidea[28]. Une autre famille de plésiosaures, Rhomaleosauridae, a depuis été attribuée aux Pliosauroidea[29] - [19], tandis que les Polycotylidae ont été réaffecté aux Plesiosauroidea[30] - [31]. En 2012, Benson et ses collègues récupèrent une topologie différente, les Pliosauridae s’avérant êtres plus étroitement liés aux Plesiosauroidea qu'aux Rhomaleosauridae. Ce clade réunissant les pliosauridés et les plésiosauroïdes est nommé Neoplesiosauria[31].
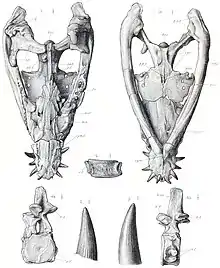
Au sein des Pliosauridae, la position phylogénétique exacte de Peloneustes est incertaine[1]. En 1889, Lydekker considère que Peloneustes représenterait une forme transitionnelle situé entre Pliosaurus et les plésiosaures antérieurs, bien qu'il trouve peu probable que Peloneustes soit l'ancêtre de Pliosaurus[7]. En 1960, Tarlo considère Peloneustes comme un proche parent de Pliosaurus, puisque les deux taxons ont des symphyses mandibulaires allongées[6]. En 2001, O'Keefe et ses collègues le récupèrent en tant que membre basal de cette famille, en dehors d'un groupe comprenant Liopleurodon, Pliosaurus et Brachauchenius[19] - [1]. Cependant, en 2008, les paléontologues Adam S. Smith et Gareth J. Dyke considèrent que Peloneustes serait le taxon frère de Pliosaurus[29] - [1]. En 2013, Benson et le paléontologue Patrick S. Druckenmiller nomment un nouveau clade au sein des Pliosauridae, appelé Thalassophonea. Ce clade comprend les pliosauridés « classiques » à col court tout en excluant les formes antérieures à long cou plus graciles, Peloneustes s'avérant être le membre le plus basal de ce groupe[32]. Des études ultérieures découvrent une position similaire pour Peloneustes[33] - [34] - [35] - [36].
Le cladogramme simplifié du clade Thalassophonea ci-dessous est basée d'après Madzia et al. (2018)[36] :
Paléobiologie
Mode de vie

Les plésiosaures furent bien adaptés à la vie marine[37] - [38] - [39]. Ils ont grandi à des taux comparables à ceux des oiseaux et avaient des métabolismes élevés, indiquant une possible homéothermie[40] et peut-être même de l'endothermie[39]. Le labyrinthe osseux, un creux dans le crâne qui contient un organe sensoriel associé à l'équilibre et à l'orientation, des Peloneustes et d'autres plésiosaures a une forme similaire à celle des tortues marines. Le paléontologue James Neenan et ses collègues émettent l'hypothèse en 2017 que cette forme a probablement évolué parallèlement aux mouvements de battement utilisés par les plésiosaures pour nager, les Peloneustes et autres plésiosaures à cou court avaient également des labyrinthes plus petits que les plésiosaures à cou plus long, un schéma également observé chez les cétacés[37]. De plus, Peloneustes avait probablement des glandes à sel dans sa tête pour faire face à une quantité excessive de sel dans son corps. Cependant, Peloneustes semble avoir été un prédateur des vertébrés, qui contiennent moins de sel que les invertébrés, ce qui conduit le paléontologue Leslie Noè à suggérer dans une thèse de 2001 que ces glandes n'auraient pas dû être particulièrement grandes[41]:257. Peloneustes, comme beaucoup d'autres pliosaures, affiche un niveau réduit d'ossification de ses os. Le paléontologue Arthur Cruickshank et ses collègues proposent en 1966 que cela aurait pu aider Peloneustes à maintenir sa flottabilité ou à améliorer sa maniabilité[42]. Une étude réalisée en 2019 par la paléontologue Corinna Fleischle et ses collègues révélent que les plésiosaures avaient des globules rouges agrandis, en fonction de la morphologie de leurs canaux vasculaires, ce qui les aurait aidés lors de la plongée[38].
Les plésiosaures tels que Peloneustes utilisaient une méthode de nage connue sous le nom de « vol sous-marin », utilisant leurs nageoires dans une manière semblables à ceux des hydroptères. Les plésiosaures sont inhabituels parmi les reptiles marins en ce sens qu'ils utilisaient leurs quatre membres mais pas les mouvements de la colonne vertébrale pour la propulsion. La queue courte, bien qu'il soit peu probable qu'elle ait été utilisée pour nager, aurait pu aider à stabiliser ou à diriger le plésiosaure[43] - [25]. Les nageoires avants de Peloneustes ont des rapports d'aspect de 6,36, tandis que les nageoires arrières ont des rapports d'aspect de 8,32. Ces rapports sont similaires à ceux des ailes des faucons actuels. En 2001, O'Keefe propose que, tout comme les faucons, les plésiosaures de type « pliosauromorphes » tels que Peloneustes étaient probablement capables de se déplacer rapidement et avec agilité, quoique de manière inefficace, afin de capturer des proies[23]. La modélisation informatique par la paléontologue Susana Gutarra et ses collègues en 2022 révèle qu'en raison de leurs grandes nageoires, un plésiosaure aurait produit plus de traînée qu'un cétacé ou un ichtyosaure de taille comparable. Cependant, les plésiosaures ont contrecarré cela avec leurs grands troncs et leur taille corporelle[44]. En raison de la réduction de la traînée par leurs corps plus courts et plus profonds, la paléontologue Judith Massare propose en 1988 que les plésiosaures pouvaient rechercher et poursuivre activement leur nourriture au lieu de devoir l'attendre[43].
Alimentation

Dans une thèse publié en 2001, paléontologue colombien Leslie Francis Noè note de nombreuses adaptations dans les crânes de pliosauridés pour la prédation. Pour éviter les dommages lors de l'alimentation, les crânes des pliosauridés comme Peloneustes sont hautement akinétiques, où les os du crâne et de la mandibule sont en grande partie verrouillés en place pour empêcher tout mouvement. Le museau contient des os allongés qui aident à empêcher la flexion et porte une jonction renforcée avec la région faciale pour mieux résister aux contraintes de l'alimentation. Vu de côté, peu de rétrécissement est visible dans la mandibule, ce qui la renforce. La symphyse mandibulaire aurait aidé à donner une morsure uniforme et à empêcher la mandibule de bouger indépendamment. Les processus coronoïdes élargie fournit une grande et forte région pour l'ancrage des muscles de la mâchoire, bien que cette structure ne soit pas aussi grande chez Peloneustes que chez d'autres pliosauridés contemporains. Les régions où les muscles de la mâchoire furent ancrés sont situées plus en arrière sur le crâne pour éviter toute interférence avec l'alimentation. La glène mandibulaire en forme de rein aurait rendu l'articulation de la mâchoire plus stable et empêché la mandibule de se disloquer. Les dents des pliosauridés sont fermement enracinées et imbriquées, ce qui renforce les bords des mâchoires. Cette configuration fonctionne également bien avec les mouvements de rotation simples auxquels les mâchoires des pliosauridés étaient limitées et renforce les dents contre les luttes des proies. Les plus grandes dents située à l'avant de la mâchoire auraient été utilisées pour empaler les proies tandis que les plus petites dents en arrière écrasaient et guidaient la proie vers la gorge. Avec leurs larges ouvertures, les pliosauridés n'auraient pas beaucoup transformé leur nourriture avant de l'avaler[41]:193, 236-240.

Les nombreuses dents de Peloneustes sont rarement cassées, mais montrent souvent des signes d'usure à leurs pointes. Leurs pointes acérées, légèrement incurvées, leur forme gracile et leur espacement proéminent indiquent qu'ils sont construits pour le perçage. Le museau mince et allongé ressemble à celui d'un dauphin. Les morphologies du museau et des dents conduit Noè à suggérer que Peloneustes était un piscivore. Pour attraper sa proie, Peloneustes aurait rapidement balancé la tête sur le côté. La section transversale à peu près circulaire du museau gracile aurait minimisé la traînée, tandis que les longues mâchoires étaient adaptées pour attraper rapidement des proies mobiles. Le palais plat et élargi et le neurocrâne renforcé de Peloneustes auraient réduit la torsion, la flexion et le cisaillement causés par le long museau. Le neurocrâne renforcé aurait réduit l'absorption des chocs dans la zone du cerveau. Comme la pointe du museau est plus éloignée de l'articulation de la mâchoire, elle aurait exercé une force de morsure plus faible que les régions plus en arrière. Les régions avant des mâchoires de Peloneustes sont allongées, ce qui indique qu'elles auraient été utilisées pour frapper et appréhender rapidement des proies. Ces adaptations indiquent une préférence pour les proies plus petites qui, bien qu'agiles, auraient été moins résistants et plus faciles à neutraliser. Cependant, Peloneustes aurait encore été capable d'attaquer des poissons de taille modérée. Alors qu'un squelette du spécimen NHMUK R3317 ayant des restes de bélemnite dans son estomac fut attribué à Peloneustes par Andrews en 1910[5]:XVI–XVII, il s'avère être très incomplet et pourrait en fait appartenir au pliosauridé contemporain Simolestes, comme le suggère Noè[41]:233-234, 241-242. Il est également suggéré en 2018 qu'un Peloneustes aurait infligé des marques de morsure à un spécimen de Cryptoclidus[45].
Paléoécologie
Environnement

Peloneustes est connu du membre de Peterborough, situé dans la formation d'Oxford Clay en Angleterre[1]. Alors que Peloneustes fut répertorié comme venant de l'étage Oxfordien (s'étalant d'environ 164 à 157 millions d'années[46]) du Jurassique supérieur[6], le membre de Peterborough date en fait du stade Callovien (situé environ vers 166 à 164 millions d'années[46]) du Jurassique moyen[1]. Le membre de Peterborough s'étend du Callovien inférieur tardif au Callovien supérieur précoce, occupant la totalité du Callovien moyen[47]. Il recouvre la formation de Kellaways (en)[47] et est recouvert par le membre de Stewartby de la formation d'Oxford Clay[48]. Le membre de Peterborough est principalement composée de schistes bitumeux[47] et d'argiles[49] riche en matière organique[50]. Ces roches s'avèrent être parfois fissiles (en) (sécables en dalles fines et plates)[48]. Le membre mesure environ 16 à 25 m d'épaisseur et s'étend de Dorset à Humber[47].
Le membre de Peterborough représente une mer intérieure à une époque d'élévation du niveau de la mer[50]. Lors de son dépôt, il aurait été situé à une latitude de 35°N[48]. Cette mer, connue sous le nom de mer d'Oxford Clay, était en grande partie encerclée par des îles et des continents, qui alimentaient la voie maritime en sédiments[48]. Sa proximité avec la terre est démontrée par la préservation de fossiles terrestres tels que le bois flotté d'Oxford Clay, en plus d'une digue clastique (en) dans les niveaux inférieurs du membre de Peterborough, la formation de la digue étant facilitée par l'eau de pluie[49]. La région sud de la mer d'Oxford Clay était reliée à l'océan Téthys, tandis qu'elle était reliée à des régions plus boréales sur son côté nord. Cela a permis des échanges fauniques entre les régions boréales et de la Téthys. Cette mer était d'environ 30 à 50 m de profondeur à moins de 150 km du rivage[48] - [1].
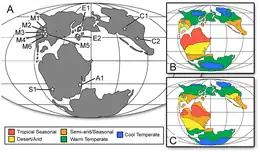
La terre environnante aurait eu un climat méditerranéen, avec des étés secs et des hivers humides, même si elle devenait de plus en plus aride. Sur la base des informations provenant des isotopes δ18O chez les bivalves, la température de l'eau du fond marin du membre de Peterborough variait de 14 à 17 °C en raison des variations saisonnières, avec une température moyenne de 15 °C. Les fossiles de bélemnite fournissent des résultats similaires, donnant une plage de température de l'eau avec un minimum de 11 °C à un maximum situé entre 14 ou 16 °C, avec une température moyenne de 13 °C[48]. Alors que des traces de bactéries soufrées vertes indiquent une eau euxinique, avec de faibles niveaux d'oxygène et de sulfure d'hydrogène élevé, des traces abondantes d'organismes benthiques suggèrent que les eaux du fond n'étaient pas anoxiques[51] - [50]. Les niveaux d'oxygène semblent avoir varié, certains dépôts étant déposés dans des conditions plus aérées que d'autres[48].
Biote contemporain
De nombreux types d'invertébrés sont préservés dans le membre de Peterborough. Parmi ceux-ci figurent les céphalopodes, qui comprennent les ammonites, les bélemnites et les nautiloïdes. Les bivalves sont un autre groupe abondant, tandis que les gastéropodes et les annélides le sont moins mais restent assez communs. Des arthropodes sont également présents. Les brachiopodes et les échinodermes sont rares. Bien qu'ils ne soient pas connus à partir de fossiles, les polychètes auraient probablement été présents dans cet écosystème, en raison de leur abondance dans des environnements modernes similaires et des terriers similaires à ceux produits par ces vers. Les microfossiles appartenant aux foraminifères, aux coccolithophoroïdes et aux dinoflagellés sont abondants dans le membre de Peterborough[52].
Une grande variété de poissons est connue du membre de Peterborough. Ceux-ci incluent les chondrichtyens tels que Heterodontus, Hybodus ou Notidanus. Des actinoptérygiens sont également présents, représentés par Aspidorhynchus, Leedsichthys, Lepidotes et Pholidophorus[53]. Ces poissons comprennent des variétés de surface, pélagiques et benthiques de différentes tailles, dont certaines sont assez grandes. Ils remplissent une variété de niches, y compris des mangeurs d'invertébrés, des piscivores et, dans le cas de Leedsichthys, des grands filtreurs[52].

Les plésiosaures sont communs dans le membre de Peterborough et, outre les pliosauridés, sont représentés par des Cryptoclididae, notamment Cryptoclidus, Muraenosaurus, Tricleidus et Picrocleidus[5]:VIII - [1]. C'étaient des plésiosaures plus petits avec des dents fines et un long cou et, contrairement aux pliosauridés tels que Peloneustes, ils auraient principalement mangé de petits animaux[52]. L'ichtyosaure Ophthalmosaurus habitait également la formation d'Oxford Clay. Ophthalmosaurus était bien adapté à la plongée profonde, grâce à son corps fuselé ressemblant à un marsouin et à ses imposants yeux, et se nourrissait probablement de céphalopodes[5]:XIV - [52]. De nombreux genres de crocodylomorphes sont également connus du membre de Peterborough. Ceux-ci incluent des téléosauroïdes[54] et les métriorhynchidés Gracilineustes, Suchodus, Thalattosuchus[9]:180 - [55] et Tyrannoneustes[56]. Bien que peu commun, le petit ptérosaure piscivore Rhamphorhynchus faisait également partie de cet écosystème marin[52].

Plus d'espèces de pliosauridés sont connues du membre de Peterborough que tout autre assemblage[1]. Outre Peloneustes, ces pliosauridés comprennent Liopleurodon, Simolestes, "Pliosaurus" andrewsi, Marmornectes[57], Eardasaurus et potentiellement Pachycostasaurus[58] - [59]. Cependant, l'anatomie de ces taxons varie considérablement, ce qui indique qu'elles se nourrissaient de proies différentes, évitant ainsi la concurrence[41]:249-251 - [42]. Le grand et puissant pliosauridé Liopleurodon semble avoir été adapté pour s'attaquer à de grandes proies, y compris d'autres reptiles marins et de gros poissons[41]:242-243, 249-251. Eardasaurus, de morphologie similaire peut également avoir pris de grandes proies[59]. Simolestes, avec son crâne large et profond et sa puissante morsure, semble avoir été un prédateur se nourrissant de grands céphalopodes[41]:243-244, 249-251. "Pliosaurus" andrewsi, comme Peloneustes, possède un museau allongé, une adaptation pour se nourrir de petits animaux agiles[52]. Cependant, ses dents sont adaptées à la coupe, indiquant une préférence pour les proies plus grosses, tandis que celles de Peloneustes sont mieux adaptées au perçage[60]. "Pliosaurus" andrewsi est également plus grand que Peloneustes[52]. Marmornectes est également similaire à Peloneustes, portant un long museau, et s'est peut-être aussi nourri de poisson[32] - [57]. Pachycostasaurus est un petit pliosaure fortement bâti qui se nourrissait probablement de proies benthiques. Ayant un crâne plus faible que les autres pliosauridés et étant plus stable, il a donc probablement utilisé différentes méthodes d'alimentation pour éviter la compétition. Contrairement aux autres pliosauridés d'Oxford Clay, Pachycostasaurus était plutôt rare, vivant peut-être principalement en dehors de la zone de dépôt de la formation, habitant peut-être des régions côtières, des eaux profondes ou même des rivières à la place[42]. Alors que plusieurs types différents de pliosauridés étaient présents au Jurassique moyen, les formes piscivores à long museau telles que Peloneustes se sont éteintes à la limite du Jurassique moyen-supérieur. Cela semble avoir été la première phase d'un déclin progressif de la diversité des plésiosaures. Bien que la cause en soit incertaine, elle peut avoir été influencée par l'évolution de la chimie des océans et, dans les phases ultérieures, par la baisse du niveau de la mer[32].
Notes et références
Notes
Références
- (en) H. F. Ketchum et R. B. J. Benson, « The cranial anatomy and taxonomy of Peloneustes philarchus (Sauropterygia, Pliosauridae) from the Peterborough member (Callovian, Middle Jurassic) of the United Kingdom », Palaeontology, vol. 54, no 3, , p. 639-665 (DOI 10.1111/j.1475-4983.2011.01050.x
 , S2CID 85851352, lire en ligne [PDF])
, S2CID 85851352, lire en ligne [PDF]) - (en) H. G. Seeley, Index to the fossil remains of Aves, Ornithosauria, and Reptilia, from the secondary system of strata arranged in the Woodwardian Museum of the University of Cambridge, , 139-140 p. (lire en ligne)
- (en) B. Creisler, « Ben Creisler's Plesiosaur Pronunciation Guide », (consulté le )
- (en) R. Araújo, A. S. Smith et J. Liston, « The Alfred Leeds Collection of the National Museum of Ireland—Natural History », Irish Journal of Earth Sciences, vol. 26, , p. 17-32 (DOI 10.3318/IJES.2008.26.17, JSTOR 20788277, S2CID 131513080, lire en ligne [PDF])
- C. W. Andrews, A descriptive catalogue of the marine reptiles of the Oxford clay. Based on the Leeds Collection in the British Museum (Natural History), London., vol. 1, Londres, British Museum, (lire en ligne).
- (en) L. B. Tarlo, « A review of the Upper Jurassic pliosaurs », Bulletin of the British Museum (Natural History), vol. 4, no 5, , p. 145-189 (lire en ligne)
- (en) R. Lydekker, « On the remains and affinities of five genera of Mesozoic reptiles », The Quarterly Journal of the Geological Society of London, vol. 45, nos 1-4, , p. 41–59 (DOI 10.1144/GSL.JGS.1889.045.01-04.04, S2CID 128586645, lire en ligne)
- (en) H. G. Seeley, « The nature of the shoulder girdle and clavicular arch in Sauropterygia », Proceedings of the Royal Society of London, vol. 51, , p. 119-151 (DOI 10.1098/rspl.1892.0017 , Bibcode 1892RSPS...51..119S, S2CID 128671277, lire en ligne)
- (en) C. W. Andrews, A descriptive catalogue of the marine reptiles of the Oxford clay. Based on the Leeds Collection in the British Museum (Natural History), London., vol. 2, Londres, British Museum, (lire en ligne)
- (en) C. W. Andrews, « On the structure of the skull of Peloneustes philarchus, a pliosaur from the Oxford Clay », Annals and Magazine of Natural History; Zoology, Botany, and Geology, vol. 16, no 93, , p. 242-256 (lire en ligne)
- F. Jaccard, « Notes sur le Peloneustes philarchus Seeley du musée paléontologique de Lausanne », Bulletin de la Société Vaudoise des Sciences Naturelles, vol. 43, no 160, , p. 395-398 (lire en ligne)
- (de) H. Linder, « Beiträge zur Kenntnis der Plesiosaurier-Gattungen Peloneustes und Pliosaurus », Geologische und Palaeontologische Abhandlungen, vol. 11, , p. 339-409 (lire en ligne)
- (en) C. W. Andrews, « Note on a mounted skeleton of a small pliosaur, Peloneustes philarchus Seeley », Geological Magazine, vol. 7, no 3, , p. 110-112 (DOI 10.1017/S0016756800132960, S2CID 130045734, lire en ligne)
- (en) J. Phillips, Geology of Oxford and the valley of the Thames, Oxford: Clarendon Press, (lire en ligne)
- (en) R. Lydekker, Catalogue of the fossil Reptilia and Amphibia in the British Museum (Natural History). Part IV. Containing the orders Anomodontia, Ecaudata, Caudata, Labyrinthodonta; and supplement, Londres, Trustees of the British Museum, (lire en ligne), p. 273
- (en) E. M. Knutsen, « A taxonomic revision of the genus Pliosaurus (Owen, 1841a) Owen, 1841b », Norwegian Journal of Geology, vol. 92, , p. 259-276 (lire en ligne [PDF])
- (ru) N. Novozhilov, « Два новых плиозавра из нижнего волжского яруса Поволжья », Doklady Akademii Nauk SSSR, vol. 60, , p. 115-118 (lire en ligne [PDF])
- (en) G. W. Storrs, M. S. Arkhangel'skii et V. M. Efimov, « Mesozoic marine reptiles of Russia and other former Soviet republics », dans M. J. Benton, M. A. Shishkin, D. M. Unwin et E. N. Kurochkin, The Age of Dinosaurs in Russia and Mongolia, Cambridge, Cambridge University Press, , 696 p. (ISBN 978-0-521-55476-3, lire en ligne [PDF]), p. 187-209
- (en) F. R. O’Keefe, « A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia) », ActaZoologica Fennica, vol. 213, , p. 1-63 (S2CID 82936031, lire en ligne)
- (en) Z. Gasparini et M. A. Iturralde-Vinent, « The Cuban Oxfordian herpetofauna in the Caribbean Seaway », Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen, vol. 240, no 3, , p. 343-371 (DOI 10.1127/njgpa/240/2006/343, S2CID 55346096, lire en ligne [archive du ] [PDF])
- (en) Z. Gasparini, « A new Oxfordian pliosaurid (Plesiosauria, Pliosauridae) in the Caribbean seaway », Palaeontology, vol. 52, no 3, , p. 661-669 (DOI 10.1111/j.1475-4983.2009.00871.x , S2CID 55353949, lire en ligne [PDF])
- (en) C. R. McHenry, « Devourer of Gods: The palaeoecology of the Cretaceous pliosaur Kronosaurus queenslandicus », The University of Newcastle, (hdl 1959.13/935911 , S2CID 132852950)
- (en) F. R. O'Keefe, « Ecomorphology of plesiosaur flipper geometry », Journal of Evolutionary Biology, vol. 14, no 6, , p. 987-991 (DOI 10.1046/j.1420-9101.2001.00347.x , S2CID 53642687)
- (en) J. Sassoon, D. Foffa et R. Marek, « Dental ontogeny and replacement in Pliosauridae », Royal Society Open Science, vol. 2, no 11, , p. 150384 (PMID 26715998, PMCID 4680613, DOI 10.1098/rsos.150384 , Bibcode 2015RSOS....250384S)
- (en) A. S. Smith, « Morphology of the caudal vertebrae in Rhomaleosaurus zetlandicus and a review of the evidence for a tail fin in Plesiosauria », Paludicola, vol. 9, no 3, , p. 144-158 (lire en ligne [PDF])
- (en) « Note on some of the generic modifications of the plesiosaurian pectoral arch », Quarterly Journal of the Geological Society, vol. 30, nos 1-4, , p. 436–449 (DOI 10.1144/gsl.jgs.1874.030.01-04.48, S2CID 128746688, lire en ligne)
- (en) T. E. White, « Holotype of Plesiosaurus longirostris Blake and classification of the plesiosaurs », Journal of Paleontology, vol. 14, no 5, , p. 451-467
- (en) P. O. Perssons, « A revision of the classification of the Plesiosauria with a synopsis of the stratigraphical and geographical distribution of the group », Lunds Universitets Arsskrift, vol. 59, no 1, , p. 1-59 (lire en ligne [PDF])
- (en) A. S. Smith et G. J. Dyke, « The skull of the giant predatory pliosaur Rhomaleosaurus cramptoni: Implications for plesiosaur phylogenetics », Naturwissenschaften, vol. 95, no 10, , p. 975-980 (PMID 18523747, DOI 10.1007/s00114-008-0402-z, Bibcode 2008NW.....95..975S, S2CID 12528732, lire en ligne)
- (en) K. Carpenter, « Comparative cranial anatomy of two North American Cretaceous plesiosaurs », dans I. M. Callaway et E. L. Nicholls, Ancient Marine Reptiles, San Diego, Academic Press, , 501 p. (ISBN 978-0-12-155210-7, DOI 10.1016/B978-012155210-7/50011-9, S2CID 88587032, lire en ligne [PDF]), p. 191–216
- (en) R. B. J. Benson, M. Evans et P. S. Druckenmiller, « High diversity, low disparity and small body size in Plesiosaurs (Reptilia, Sauropterygia) from the Triassic–Jurassic boundary », PLOS ONE, vol. 7, no 3, , e31838 (PMID 22438869, PMCID 3306369, DOI 10.1371/journal.pone.0031838 , Bibcode 2012PLoSO...731838B)
- (en) R. B. J. Benson et P. S. Druckenmiller, « Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition », Biological Reviews, vol. 89, no 1, , p. 1-23 (PMID 23581455, DOI 10.1111/brv.12038, S2CID 19710180)
- (en) R. B. J. Benson, M. Evans, A. S. Smith, J. Sassoon, S. Moore-Faye, H. F. Ketchum et R. Forrest, « A Giant Pliosaurid Skull from the Late Jurassic of England », PLOS ONE, vol. 8, no 5, , e65989 (PMID 23741520, PMCID 3669260, DOI 10.1371/journal.pone.0065989 , Bibcode 2013PLoSO...865989B)
- (en) Valentin Fischer, Maxim S. Arkhangelsky, Ilya M. Stenshin et Gleb N. Uspensky, « Peculiar macrophagous adaptations in a new Cretaceous pliosaurid », Royal Society Open Science, vol. 2, no 12, , p. 150552 (PMID 27019740, PMCID 4807462, DOI 10.1098/rsos.150552 , Bibcode 2015RSOS....250552F)
- (en) V. Fischer, R. B. J. Benson, N. G. Zverkov, L. C. Soul, M. S. Arkhangelsky, O. Lambert, I. M. Stenshin, G. N. Uspensky et P. S. Druckenmiller, « Plasticity and Convergence in the Evolution of Short-Necked Plesiosaurs », Current Biology, vol. 27, no 11, , p. 1667-1676 (PMID 28552354, DOI 10.1016/j.cub.2017.04.052 , S2CID 39217763)
- (en) D. Madzia, S. Sachs et J. Lindgren, « Morphological and phylogenetic aspects of the dentition of Megacephalosaurus eulerti, a pliosaurid from the Turonian of Kansas, USA, with remarks on the cranial anatomy of the taxon », Geological Magazine, vol. 156, no 7, , p. 1-16 (DOI 10.1017/S0016756818000523, Bibcode 2019GeoM..156.1201M, S2CID 133859507)
- (en) J. M. Neenan, T. Reich, S. W. Evers, P. S. Druckenmiller, D. F. A. E. Voeten, J. N. Choiniere, P. M. Barrett, S. E. Pierce et R. B. J. Benson, « Evolution of the sauropterygian labyrinth with increasingly pelagic lifestyles », Current Biology, vol. 27, no 24, , p. 3852-3858 (PMID 29225027, DOI 10.1016/j.cub.2017.10.069 , S2CID 207053689, lire en ligne)
- (en) C. V. Fleischle, P. M. Sander, T. Wintrich et K. R. Caspar, « Hematological convergence between Mesozoic marine reptiles (Sauropterygia) and extant aquatic amniotes elucidates diving adaptations in plesiosaurs », PeerJ, vol. 7, , e8022 (PMID 31763069, PMCID 6873879, DOI 10.7717/peerj.8022)
- (en) C. V. Fleischle, T. Wintrich et P. M. Sander, « Quantitative histological models suggest endothermy in plesiosaurs », PeerJ, vol. 6, , e4955 (PMID 29892509, PMCID 5994164, DOI 10.7717/peerj.4955 )
- (en) A. Houssaye, « Bone histology of aquatic reptiles: What does it tell us about secondary adaptation to an aquatic life? », Biological Journal of the Linnean Society, vol. 108, no 1, , p. 3-21 (DOI 10.1111/j.1095-8312.2012.02002.x , S2CID 82741198)
- (en) L. F. Noè, « A taxonomic and functional study of the Callovian (Middle Jurassic) Pliosauroidea (Reptilia, Sauropterygia) », University of Derby, Chicago,
- (en) A. R. I. Cruickshank, D. M. Martill et L. F. Noè, « A pliosaur (Reptilia, Sauropterygia) exhibiting pachyostosis from the Middle Jurassic of England », Journal of the Geological Society, vol. 153, no 6, , p. 873-879 (DOI 10.1144/gsjgs.153.6.0873, Bibcode 1996JGSoc.153..873C, S2CID 129602868, lire en ligne)
- J. A. Massare, « Swimming Capabilities of Mesozoic Marine Reptiles: Implications for Method of Predation », Paleobiology, vol. 14, no 2, , p. 187-205 (DOI 10.1017/S009483730001191X, S2CID 85810360)
- (en) S. Gutarra, T. L. Stubbs, B. C. Moon, C. Palmer et M. J. Benton, « Large size in aquatic tetrapods compensates for high drag caused by extreme body proportions », Communications Biology, vol. 5, , p. 380 (PMID 35484197, PMCID 9051157, DOI 10.1038/s42003-022-03322-y )
- (en) B. M. Rothschild, N. D. L. Clark et C. M. Clark, « Evidence for survival in a Middle Jurassic plesiosaur with a humeral pathology: What can we infer of plesiosaur behaviour? », Palaeontologia Electronica, vol. 21, no 16, , p. 1-11 (DOI 10.26879/719 , S2CID 133806762)
- (en) K. M. Cohen, S. Finney et P. L. Gibbard, « International Chronostratigraphic Chart » [PDF], International Commission on Stratigraphy,
- (en) K. L. Duff, « Palaeoecology of a bituminous shale—the Lower Oxford Clay of central England », Palaeontology, vol. 18, no 3, , p. 443-482 (résumé, lire en ligne [PDF])
- (en) C. Mettam, A. L. A. Johnson, E. V. Nunn et B. R. Schöne, « Stable isotope (δ18O and δ13C) sclerochronology of Callovian (Middle Jurassic) bivalves (Gryphaea (Bilobissa) dilobotes) and belemnites (Cylindroteuthis puzosiana) from the Peterborough Member of the Oxford Clay Formation (Cambridgeshire, England): evidence of palaeoclimate, water depth and belemnite behaviour », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 399, , p. 187–201 (DOI 10.1016/j.palaeo.2014.01.010, Bibcode 2014PPP...399..187M, hdl 10545/592777 , S2CID 129844404, lire en ligne [PDF])
- (en) J. D. Hudson, « The Peterborough Member (Callovian, Middle Jurassic) of the Oxford Clay Formation at Peterborough, UK », Journal of the Geological Society, vol. 151, no 1, , p. 113-124 (DOI 10.1144/gsjgs.151.1.0113, Bibcode 1994JGSoc.151..113H, S2CID 130058981, lire en ligne)
- (en) S. Belin et F. Kenig, « Petrographic analyses of organo-mineral relationships: depositional conditions of the Oxford Clay Formation (Jurassic), UK », Journal of the Geological Society, vol. 151, no 1, , p. 153-160 (DOI 10.1144/gsjgs.151.1.0153, Bibcode 1994JGSoc.151..153B, S2CID 131433536, CiteSeerx 10.1.1.1001.7308)
- (en) F. Kenig, J. D. Hudson, J. S. S. Damsté et B. N. Popp, « Intermittent euxinia: Reconciliation of a Jurassic black shale with its biofacies », Geology, vol. 32, no 5, , p. 421-424 (DOI 10.1130/G20356.1, Bibcode 2004Geo....32..421K, lire en ligne)
- (en) D. M. Martill, M. A. Taylor, K. L. Duff, J. B. Riding et P. R. Bown, « The trophic structure of the biota of the Peterborough Member, Oxford Clay Formation (Jurassic), UK », Journal of the Geological Society, vol. 151, no 1, , p. 173-194 (DOI 10.1144/gsjgs.151.1.0173, Bibcode 1994JGSoc.151..173M, S2CID 131200898, lire en ligne)
- (en) D. M. Martill, « Fish », dans D. M. Martill et J. D. Hudson, Fossils of the Oxford Clay, Londres, The Palaeontological Association, , 286 p., PDF (ISBN 0-901702-46-3, lire en ligne), p. 197-225
- (en) M. M. Johnson, M. T. Young et S. L. Brusatte, « The phylogenetics of Teleosauroidea (Crocodylomorpha, Thalattosuchia) and implications for their ecology and evolution », PeerJ, vol. 8, , e9808 (PMID 33083104, PMCID 7548081, DOI 10.7717/peerj.9808 )
- (en) M. T. Young, A. Brignon, S. Sachs, J. J. Hornung, D. Foffa, J. J. N. Kitson, M. M. Johnson et L. Steel, « Cutting the Gordian knot: A historical and taxonomic revision of the Jurassic crocodylomorph Metriorhynchus », Zoological Journal of the Linnean Society, vol. 192, no 2, , p. 510-553 (DOI 10.1093/zoolinnean/zlaa092 , S2CID 228853687)
- (en) S. Sachs, M. T. Young, P. Abel et H. Mallison, « A new species of the metriorhynchid crocodylomorph Cricosaurus from the Upper Jurassic of southern Germany », Acta Palaeontologica Polonica, vol. 64, no 2, , p. 343-356 (DOI 10.4202/app.00541.2018 , S2CID 133953687, lire en ligne [PDF])
- (en) H. F. Ketchum et R. B. J. Benson, « A new pliosaurid (Sauropterygia, Plesiosauria) from the Oxford Clay Formation (Middle Jurassic, Callovian) of England: Evidence for a gracile, longirostrine grade of Early-Middle Jurassic pliosaurids », Special Papers in Palaeontology, vol. 86, , p. 109-129 (S2CID 133227079)
- (en) L. F. Noè, J. Liston et M. Evans, « The first relatively complete exoccipital-opisthotic from the braincase of the Callovian pliosaur, Liopleurodon », Geological Magazine, vol. 140, no 4, , p. 479–486 (DOI 10.1017/S0016756803007829, Bibcode 2003GeoM..140..479N, S2CID 22915279, lire en ligne [archive du ])
- (en) H. F. Ketchum et R. B. J. Benson, « A new pliosaurid from the Oxford Clay Formation of Oxfordshire, UK », Acta Palaeontologica Polonica, vol. 67, no 2, , p. 297-315 (ISSN 0567-7920, DOI 10.4202/app.00887.2021 , S2CID 249034986)
- (en) J. A. Massare, « Tooth morphology and prey preference of Mesozoic marine reptiles », Journal of Vertebrate Paleontology, vol. 7, no 2, , p. 121-137 (DOI 10.1080/02724634.1987.10011647, S2CID 83647928)
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :