Cretoxyrhina
Requin Ginsu

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Chondrichthyes |
| Super-ordre | Selachimorpha |
| Ordre | Lamniformes |
Espèces de rang inférieur
- P. mantelli Zhelezko, 2000
- P. vraconensis Zhelezko, 2000
- T. agassizensis Underwood & Cumbaa, 2010
Cretoxyrhina (littéralement « nez pointu du Crétacé ») est un genre éteint de grands requins lamniformes ayant vécu durant le Crétacé (Albien-Campanien), il y a entre 107 et 73 millions d'années avant notre ère. L'espèce type, Cretoxyrhina mantelli, est communément désignée sous le nom vernaculaire de « requin Ginsu », en référence à la marque de couteau éponyme, car son mécanisme d'alimentation théorique est souvent comparé à la technique de « tranchage et découpage » mise en avant par la marque. Cretoxyrhina est traditionnellement classé comme le seul membre probable de la famille des Cretoxyrhinidae, bien que certains auteurs proposent de le classer au sein des Alopiidae ou des Lamnidae.
Mesurant jusqu'à 8 m de long pour un poids estimé à plus de 4,9 tonnes, Cretoxyrhina est l'un des plus grands requins de son temps. D'une morphologie similaire à celle du grand requin blanc actuel, il était un superprédateur de son écosystème et se nourrissait d'une grande variété d'animaux marins, dont des mosasaures, des plésiosaures, des requins et d'autres gros poissons, mais aussi des ptérosaures et parfois même des dinosaures. Ses dents, mesurant jusqu'à 8 cm de haut, ressemblent à des rasoirs et ont un épais émail spécialisé pour poignarder et trancher des proies. Cretoxyrhina figurait également parmi les requins les plus rapides : des modèles hydrodynamiques suggérant des capacités de vitesse de pointe allant jusqu'à 70 km/h. On suppose que Cretoxyrhina chassait en se jetant sur sa proie à une grande vitesse pour lui infliger des coups puissants en s'appuyant sur une vue efficace, une tactique de chasse similaire à celle du grand requin blanc.
Depuis la fin du XIXe siècle, plusieurs squelettes fossiles exceptionnellement bien conservés de Cretoxyrhina ont été découverts au Kansas. Des études ont réussi à calculer sa longévité en utilisant les vertèbres de certains spécimens. Cretoxyrhina aurait grandi rapidement durant les premières années pour atteindre la maturité sexuelle vers l'âge de quatre à cinq ans. Sa durée de vie estimée s'étend à près de quarante ans. L'analyse anatomique des squelettes de Cretoxyrhina révèle que le requin possède des caractéristiques faciales et optiques similaires à celles des requins-renards et des requins-crocodiles et une construction hydrodynamique qui suggère l'utilisation de l'endothermie régionale.
C'était un genre cosmopolite, ses fossiles ayant été trouvés dans le monde entier, bien qu'étant le plus souvent signalés dans la région de la voie maritime intérieure de l'Ouest, en Amérique du Nord. Il préférait les environnements pélagiques subtropicaux à tempérés, mais a également vécu dans des eaux froides ayant une température de 5 °C. Cretoxyrhina connait son apogée au Coniacien, puis décline continuellement jusqu'à son extinction au Campanien. Les causes suggérées de sa disparition sont la pression croissante de la concurrence avec l'apparition de nouveaux prédateurs comme Tylosaurus, un mosasauridé plus imposant que Cretoxyrhina et la disparition progressive de la voie maritime intérieure de l'Ouest.
Historique des recherches
Découvertes et fossiles
.jpg.webp)
Cretoxyrhina est décrit pour la première fois par le paléontologue anglais Gideon Mantell à partir de huit dents provenant de C. mantelli qu'il a recueillies dans le Southerham Grey Pit près de Lewes, dans le Sussex de l'Est, en Angleterre[11]. Dans son livre de 1822 intitulé The fossils of the South Downs, il les identifie comme des dents appartenant à deux espèces de requins modernes connues localement : les plus petites proviendraient de l'émissole lisse et les plus grandes du requin-marteau commun, avec une certaine hésitation à l'égard de ce dernier[12]. En 1843, le naturaliste suisse Louis Agassiz publie le troisième volume de son livre intitulé Recherches sur les poissons fossiles, où il réexamine les huit dents découvertes par Mantell. En les comparant avec une autre dent de la collection du musée zoologique de la ville de Strasbourg (dont la localisation exacte n'est pas précisée mais qui provient aussi d'Angleterre), il conclut que les fossiles appartiennent en fait à une seule espèce de requin disparue, qui présente de fortes similitudes dentaires avec les trois espèces alors classées dans le genre maintenant invalide Oxyrhina, à savoir O. hastalis, O. xiphodon et O. desorii[13] - [N 1]. Agassiz place l'espèce dans le genre Oxyrhina mais, notant que la racine plus épaisse de ses dents possède suffisamment de différences pour être distinguée, il la classe sous le nom scientifique Oxyrhina mantellii[N 2], qu'il nomme en l'honneur de Mantell[13].
À la fin du XIXe siècle, les paléontologues décrivent de nombreuses espèces qui sont maintenant considérées comme synonymes de C. mantelli. Selon certains chercheurs, il pourrait y avoir jusqu'à près de 30 synonymes différents d'O. mantelli définis à cette époque[17]. La plupart de ces espèces sont décrites à partir de dents qui représentent des variations de C. mantelli mais s'écartent des caractéristiques exactes des syntypes[7]. Par exemple, en 1870, le paléontologue français Henri Émile Sauvage identifie des dents provenant de France qui ressemblent beaucoup aux syntypes d'O. mantelli d'Angleterre. Les dents comprennent également des mini-cuspides latérales (petites cuspides émaillées qui apparaissent à la base de la couronne principale de la dent), qui ne sont pas présentes dans les syntypes, ce qui l'amène à décrire ces dents sous l'épithète spécifique Otodus oxyrinoides[18]. En 1873, le paléontologue américain Joseph Leidy identifie des dents du Kansas et du Mississippi et les décrit sous le nom d'espèce Oxyrhina extenta. Ces dents sont plus larges et plus robustes que les syntypes d'O. mantelli d'Angleterre[18] - [19].
Tout cela change avec les découvertes des squelettes du requin exceptionnellement bien conservés de la formation de Niobrara, situé dans l'ouest du Kansas. Charles H. Sternberg découvre le premier squelette en 1890, qu'il décrit dans un article de 1907[17] :
« La chose remarquable à propos de ce spécimen est que la colonne vertébrale, bien que cartilagineuse, était presque complète, et que le grand nombre de 250 dents étaient en place. Quand Chas. R. Eastman, de Harvard, a décrit ce spécimen, il s'est avéré si complet qu'il a détruit près de trente synonymes utilisés pour nommer l'animal, et dérivés de nombreuses dents trouvées autrefois[trad 1]. »
— Charles H. Sternberg, Some animals discovered in the fossil beds of Kansas, 1907.
_(20765045131).jpg.webp)
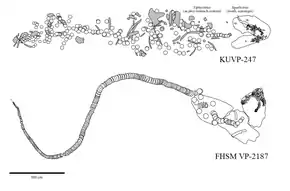
Charles R. Eastman publie son analyse du squelette en 1894 où il reconstruit la dentition sur la base de l'ensemble de dents désarticulées du squelette. À l'aide de la reconstitution, Eastman examine de nombreuses espèces éteintes de requins et découvre que leurs fossiles sont en fait différents types de dents d'O. mantelli, et les déplace tous au sein de l'espèce[7] - [18]. Le squelette, que Sternberg a vendu à l'université Louis-et-Maximilien de Munich, est détruit en 1944 par les bombardements alliés durant la Seconde Guerre mondiale[18] - [17]. En 1891, le fils de Sternberg, George F. Sternberg, découvre un deuxième squelette d'O. mantelli, hébergé au musée d'histoire naturelle de l'université du Kansas sous le nom de code KUVP 247. Ce squelette mesurerait 6,1 m de long et se compose d'une partie de colonne vertébrale avec des restes squelettiques d'un Xiphactinus comme contenu de l'estomac et une mâchoire partielle contenant environ 150 dents visibles. Ce squelette est considéré comme l'une des plus grandes découvertes scientifiques de cette année-là en raison de la préservation inattendue du cartilage[17]. G. F. Sternberg découvrira plus tard d'autres squelettes d'O. mantelli tout au long de sa carrière. Ses découvertes les plus notables sont les spécimens FHSM VP-323 et FHSM VP-2187, respectivement trouvés en 1950[20] et en 1965[21]. Le premier est un squelette partiel composé d'un ensemble de mâchoires bien conservé, d'une paire de cinq branchies et de quelques vertèbres tandis que le second est un squelette disposant d'une colonne vertébrale presque complète et d'un crâne exceptionnellement bien préservé contenant une grande partie des éléments de ce dernier, incluant des mâchoires, des dents, un ensemble d'écailles, des fragments de la ceinture pectorale et des nageoires dans leurs positions naturelles. Les deux squelettes sont actuellement conservés au musée d'histoire naturelle de Sternberg[22]. En 1968, un collectionneur nommé Tim Basgall découvre un autre squelette notable qui, à l'image de FHSM VP-2187, se compose également d'une colonne vertébrale presque complète et d'un crâne partiellement préservé. Ce fossile est conservé au musée d'histoire naturelle de l'université du Kansas sous le nom de code KUVP 69102[23].
En 1958, le paléontologue soviétique Leonid Glickman découvre que la conception dentaire d'O. mantelli reconstruite par Eastman le rend suffisamment distinct pour justifier un nouveau genre, qu'il nomme Cretoxyrhina[18] - [24]. Il identifie également une deuxième espèce de Cretoxyrhina, basée sur certaines dents plus anciennes connues du genre, qu'il nomme Cretoxyrhina denticulata[25] - [26]. À l'origine, Glickman désigne C. mantelli comme espèce type, mais il remplace brusquement la position par un autre taxon identifié comme « Isurus denticulatus », sans explication, dans un article de 1964, une décision maintenant rejetée comme taxonomiquent invalide[8]. Cela conduit néanmoins le paléontologue russe Viktor Zhelezko à invalider à tort le genre Cretoxyrhina dans un article de 2000 en synonymisant « Isurus denticulatus » (et donc le genre Cretoxyrhina dans son ensemble) avec un autre taxon identifié comme « Pseudoisurus tomosus »[N 3]. Zhelezko décrit également une nouvelle espèce congénère de C. mantelli basée sur du matériel dentaire provenant du Kazakhstan, qu'il identifie comme Pseudoisurus vraconensis conformément à sa réévaluation taxonomique[8] - [27]. Une étude de 2013 dirigée par le conservateur et paléontologue du Western Australian Museum Mikael Siverson corrige l'erreur taxonomique, rétablissant le genre Cretoxyrhina et déplaçant 'P'. vraconensis dans ce dernier[8]. En 2010, les paléontologues britanniques et canadiens Charlie Underwood et Stephen Cumbaa décrivent Telodontaspis agassizensis à partir de dents trouvées dans le lac Agassiz au Manitoba, qui avaient été précédemment identifiées comme des dents juvéniles de Cretoxyrhina[28]. Cette espèce est réintroduite dans le genre Cretoxyrhina dans une étude publiée en 2013 et menée par le paléontologue américain Michael G. Newbrey, en utilisant du matériel fossile supplémentaire du même taxon trouvé cette fois-ci en Australie-Occidentale[N 4] - [10].
Entre 1997 et 2008, le paléontologue Kenshu Shimada publie une série d'articles dans lesquels il analyse les squelettes de C. mantelli, y compris ceux trouvés par les Sternberg, en utilisant des techniques modernes pour effectuer des recherches approfondies sur la biologie possible de Cretoxyrhina. Certains de ses travaux incluent des reconstructions plus précises de sa dentition[29], de sa morphologie, physiologie[30], paléoécologie[31], ontogenèse[32] et de sa phylogénétique basées sur des variables morphologiques[33] sur Cretoxyrhina. Les recherches de Shimada sur Cretoxyrhina ont aidé à jeter un nouvel éclairage sur la compréhension du requin et, grâce à ses nouvelles méthodes, sur d'autres animaux disparus[10].
Étymologie du genre et des espèces
Cretoxyrhina est un mot-valise du mot creto, une abréviation du mot « Crétacé », préfixé au genre Oxyrhina, qui est dérivé du grec ancien ὀξύς / oxús « pointu » et ῥίς / rhís « nez ». Rassemblés, ces termes peuvent se traduire littéralement par « nez pointu du Crétacé », bien qu'il s'avère que Cretoxyrhina avait en fait un museau plutôt émoussé[30]. L'espèce mantelli fut dénommé par Agassiz en l'honneur de Gideon Mantell pour lui avoir fourni les syntypes de l'espèce[13]. L'épithète spécifique denticulata est dérivée du mot latin denticulus « petite dent » et du suffixe āta « possession de », signifiant ensemble « qui a de petites dents ». Il s'agit d'une référence à l'apparition de cuspides latérales dans la plupart des dents de C. denticulata[25]. L'espèce vraconensis est nommé en référence au Vraconien, sous-étage de l'Albien, dans lequel l'espèce a été découvert[27]. Le nom d'espèce agassizensis dérive du lac Agassiz, où l'espèce fut découverte[28]. Par coïncidence, le lac lui-même est nommé en l'honneur d'Agassiz[34], le naturaliste ayant nommé l'espèce type[13]. Le nom commun de « requin Ginsu », inventé à l'origine en 1999 par les paléontologues Mike Everhart et Kenshu Shimada, est une référence au couteau de la marque éponyme, car les mécanismes d'alimentation théoriques de C. mantelli furent souvent comparés au « tranchage et découpage », pour la manière dont le couteau est utilisé[35].
Description
Dentition
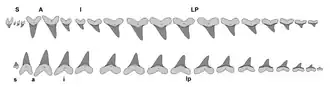
Les caractéristiques distinctives des dents de Cretoxyrhina comprennent une forme triangulaire presque symétrique ou inclinée, des tranchants en forme de rasoir et non dentelés, des cols de dents visibles (bourlette) et une couche d'émail épaisse. La dentition de Cretoxyrhina possède les caractéristiques dentaires de base d'un lamniforme, avec des rangées de dents étroitement espacées sans aucun chevauchement. Les dents antérieures sont droites et presque symétriques, tandis que les dents latéropostérieures sont inclinées. La face linguale (interne) est convexe et possède une protubérance massive et des rainures nutritives sur la racine, tandis que le côté labial (externe), est plat ou légèrement gonflé[29]. Les formes juvéniles possèdent des cuspides latérales sur toutes les dents[28], C. vraconensis les conservant systématiquement à l'âge adulte[8]. Les cuspides latérales ne sont conservées sur toutes les dents latéropostérieures à l'âge adulte chez C. denticulata et C. agassizensis et seulement jusqu'aux dents latéropostérieures arrière chez C. mantelli[28]. Les mini-cuspides latérales de C. vraconensis et C. denticulata sont rondes[8], tandis que celles de C. agassizensis sont aiguisés en pointe[11]. Les dents antérieures de C. vraconensis mesurent 2,1 à 3,5 cm de hauteur[8], tandis que la plus grande dent connue de C. denticulata mesure 3 cm[28]. Les dents de C. mantelli sont plus grandes, mesurant 3 à 4 cm de hauteur oblique moyenne. La plus grande dent découverte de cette espèce mesure 8 cm[10].


La dentition de C. mantelli est parmi la plus connus de tous les requins disparus, grâce à la préservation exceptionnelle de plusieurs squelettes fossiles comme le spécimen FHSM VP-2187, qui consiste en une dentition articulée et presque complète. D'autres squelettes de C. mantelli, tels que KUVP-247 et KUVP-69102, comprennent également des mâchoires partielles avec certaines dents dans leurs positions naturelles, dont certaines ne sont pas présentes dans des squelettes plus complets comme FHSM VP-2187. À l'aide de ces spécimens, la formule dentaire est reconstruite par Shimada en 1997 et révisée par ce dernier en 2002, où elle fut S4.A2.I4.LP11(+?) | S1?.A2.I1.LP15(+?). Cela signifie que, d'avant en arrière, C. mantelli avait : quatre symphysaires (petites dents situées dans la symphyse d'une mâchoire), deux antérieures, quatre intermédiaires et onze latéropostérieures ou plus pour la mâchoire supérieure et éventuellement une symphysaire, deux antérieures, un intermédiaire et quinze latéropostérieures ou plus pour la mâchoire inférieure. La structure de la rangée de dents montre une dentition adaptée à un comportement alimentaire similaire à celui des requins makos actuels qui possèdent de grandes dents antérieures en forme de lance pour poignarder et ancrer la proie et des latéropostérieurs incurvés pour la couper en morceaux de la taille d'une bouchée[29] - [36], un mécanisme souvent décrit de manière informelle en « tranchage et découpage » par les paléontologues[35]. En 2011, les paléontologues Jim Bourdon et Mike Everhart reconstruisent la dentition de plusieurs individus de C. mantelli en fonction de leurs ensembles de dents associés. Ils découvrent que deux de ces reconstitutions montrent des différences notables dans la taille de la première dent intermédiaire (I1) et les profils latéraux, concluant que ces différences pourraient éventuellement représenter un dimorphisme sexuel ou des variations individuelles[37].
Crâne
.jpg.webp)

L'analyse des motifs du crâne et des écailles montre que C. mantelli avait une tête conique avec un crâne plat et large dorsalement. Le rostre ne s'étend pas beaucoup vers l'avant à partir de la marge frontale du neurocrâne, ce qui suggère que le museau serait émoussé[30]. Semblable aux requins-crocodiles et aux requins-renards modernes, C. mantelli a des yeux proportionnellement grands, l'orbite occupant environ un tiers de la longueur totale du crâne, ce qui lui donnerait une vision aiguë. En tant que prédateur, une bonne vue est importante lors de la chasse aux grandes proies dont Cretoxyrhina se nourrissait. En revanche, les processus orbitaux moins avancés mais plus forts de l'anacoracidé contemporain Squalicorax, plus dépendants de l'odeur, conviendrait mieux à un charognard opportuniste[30] - [38].
Les mâchoires de C. mantelli auraient été cinétiquement puissantes. Elles ont une courbure antérieure légèrement plus lâche et une construction plus robuste que celle des requins makos actuels, bien que similaires dans leur forme générale. L'hyomandibule est allongée et se balancerait latéralement, ce qui permettrait une protrusion de la mâchoire et une morsure profonde[30].
Anatomie squelettique

La plupart des espèces de Cretoxyrhina ne sont représentées que par des dents et des vertèbres fossiles. À l'image des autres requins, le squelette de Cretoxyrhina est constitué de cartilage, qui se fossilise plus difficilement que l'os. Cependant, les fossiles de C. mantelli découverts dans la formation de Niobrara sont exceptionnellement bien préservés[39] grâce à la présence de craie, et sa teneur élevée en calcium, qui permet une calcification des tissus mous[40] et permet leur fossilisation[39].
De nombreux squelettes dont la colonnes vertébrales est presque complètes ont été trouvés. La plus grande vertèbre mesure 8,7 cm de diamètre. Les deux spécimens possédant la colonne vertébrale la mieux conservée (FHSM VP-2187 et KUVP 69102) ont respectivement 218 et 201 centra, et presque toutes les vertèbres de la colonne sont préservées, seules des parties de l'extrémité de la queue manquent. Les estimations de la longueur de la queue fournissent un nombre total de vertèbres d'environ 230 centra, ce qui est exceptionnel car tous les requins lamniformes existants possèdent un nombre de vertèbres inférieur à 197 ou supérieur à 282 sans aucun individus entre ces deux extrêmes. Les centra vertébraux de la région du tronc sont larges et circulaires, créant un corps global en forme de fuseau avec un tronc trapu[30].
Une analyse d'un fossile de nageoire caudale partiellement complet montre que Cretoxyrhina aurait une queue en forme de croissant très similaire à celles des lamnidés, des requins-baleines et des requins pèlerins actuels. Les chercheurs estiment que la transition vers les vertèbres caudales se situe entre la 140e et la 160e vertèbre sur un total de 230, ce qui donne un nombre total de vertèbres caudales de 70 à 90 et représente environ 30 à 39 % de la colonne vertébrale. La transition entre les vertèbres précaudales et les vertèbres caudales est également marquée par une courbure vertébrale d'environ 45°, qui est l'angle le plus élevé possible connu chez les requins existants et se trouve principalement chez les requins à nage rapide, tels que les lamnidés[41]. Les propriétés de la queue, ainsi que d'autres caractéristiques telles que des écailles lisses parallèles à l'axe du corps, une nageoire pectorale plésodique (une nageoire pectorale dans laquelle le cartilage s'étend partout, lui donnant une structure plus sûre qui aide à réduire la traînée), et un corps trapu en forme de fuseau, montrent que C. mantelli aurait été capable de nager rapidement[30] - [41].
Thermorégulation
Cretoxyrhina représente l'une des premières formes et l'origine possible de l'endothermie chez les lamniformes[42]. Grâce à une endothermie régionale (également connue sous le nom de mésothermie), il peut avoir possédé une construction similaire aux requins endothermiques régionaux modernes comme les membres des familles des requins-renards et des lamnidés[42], chez qui les muscles rouges sont plus proches de l'axe du corps que chez les requins ectothermes (dont les muscles rouges sont plus proches de la circonférence du corps), et où un système de vaisseaux sanguins spécialisés appelé rete mirabile est présent, permettant à la chaleur métabolique d'être conservée et échangée avec les organes vitaux. Cette construction morphologique permet au requin d'avoir partiellement le sang chaud[43], et de fonctionner ainsi efficacement dans les environnements plus froids où Cretoxyrhina fut trouvé. Des fossiles ont été trouvés dans des zones où les estimations paléoclimatiques montrent une température de surface aussi basse que 5 °C[42]. L'endothermie régionale de Cretoxyrhina peut avoir été développée en réponse à la pression croissante du refroidissement global progressif et à la concurrence des mosasaures et d'autres reptiles marins qui avaient également développé l'endothermie[42].
Hydrodynamique et locomotion
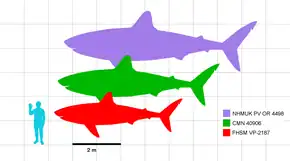
- Spécimen le plus imposant
- Spécimen nord-américain le plus imposant
- Spécimen le plus complet
Cretoxyrhina possède des écailles placoïdes superposées très denses, parallèles à l'axe du corps et disposées en structures parallèles appelées kneels, et séparées par des rainures en forme de « U ». Chaque rainure ayant une largeur moyenne d'environ 45 μm. Cette formation d'écailles permet une réduction efficace de la traînée et une plus habilité à nager à de grande vitesse, un morphotype observé uniquement chez les requins actuels les plus rapides[30] - [42].
La une nageoire caudale de « type 4 » de Cretoxyrhina possède également un angle de Cobb (courbure des vertèbres de la queue) et un angle du cartilage de la queue (respectivement entre 49° et 133°) les plus élevés jamais enregistrés chez les lamniformes[44] - [42]. Une structure de nageoire caudale de « type 4 » présente une symétrie maximale des lobes de la nageoire caudale, ce qui est le plus efficace en nage rapide, structure que l'on retrouve uniquement chez les lamnidés parmi les requins[44].
Une étude réalisée en 2017 par le doctorant Humberto Ferron analyse les relations entre les variables morphologiques, comprenant le squelette et la nageoire caudale de C. mantelli, et obtient une vitesse de croisière moyenne de 12 km/h et une vitesse de pointe d'environ 70 km/h, faisant probablement de Cretoxyrhina l'un des requins les plus rapides connus[42]. À titre de comparaison, le grand requin blanc moderne atteint des vitesses allant jusqu'à 56 km/h[45] et le requin mako, le requin existant le plus rapide connu, atteint 70 km/h[N 5] - [48].
Reproduction
Bien qu'aucune preuve fossile n'ait été trouvée, il est déduit que Cretoxyrhina aurait été ovovivipare comme tous les lamniformes actuels. Chez les requins ovovivipares, les jeunes éclosent et grandissent à l'intérieur de la mère tout en rivalisant avec les compagnons de portée par le cannibalisme tel que l'oophagie. Comme Cretoxyrhina habitait des eaux oligotrophes et pélagiques où les prédateurs tels que les grands mosasaures comme Tylosaurus et les grands poissons superprédateurs comme Xiphactinus étaient courants, il est probable qu'il s'agissait également d'un requin pratiquant la stratégie r, où de nombreux nourrissons sont produits pour compenser les taux élevés de mortalité infantile. De même, les requins pélagiques tels que le grand requin blanc, les requins-renards, les requins makos, le requin-taupe commun et le requin-crocodile produisent de deux à neuf petits par portée[32].
Croissance et longévité
.png.webp)
Comme chez tous les lamniformes, les vertèbres de Cretoxyrhina présentent des anneaux de croissance (un anneau apparait chaque année) et l’âge d’un individu peut être déterminé en les mesurant. En raison de la rareté des vertèbres bien conservées, seuls quelques individus connus de Cretoxyrhina ont vieilli. Dans l'étude de 1997 menée par Shimada, une équation linéaire pour calculer la longueur totale du corps de Cretoxyrhina en utilisant le diamètre du centrum d'une vertèbre a été développée et est illustrée ci-dessous, avec TL représentant la longueur totale du corps et CD le diamètre du centrum (le diamètre de chaque bande)[32] :
TL = 0.281 + 5.746CD
À l'aide de cette équation linéaire, des mesures ont d'abord été effectuées sur le spécimen de C. mantelli le mieux conservé, FHSM VP-2187, par Shimada en 2008. Les mesures montrent une longueur de 1,28 m et un poids d'environ 16,3 kg à la naissance, et une croissance rapide au cours des deux premières années de la vie, doublant la longueur en 3,3 ans. À partir de ce moment, la croissance de la taille devient régulière et progressive, augmentant en estimation moyenne de 21,1 cm/an jusqu'à sa mort vers l'âge de 15 ans, année à laquelle il passe à 5 cm. En utilisant le modèle de croissance de von Bertalanffy sur le spécimen FHSM VP-2187, la durée de vie maximale de C. mantelli est estimée à 38,2 ans. À cet âge, il aurait atteint plus de 7 m de long. Sur la base de la mise à l'échelle allométrique d'un grand requin blanc, Shimada découvre qu'un tel individu aurait pesé jusqu'à 3,4 tonnes[32].
Une nouvelle mesure effectuée par Newbrey et ses collègues en 2013 constate que C. mantelli et C. agassizensis auraient atteint la maturité sexuelle vers l'âge de quatre à cinq ans et propose une révision possible des mesures des anneaux de croissance chez FHSM VP-2187. Il a également été proposé de réviser la durée de vie de FHSM VP-2187 et la durée de vie maximale de C. mantelli entre 18 et 21 ans respectivement en utilisant les nouvelles mesures. Une étude menée en 2019 par le scientifique italien Jacopo Amalfitano sur les vertèbres de deux fossiles de C. mantelli, indique que l'individu le plus âgé serait mort vers l'âge de 26 ans[11]. Des mesures ont également été effectuées sur d'autres squelettes de C. mantelli et une vertèbre de C. agassizensis, donnant des résultats similaires de taux de croissance rapide aux premiers stades de vie[10]. Une croissance aussi rapide en quelques années seulement aurait pu aider Cretoxyrhina à mieux survivre en éliminant rapidement sa petitesse juvénile et ses vulnérabilités, car un individu adulte aurait peu, voire aucun prédateur naturel[32]. L'étude de 2013 identifie également une des dents syntypes de C. mantelli d'Angleterre et calcule la longueur maximale de l'individu à 8 m, faisant de la dent le plus grand spécimen identifié à ce jour[10] - [N 6]. Un C. mantelli d'une telle longueur donnerait une masse corporelle estimée à environ 4,9 tonnes en utilisant la méthode de Shimada[32] - [50].
Le graphique ci-dessous représente la croissance en longueur par an du spécimen FHSM VP-2187 selon Shimada (2008)[N 7] - [32] :
Les chercheurs estiment que les autres espèces auraient été beaucoup plus petites. C. denticulata et C. vraconensis auraient atteint une longueur corporelle totale allant jusqu'à 4 m à l'âge adulte[51] - [8] et un poids de 744 kg[50]. C. agassizensis est encore plus petit, avec une longueur corporelle totale estimée allant jusqu'à 1,29 m sur la base d'un spécimen de dent mesurant 12 mm de haut (catalogué P2989.152)[52] - [28].
Classification et évolution

Cretoxyrhina ressemble au grand requin blanc moderne en termes de taille, de morphologie et d'écologie, mais les deux requins ne sont pas liés d'un point de vue phylogénique, leurs similitudes étant le résultat d'une évolution convergente[32]. Cretoxyrhina a été traditionnellement inclut au sein des Cretoxyrhinidae, une famille de requins lamniformes qui comporte traditionnellement d'autres genres et formant ainsi une famille paraphylétique ou polyphylétique[53]. En 1999, Siverson fait remarquer que les Cretoxyrhinidae étaient utilisés comme taxon poubelle pour les requins du Crétacé et du Paléogène et déclare que Cretoxyrhina est le seul membre valide de cette famille[54].
| Relation phylogénétique possible entre Cretoxyrhina et les lamniformes actuels selon Shimada (2007)[55]. |
Cretoxyrhina contient quatre espèces reconnues : C. vraconensis, C. denticulata, C. agassizensis et C. mantelli. Ces espèces représentent chacune une chrono-espèce[11]. Les plus anciens fossiles de Cretoxyrhina datent d'environ 107 millions d'années (Albien) et appartiennent à C. vraconensis[3]. Ce dernier évoluera au cours du Cénomanien précoce en C. denticulata, lequel évoluera ensuite au cours du Cénomanien moyen en C. agassizensis, qui à son tour évoluera en C. mantelli au cours du Cénomanien tardif. Il est à noter que C. agassizensis a existé jusqu'au milieu du Turonien et fut brièvement contemporain de C. mantelli[11]. Cette évolution est caractérisée par la réduction des cuspides latérales et l'augmentation de la taille et de la robustesse des dents[2] - [56]. L'intervalle Albien tardif-Turonien moyen voit principalement la réduction des cuspides latérales ; C. vraconensis possède des cuspides latérales sur toutes les dents à l'exception de quelques-unes en position antérieure, qui se limiteraient progressivement aux latéropostérieures arrière chez les adultes à la fin de l'intervalle chez C. mantelli[2]. L'intervalle Cénomanien tardif-Coniacien est caractérisé par une augmentation rapide de la taille des dents et du corps, une diminution significative du rapport hauteur / largeur de la couronne[1] - [2], et une transition d'une forme de dent de type déchirant à une forme de dent de type coupant[51]. La taille des dents des individus de C. mantelli à l'intérieur de la voie maritime intérieure de l'Ouest culmine vers le Coniacien tardif, il y a environ 86 millions d'années, puis commence à décliner lentement[1]. En Europe, ce pic a lieu plus tôt, au cours du Turonien tardif[11]. Le plus récent fossile connu de C. mantelli provient de la formation de Bearpaw en Alberta, et date de 73,2 millions d'années (Campanien tardif) [5]. Une seule dent identifiée comme Cretoxyrhina sp. a été récupérée dans la formation voisine de Horseshoe Canyon et est datée de 70,44 millions d'années, ce qui suggère que Cretoxyrhina aurait peut-être survécu jusqu'au Maastrichtien. Cependant, la formation de Horseshoe Canyon s'est déposé dans un environnement saumâtre ce qui n'exclut pas l'éventualité que le fossile ait été remanié depuis une couche plus ancienne[6].
Des études phylogénétiques menées à travers des données morphologiques par Shimada en 2005 suggèrent que Cretoxyrhina pourrait alternativement être congénère avec Alopias, correspondant aux requins-renards actuels. L'étude déclare cependant que les résultats sont prématurés et peuvent être inexacts, et recommande que Cretoxyrhina soit conservé dans la famille des Cretoxyrhinidae, citant principalement le manque de données substantielles à son sujet lors de l'analyse[33].
Un autre modèle possible d'évolution de Cretoxyrhina, proposé en 2014 par le paléontologue Cajus Diedrich, suggère que C. mantelli aurait été congénère du genre Isurus et ferait partie d'une lignée étendue de ce dernier commençant jusqu'à l'Aptien. Selon ce modèle, la lignée Isurus débuta par 'Isurus appendiculatus' (Cretolamna appendiculata), qui a évolué en Isurus denticulatus (C. mantelli[11]) au Cénomanien moyen, puis 'Isurus mantelli' (C. mantelli) au début du Coniacien, Isurus schoutedenti au Paléocène, et Isurus praecursor, où se poursuit le reste de la lignée Isurus[N 8]. L'étude affirme que l'absence de fossiles du Maastrichtien ne serait pas le résultat d'une extinction prématurée de C. mantelli, mais simplement une lacune dans le registre fossile[51]. Shimada et son collègue paléontologue Phillip Sternes publient en 2018 un poster[9] qui met en doute la crédibilité de cette proposition, notant que l'interprétation de l'étude est largement basée sur des données qui avaient été arbitrairement sélectionnées et ne citait pas les études de Shimada publiées respectivement entre 1997[N 9] et 2005, qui sont des articles clés concernant la position systématique de C. mantelli.
Paléobiologie
Relations avec les proies

Les mâchoires cinétiquement puissantes, les capacités de vitesse élevées et la grande taille de Cretoxyrhina suggèrent un prédateur particulièrement agressif[30] - [31]. L'association de Cretoxyrhina avec un nombre diversifié de fossiles montrant des signes de dévoration confirme qu'il s'agissait d'un superprédateur actif qui se nourrissait d'une grande partie de la variété de la mégafaune marine du Crétacé tardif[31] - [57]. Le niveau trophique le plus élevé qu'il occupait était une position partagée uniquement avec les grands mosasaures tels que Tylosaurus au cours des dernières étapes du Crétacé supérieur. Il a joué un rôle essentiel dans de nombreux écosystèmes marins[57].

Cretoxyrhina se nourrissait principalement d'autres prédateurs actifs, notamment les ichtyodectidés (une famille de gros poissons qui inclut Xiphactinus)[31], des plésiosaures[31] - [58], des tortues[59], des mosasaures[31] - [60] - [35], et d'autres requins[31]. La plupart des fossiles de Cretoxyrhina se nourrissant d'autres animaux consistent en de grandes et profondes marques de morsures et de perforations sur les os, parfois avec des dents incrustées[35]. Les os isolés de mosasaures et d'autres reptiles marins qui ont été partiellement dissous par digestion, ou trouvés dans des coprolithes, sont également des exemples d'alimentation de Cretoxyrhina[35] - [60]. Il existe quelques squelettes de Cretoxyrhina contenant du contenu stomacal connu, y compris un grand squelette de C. mantelli (KUVP 247) qui possède des restes squelettiques de Xiphactinus et un mosasaure au niveau de l'estomac[31]. Cretoxyrhina peut s'être parfois nourri de ptérosaures, comme en témoigne un ensemble de vertèbres cervicales d'un Pteranodon de la formation de Niobrara avec une dent de C. mantelli logée profondément entre elles. Bien qu'il ne soit pas certain que le fossile lui-même soit le résultat d'une prédation ou de charognage, il est probable que Pteranodon et les ptérosaures similaires seraient des cibles naturelles pour des prédateurs comme Cretoxyrhina, car ils entreraient régulièrement dans l'eau pour se nourrir et seraient donc à portée de main[61] - [62].
Bien que Cretoxyrhinaait été principalement un chasseur actif, c'était aussi un opportuniste et il peut avoir eu un régime charognard occasionnellement. De nombreux fossiles avec des marques d'alimentation de Cretoxyrhina ne montrent aucun signe de guérison, un indicateur d'une attaque prédatrice délibérée sur un animal vivant, conduisant à la possibilité qu'au moins certaines des marques d'alimentation aient été faites à partir de charognards[31]. Des restes de squelettes partiels de dinosaures comme Claosaurus et Niobrarasaurus montrent des signes d'alimentation et de digestion par C. mantelli. Il s'agissait probablement de carcasses récupérées et balayées dans l'océan, l'emplacement paléogéographique des fossiles étant celui d'un océan ouvert[63] - [64].
Stratégies de chasse

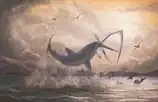
Les stratégies de chasse de Cretoxyrhina ne sont pas bien documentées car il n’est pas possible de distinguer si ses marques d’alimentation sur de nombreux fossiles résultent de la prédation et ou du charognage. Si elles étaient effectivement le résultat de la première option, cela signifierait que Cretoxyrhina aurait très probablement utilisé des stratégies de chasse impliquant un coup principal puissant et fatal similaire à l'alimentation observée chez les carcharhinidés et les lamnidés actuels. Une étude réalisée en 2004 observe qu'un grand requin blanc moderne atteignant des longueurs supérieures à 4 m fonce généralement sur sa proie avec une vitesse et une force massives pour infliger des coups mortels uniques, parfois si puissants que la proie est propulsée hors de l'eau par la force de l'impact. Comme Cretoxyrhina possède une construction robuste et trapue capable de nager rapidement, de puissantes mâchoires cinétiques semblables à celles du grand requin blanc et une taille similaire voire supérieure à celui-ci, un style de chasse comparable entre les deux squales serait donc probable[58].
Paléoécologie
Gamme et distribution

Cretoxyrhina avait une distribution cosmopolite, ses fossiles ayant été trouvés dans le monde entier. Les emplacements notables incluent l'Amérique du Nord, l'Europe[65], Israël[66] et le Kazakhstan[8]. Cretoxyrhina a vécu principalement dans les zones tempérées et subtropicales[2]. Il a été trouvé à des latitudes aussi élevées que 55 °N, où les estimations paléoclimatiques calculent une température moyenne de surface de la mer de 5 à 10 °C. Les fossiles de Cretoxyrhina sont surtout connus dans la région de la voie maritime intérieure de l'Ouest[66], une ancienne mer disparue qui a existé dans ce qui est maintenant le centre des États-Unis et le Canada[67]. En 2013, Siverson et ses collègues notent qu'au cours du Turonien ou du début du Coniacien, les individus de Cretoxyrhina vivant au large étaient généralement plus grands que ceux habitant la voie maritime intérieure de l'Ouest, certains fossiles de C. mantelli au large, comme l'un des syntypes, donnant des longueurs totales allant jusqu'à 8 m, voire peut-être même 9 m[8].
Habitat
Cretoxyrhina habitait principalement les océans pélagiques tempérés à subtropicaux. Une dent de Cretoxyrhina trouvée dans la formation de Horseshoe Canyon en Alberta (une formation déposée dans un environnement de littoral dominé par des eaux saumâtres) suggère qu'il a pu, à l'occasion, nager dans des estuaires partiellement d'eaux douces et des plans d'eaux similaires. Cependant, un remaniement à partir d'une couche sous-jacente peut être une explication plus probable d'une telle occurrence[68]. Le climat des écosystèmes marins pendant la plage temporelle de Cretoxyrhina est généralement beaucoup plus chaud qu’à l'époque moderne en raison des niveaux atmosphériques plus élevés de dioxyde de carbone et d'autres gaz à effet de serre influencés par la forme des continents de l'époque[69].
L'intervalle entre 97 et 91 millions d'années (Cénomanien à Turonien) a connu un pic des températures de la surface de la mer en moyenne supérieure à 35 °C et des températures de l'eau de fond d'environ 20 °C, environ 7 à 8 °C plus élevées qu'actuellement[69]. Durant cette période, Cretoxyrhina a coexisté avec une faune comme les mosasaures marquée par une forte diversification[70]. Cet intervalle comprend également une augmentation des niveaux globaux de δ13C, qui marque un appauvrissement significatif de l'oxygène dans l'océan, et a provoqué l'événement anoxique du Cénomanien-Turonien[69]. Bien que cet événement ait entraîné l'extinction de 27 % des invertébrés marins[71], les vertébrés comme Cretoxyrhina ne furent pas affectés[72]. Le reste du Crétacé a connu ensuite un refroidissement global et progressif des océans, conduisant à l'apparition d'écosystèmes tempérés et à une possible glaciation au Maastrichtien précoce[69]. Les zones subtropicales conservent une biodiversité élevée de tous les taxons, tandis que les écosystèmes tempérés avaient généralement une diversité beaucoup plus faible. En Amérique du Nord, les provinces subtropicales étaient dominées par les requins, les tortues et les mosasaures tels que Tylosaurus et Clidastes, tandis que les provinces tempérées étaient principalement dominées par les plésiosaures, les hesperornithidés et le mosasaure Platecarpus[73].
Compétition
Cretoxyrhina vivait aux côtés de nombreux prédateurs qui partageaient un niveau trophique similaire dans un écosystème pélagique diversifié au cours du Crétacé[57]. La plupart de ces prédateurs sont de grands reptiles marins, dont certains occupaient déjà le niveau trophique le plus élevé lorsque Cretoxyrhina est apparu pour la première fois[87]. De l'Albien jusqu'au Turonien (vers environ 107 à 91 millions d'années), Cretoxyrhina a coexisté avec les pliosaures du Crétacé précoce. Parmi certains de ces pliosaures figure l'imposant Megacephalosaurus dont la taille est estimée entre 6 et 9 m de long[88], mais les pliosaures s'éteignent au milieu du Turonien, vers environ 91 millions d'années[72] - [89]. La radiation des requins comme Cretoxyrhina pourrait avoir été un facteur majeur contribuant à l'accélération de leur extinction[90]. Le vide écologique qu'ils ont laissé a été rapidement comblé par les mosasaures émergents, qui sont également venus occuper les niveaux trophiques les plus élevés[91]. De grands mosasaures comme Tylosaurus, pouvant atteindre plus de 14 m de long[37], peuvent avoir rivalisé avec Cretoxyrhina, et des preuves d'interactions interspécifiques telles que des marques de morsure provenant de l'un ou de l'autre ont été trouvées[31]. Il y a aussi de nombreux requins qui occupaient un rôle écologique similaire à celui de Cretoxyrhina, comme les cardabiodontidés Cardabiodon[10] et Dwardius, ce dernier montrant des preuves de concurrence directe avec C. vraconensis sur la base de schémas de distribution complexes entre les deux[82].
Une étude réalisée en 2010 par les paléontologues Corinne Myers et Bruce Lieberman sur la concurrence dans la voie maritime intérieure de l'Ouest utilise des techniques analytiques quantitatives basées sur des systèmes d'information géographique et des reconstructions tectoniques pour reconstruire les relations de concurrence hypothétiques entre dix des vertébrés marins les plus répandus et les plus abondants de la région, incluant Cretoxyrhina. Leurs calculs trouvent des corrélations négatives entre la distribution de Cretoxyrhina et les trois concurrents potentiels Squalicorax kaupi, Tylosaurus proriger et Platecarpus spp. ; une corrélation négative statistiquement significative implique que la distribution d'une espèce a été affectée parce qu'elle a été supplantée par une autre. Cependant, aucune des relations n'est statistiquement significative, ce qui indique plutôt que les tendances sont peu susceptibles d'être le résultat de la concurrence[92].
Extinction

Les causes d'extinction de Cretoxyrhina sont incertaines. Ce que l'on sait, c'est que le taxon décline lentement sur des millions d'années[2]. Depuis son apogée au Coniacien, la taille[1] et la répartition[2] des fossiles de Cretoxyrhina diminue progressivement jusqu'à sa disparition éventuelle au cours du Campanien[1]. En 2005, Siverson et Lindgren notent que l'âge des plus récents fossiles de Cretoxyrhina diffère d'un continent à l'autre. En Australie, les plus récents fossiles connus de Cretoxyrhina sont datés d'environ 83 millions d'années (Santonien), les plus jeunes fossiles nord-américains datent du Campanien précoce, tandis que les plus jeunes fossiles connus d'Europe sont au moins deux millions d'années plus jeunes. Ces différences entre les âges suggèrent que Cretoxyrhina a pu s'éteindre localement au fil du temps jusqu'à ce que le genre dans son ensemble disparaisse[2].
Le déclin de Cretoxyrhina coïncide avec l'émergence de nouveaux prédateurs tels que Tylosaurus, ce qui suggère que la pression croissante de la concurrence avec le mosasaure et d'autres prédateurs de niveau trophique similaire peut avoir joué un rôle majeur dans le déclin puis l'extinction de Cretoxyrhina. Une autre cause possible est la diminution progressive de la profondeur et le rétrécissement de la voie maritime intérieure de l'Ouest, qui aurait entrainé la disparition de l'environnement pélagique privilégié du requin, ce qui n'explique cependant pas le déclin et l'extinction de Cretoxyrhina ailleurs dans le monde[37]. Enfin, il est possible que l'extinction de Cretoxyrhina ait libéré des niches écologiques ensuite occupées par les grands mosasaures[91].
Notes et références
Notes
- Ceux-ci sont maintenant identifiés sous les noms de Cosmopolitodus hastalis et Isurus desori[14], et Oxyrhina xiphodon est maintenant considérée comme conspécifique de Cosmopolitodus hastalis[15].
- L'orthographe originale choisie par Agassiz se termine par -ii[13]. Les auteurs ultérieurs ont laissé tomber la lettre supplémentaire, l'épelant comme mantelli. Bien qu'il s'agisse d'une faute d'orthographe claire, elle est maintenue comme l'orthographe valide de l'espèce selon l'article 33.4 de la CINZ en raison de son utilisation prédominante par rapport à l'originale[16].
- Ce taxon est un nomen dubium dont les spécimens référés représentent désormais Cardabiodon et Dwardius[8].
- Pour une raison peu claire, cette étude fut d'abord publiée en ligne sur BioOne en 2013, avant d'être officiellement republiée deux ans plus tard dans la revue scientifique polonaise Acta Palaeontologica Polonica[10].
- Ces estimations de vitesse de nage ont été calculées sur la base de modèles hydrodynamiques. Cependant, certaines recherches suggèrent que les poissons actuels peuvent être incapables de nager jusqu'à de tels taux calculés en raison des limitations physiologiques imprévues par une telle modélisation hydrodynamique. Par exemple, Morten B. S. Svendsen et ses collègues en 2016 constatent que les balises de l'accéléromètre et les analyses vidéo à grandes vitesses suggèrent que les vitesses maximales du marlin sont plus proches de 36 à 54 km/h contrairement aux estimations calculées de 126 km/h à partir de la modélisation hydrodynamique traditionnelle, car des vitesses plus élevées que les premières devaient causer de graves dommages aux tissus des nageoires. Il est également observé que d'autres poissons nageant rapidement ont des limitations similaires[46]. De même, l'étude de Jayson M. Semmens et ses collègues en 2019 observe que les grands requins blancs ne dépassent pas des vitesses de pointe de 23,4 km/h lorsqu'ils attaquent des phoques[47].
- Ce syntype réside dans la collection du musée d'histoire naturelle de Londres et est catalogué NHMUK PV OR 4498[49]. Ces détails ne sont pas mentionnés dans l'étude de Newbrey et al. (2013)[10].
- Le tableau est basé uniquement sur les vertèbres de FHSM VP-2187 et ne représente pas le genre Cretoxyrhina dans son ensemble[32].
- 'Isurus appendiculatus' et 'Isurus mantelli' sont des noms scientifiques proposés et utilisés par l'étude de 2014 de Diedrich. Les noms scientifiques originaux et plus largement acceptés de ces taxons sont indiqués entre parenthèses[51].
- L'étude de Shimada publié en 1997 est citée dans l'étude de 2014 de Diedrich. Ce dernier n'utilise la citation dans aucune de ses affirmations concernant le placement systématique de C. mantelli, ne l'utilisant que dans des notes stratigraphiques et historiques[51].
- Cette carte ne montre que certaines localités mentionnées dans la littérature scientifique et ne représente pas toutes les trouvailles connues concernant Cretoxyrhina.
Références
- (en) Kenshu Shimada, « Stratigraphic Record of the Late Cretaceous Lamniform Shark, Cretoxyrhina mantelli (Agassiz), in Kansas », Transactions of the Kansas Academy of Science, vol. 100, nos 3-4, , p. 139–149 (DOI 10.2307/3628002, JSTOR 3628002, S2CID 87585388)
- (en) Mikael Siverson et Johan Lindgren, « Late Cretaceous sharks Cretoxyrhina and Cardabiodon from Montana, USA », Acta Palaeontologica Polonica, vol. 50, no 2, , p. 301-314 (résumé, lire en ligne [PDF])
- (en) David Ward, Fossils of the Gault Clay – Sharks and Rays, The Paleontological Association, , 279-299 p. (lire en ligne)
- (en) James G. Ogg et Linda A. Hinnov, « Chapter 27 - Cretaceous », dans Felix M. Gradstein, James G. Ogg, Mark D. Schmitz et Gabi M. Ogg, The Geologic Time Scale, Oxford, Elsevier, , 1144 p. (ISBN 978-0-444-59425-9, DOI 10.1016/B978-0-444-59425-9.00027-5, S2CID 127523816), p. 793-853
- (en) Todd Cook, Eric Brown, Patricia E. Ralrick et Takuya Konish., « A late Campanian euselachian assemblage from the Bearpaw Formation of Alberta, Canada: some notable range extensions », Canadian Journal of Earth Sciences, vol. 54, no 9, , p. 973-980 (DOI 10.1139/cjes-2016-0233, Bibcode 2017CaJES..54..973C, hdl 1807/77762
 , S2CID 134789628)
, S2CID 134789628) - (en) Derek William Larson, Donald B. Brinkman et Phil R. Bell, « Faunal assemblages from the upper Horseshoe Canyon Formation, an early Maastrichtian cool-climate assemblage from Alberta, with special reference to the Albertosaurus sarcophagus bonebed », Canadian Journal of Earth Sciences, vol. 47, no 9, , p. 1159-1181 (DOI 10.1139/E10-005, Bibcode 2010CaJES..47.1159L, S2CID 129391823)
- (de) Charles R. Eastman, « Beiträge zur Kenntniss der Gattung Oxyrhina: mit besonderer Berücksichtigung von Oxyrhina mantelli Agassiz », Palaeontographica, E. Schweizerbart'sche Verlagsbuchhandlung, , p. 149-191 (lire en ligne)
- (en) Mikael Siverson, David John Ward, Johan Lindgren et L. Scott Kelly, « Mid-Cretaceous Cretoxyrhina (Elasmobranchii) from Mangyshlak, Kazakhstan and Texas, USA », Alcheringa: An Australasian Journal of Palaeontology, vol. 37, no 1, , p. 1-18 (DOI 10.1080/03115518.2012.709440, S2CID 129785545, lire en ligne)
- (en) Phillip Sternes et Kenshu Shimada, « Caudal fin of the Late Cretaceous shark, Cretoxyrhina mantelli (Lamniformes: Cretoxyrhinidae), morphometrically compared to that of extant lamniform sharks », Society of Vertebrate Paleontology, (lire en ligne [archive du ], consulté le )
- (en) Michael G. Newbrey, Mikael Siverson, Todd D. Cook, Allison M. Fotheringham et Rebecca L. Sanchez, « Vertebral Morphology, Dentition, Age, Growth, and Ecology of the Large Lamniform Shark Cardabiodon ricki », Acta Palaeontologica Polonica, vol. 60, no 4, , p. 877-897 (DOI 10.4202/app.2012.0047 , S2CID 56258221)
- (en) Jacopo Amalfitano, Luca Giusberti, Eliana Fornaciari, Fabio Marco Dalla Vicchia, Valeria Luciani, Jürgen Kriwet et Giorgio Carnevale, « Large deadfall of the 'ginsu' shark Cretoxyrhina mantelli (Agassiz, 1835) (Neoselachii, Lamniformes) from the Upper Cretaceous of northeastern Italy », Cretaceous Research, vol. 98, , p. 250-275 (DOI 10.1016/j.cretres.2019.02.003, hdl 11392/2402988, S2CID 135196193, lire en ligne)
- (en) Gideon Mantell et Mary Ann Mantell, The fossils of the South Downs; or illustrations of the geology of Sussex, Lupton Relfe, (PMCID 128930354, DOI 10.3931/e-rara-16021, S2CID 128930354, lire en ligne)
- Louis Agassiz, Recherches sur les poissons fossiles, vol. 3, Museum of Comparative Zoology, 1833-1843 (lire en ligne)
- (en) László Kocsis, « Central Paratethyan shark fauna (Ipolytarnóc, Hungary) », Geologica Carpathica, vol. 58, no 1, , p. 27-40 (lire en ligne [archive du ], consulté le )
- (en) David J. Ward et Charles G. Bonavia, « Additions to and a review of the Miocene Shark and Ray fauna of Malta », The Central Mediterranean Naturalist, vol. 3, no 3, , p. 131-146 (lire en ligne)
- Bruno Cahuzac, Sylvain Adnet, Henri Cappetta et Romain Vullo, « Les espèces et genres de poissons sélaciens fossiles (Crétacé, Tertiaire) créés dans le Bassin d'Aquitaine ; recensement, taxonomie », Bulletin de la Société Linnéenne de Bordeaux, vol. 142, no 45, , p. 3-43 (lire en ligne)
- Mike Everhart, « A Giant Ginsu Shark » [archive du ], sur Oceans of Kansas, (consulté le )
- (en) Jim Bourdon, « Cretoxyrhina » [archive du ] (consulté le )
- (en) Joseph Leidy, « Fossil Vertebrates », Contributions to the Extinct Vertebrate Fauna of the Western Territories, vol. 1, , p. 302-303 (lire en ligne)
- (en) « FHSM VP 323 », Fort Hays State University's Sternberg Museum of Natural History
- (en) « FHSM VP 2187 », Fort Hays State University's Sternberg Museum of Natural History
- (en) Mike Everhart, « Large Sharks in the Western Interior Sea » [archive du ], sur Oceans of Kansas, (consulté le )
- (en) « Kansas University Vertebrate Paleontology Database », Kansas University Biodiversity Institute and Natural History Museum
- (en + ru) E. V. Popov, « An annotated bibliography of the Soviet paleoichthyologist Leonid Glickman (1929–2000) », Proceedings of the Zoological Institute of the Russian Academy of Sciences, vol. 320, no 1, , p. 25–49 (DOI 10.31610/trudyzin/2016.320.1.25 , S2CID 135042023)
- (ru) Leonid Glickman, « Rates of evolution in Lamoid sharks », Doklady Akademii Nauk SSSR, vol. 123, , p. 568-571 (lire en ligne)
- (en) W. James Kennedy, Christopher King et David J. Ward, « The Upper Albian and Lower Cenomanian succession at Kolbay, eastern Mangyshlak, southwest Kazakhstan », Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, vol. 78, , p. 117-147 (lire en ligne)
- (ru) V.I. Zhelezko, « The evolution of teeth systems of sharks of Pseudoisurus Gluckman, 1957 genus – the biggest pelagic sharks of Eurasia », Materialy Po Stratigrafii I Paleontologii Urala, vol. 1, no 4, , p. 136-141 (lire en ligne [PDF])
- (en) Charlie J. Underwood et Stephen L. Cumbaa, « Chondrichthyans from a Cenomanian (Late Cretaceous) bonebed, Saskatchewan, Canada », Palaeontology, vol. 53, no 4, , p. 903-944 (DOI 10.1111/j.1475-4983.2010.00969.x , S2CID 129260151)
- (en) Kenshu Shimada, « Dentition of the Late Cretaceous lamniform shark, Cretoxyrhina mantelli from the Niobrara Chalk of Kansas », Journal of Vertebrate Paleontology, vol. 17, no 2, , p. 269-279 (DOI 10.1080/02724634.1997.10010974, JSTOR 4523806, S2CID 129515433)
- (en) Kenshu Shimada, « Skeletal Anatomy of the Late Cretaceous Lamniform Shark, Cretoxyrhina mantelli, from the Niobrara Chalk in Kansas », Journal of Vertebrate Paleontology, vol. 17, no 4, , p. 642-652 (DOI 10.1080/02724634.1997.10011014, JSTOR 4523854, S2CID 84706874)
- (en) Kenshu Shimada, « Paleoecological relationships of the Late Cretaceous lamniform shark, Cretoxyrhina mantelli (Agassiz) », Journal of Paleontology, vol. 71, no 5, , p. 926–933 (DOI 10.1017/S002233600003585X, JSTOR 1306568, S2CID 132634254)
- (en) Kenshu Shimada, « Ontogenetic parameters and life history strategies of the late Cretaceous lamniform shark, Cretoxyrhina mantelli, based on vertebral growth increments », Journal of Vertebrate Paleontology, vol. 28, , p. 21-33 (DOI 10.1671/0272-4634(2008)28[21:OPALHS]2.0.CO;2, JSTOR 30126330, S2CID 85777970)
- (en) Kenshu Shimada, « Phylogeny of lamniform sharks (Chondrichthyes: Elasmobranchii) and the contribution of dental characters to lamniform systematics », Paleontological Research, vol. 9, , p. 55-72 (DOI 10.2517/prpsj.9.55, S2CID 84715232)
- (en) Warren Upham, The Geology of Central and Western Minnesota. A Preliminary Report. [From the General Report of Progress for the Year 1879.], St. Paul, Minnesota, U.S.A., The Pioneer Press Co., (lire en ligne), p. 18
- (en) Mike Everhart, « Cretoxyrhina mantelli, the Ginsu Shark » [archive du ], sur Oceans of Kansas, (consulté le )
- (en) Kenshu Shimada, « Dental homologies in lamniform sharks (Chondrichthyes: Elasmobranchii) », Journal of Morphology, vol. 251, no 1, , p. 38-72 (PMID 11746467, DOI 10.1002/jmor.1073, S2CID 22195080)
- (en) Jim Bourdon et Michael Everhart, « Analysis of an associated Cretoxyrhina mantelli dentition from the Late Cretaceous (Smoky Hill Chalk, Late Coniacian) of western Kansas », Transactions of the Kansas Academy of Science, vol. 114, nos 1–2, , p. 15-32 (DOI 10.1660/062.114.0102, S2CID 56237457, lire en ligne [archive du ] [PDF], consulté le )
- (en) Kenshu Shimada et David J. Cicimurri, « Skeletal anatomy of the Late Cretaceous shark, Squalicorax (Neoselachii, Anacoracidae) », Paläontologische Zeitschrift, vol. 79, no 2, , p. 241-261 (DOI 10.1007/BF02990187, S2CID 83861009, lire en ligne)
- (en) Mike Everhart, « Sharks of Kansas » [archive du ], sur Oceans of Kansas (consulté le )
- (en) Denton Lee O’Neal, « Chemostratigraphic and depositional characterization of the Niobrara Formation, Cemex Quarry, Lyons, CO », Colorado School of Mines, (lire en ligne [archive du ] [PDF], consulté le )
- (en) Kenshu Shimada, Stephen L. Cumbaa et Deanne van Rooyen, « Caudal Fin Skeleton of the Late Cretaceous Lamniform shark, Cretoxyrhina mantelli, from the Niobrara Chalk of Kansas », New Mexico Museum of Natural History Bulletin, vol. 35, , p. 185-191 (lire en ligne)
- (en) Humberto G. Ferron, « Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks », PLOS ONE, vol. 12, no 9, , e0185185 (PMID 28938002, PMCID 5609766, DOI 10.1371/journal.pone.0185185 , Bibcode 2017PLoSO..1285185F)
- (en) John K. Carlson, Kenneth J. Goldman et Christopher G. Lowe, « Metabolism, Energetic Demand, and Endothermy », dans Jeffrey C. Carrier, John A. Musick et Michael R. Heithaus, Biology of sharks and their relatives, Boca Raton, CRC Press, , 596 p. (ISBN 978-0-8493-1514-5), p. 203–224
- (en) Sun H. Kim, Kenshu Shimada et Cynthia K. Rigsby, « Anatomy and Evolution of Heterocercal Tail in Lamniform Sharks », The Anatomical Record, vol. 296, no 3, , p. 433-442 (PMID 23381874, DOI 10.1002/ar.22647 , S2CID 205411092)
- (en) Bruce A. Wright, Alaska's Great White Sharks, Top Predator Publishing Company, , 115 p. (ISBN 978-0-615-15595-1, lire en ligne), p. 27
- (en) Morten B. S. Svendsen, Paolo Domenici, Stefano Marras, Jens Krause, Kevin M. Boswell, Ivan Rodriguez-Pinto, Alexander D. M. Wilson, Ralf H. J. M. Kurvers, Paul E. Viblanc, Jean S. Finger et John F. Steffensen, « Maximum swimming speeds of sailfish and three other large marine predatory fish species based on muscle contraction time and stride length: a myth revisited », Biology Open, vol. 5, no 10, , p. 1415-1419 (ISSN 2046-6390, PMID 27543056, PMCID 5087677, DOI 10.1242/bio.019919 )
- (en) Jayson M. Semmens, Alison A. Kock, Yuuki Y. Watanabe, Charles M. Shepard, Eric Berkenpas, Kilian M. Stehfest, Adam Barnett et Nicholas L. Payne, « Preparing to launch: biologging reveals the dynamics of white shark breaching behaviour », Marine Biology, vol. 166, no 7, (ISSN 0025-3162, DOI 10.1007/s00227-019-3542-0, S2CID 196664873)
- (en) C. Díez, M. Soto et J. M. Blanco, « Biological characterization of the skin of shortfin mako shark Isurus oxyrinchus and preliminary study of the hydrodynamic behaviour through computational fluid dynamics », Journal of Fish Biology, vol. 87, no 1, , p. 123-137 (PMID 26044174, DOI 10.1111/jfb.12705, S2CID 206057366)
- (en) « PV OR 4498 », sur Natural History Museum Specimen Collection, (consulté le )
- (en) Henry F. Mollet et Gregor M. Cailliet, « Using allometry to predict body mass from linear measurements of the white shark », dans A. Peter Klimley & David G. Ainley, Great white sharks : the biology of Carcharodon carcharias, San Diego, Academic Press, , PDF (ISBN 978-0-12-415031-7, DOI 10.1016/B978-012415031-7/50010-0, S2CID 88647451, lire en ligne), p. 81–89
- (en) C. G. Diedrich, « Skeleton of the Fossil Shark Isurus denticulatus from the Turonian (Late Cretaceous) of Germany – Ecological Coevolution with Prey of Mackerel Sharks », Paleontology Journal, vol. 2014, , p. 1-20 (DOI 10.1155/2014/934235 , S2CID 129451357)
- (en) Kensu Shimada, Martin A. Becker et Michael L. Griffiths, « Body, jaw, and dentition lengths of macrophagous lamniform sharks, and body size evolution in Lamniformes with special reference to 'off-the-scale' gigantism of the megatooth shark, Otodus megalodon », Historical Biology, vol. 33, no 11, , p. 1-17 (DOI 10.1080/08912963.2020.1812598, S2CID 224935604)
- (en) J. G. Maisey, « What is an 'elasmobranch'? The impact of palaeontology in understanding elasmobranch phylogeny and evolution », Journal of Fish Biology, vol. 80, no 5, , p. 918-951 (PMID 22497368, DOI 10.1111/j.1095-8649.2012.03245.x, S2CID 12512063)
- (en) Mikael Siverson, « A new large lamniform shark from the uppermost Gearle Siltstone (Cenomanian, Late Cretaceous) of Western Australia », Transactions of the Royal Society of Edinburgh: Earth Sciences, vol. 90, , p. 49-66 (DOI 10.1017/S0263593300002509, S2CID 131195702)
- (en) Kenshu Shimada, « Skeletal and Dental Anatomy of Lamniform shark Cretalamna appendiculata, from Upper Cretaceous Niobrara Chalk of Kansas », Journal of Vertebrate Paleontology, vol. 27, no 3, , p. 584-602 (DOI 10.1671/0272-4634(2007)27[584:SADAOL]2.0.CO;2, S2CID 131264105)
- (en) Mark Wilson, Mark V. H. Wilson, Alison M. Murray, A. Guy Plint, Michael G. Newbrey et Michael J. Everhart, « A high-latitude euselachian assemblage from the early Turonian of Alberta, Canada », Journal of Systematic Palaeontology, vol. 11, no 5, , p. 555-587 (DOI 10.1080/14772019.2012.707990, S2CID 129551937, lire en ligne)
- (en) Anne Sørensen, Finn Surlyk et Johan Lindgren, « Food resources and habitat selection of a diverse vertebrate fauna from the upper lower Campanian of the Kristianstad Basin, southern Sweden », Cretaceous Research, vol. 42, , p. 85-92 (DOI 10.1016/j.cretres.2013.02.002, S2CID 140653224)
- (en) Mike Everhart, « Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the lamniform shark, Cretoxyrhina mantelli », PalArch's Journal of Vertebrate Paleontology, vol. 2, no 2, (lire en ligne [archive du ] [PDF], consulté le )
- (en) Kenshu Shimada et G. E. Hooks, « Shark-bitten protostegid turtles from the Upper Cretaceous Mooreville Chalk, Alabama », Journal of Paleontology, vol. 78, , p. 205-210 (DOI 10.1666/0022-3360(2004)078<0205:SPTFTU>2.0.CO;2, S2CID 129391058)
- (en) Mike Everhart, « Parts is parts and pieces is pieces » [archive du ], sur Oceans of Kansas, (consulté le )
- (en) Mark P. Witton, « Pterosaurs in Mesozoic food webs: a review of fossil evidence », Geological Society, London, Special Publications, vol. 455, no 1, , p. 7-23 (DOI 10.1144/SP455.3, Bibcode 2018GSLSP.455....7W, S2CID 90573936)
- (en) David W.E. Hone, Mark P. Witton et Michael B. Habib, « Evidence for the Cretaceous shark Cretoxyrhina mantelli feeding on the pterosaur Pteranodon from the Niobrara Formation », PeerJ, vol. 6, no e6031, , e6031 (PMID 30581660, PMCID 6296329, DOI 10.7717/peerj.6031 )
- (en) Mike Everhart et Keith Ewell, « Shark-bitten dinosaur (Hadrosauridae) caudal vertebrae from the Niobrara Chalk (Upper Coniacian) of western Kansas », Kansas Academy of Science, vol. 109, , p. 27-35 (DOI 10.1660/0022-8443(2006)109[27:SDHCVF]2.0.CO;2, S2CID 86366930)
- (en) Mike J. Everhart et Shawn A. Hamm, « A new nodosaur specimen (Dinosauria: Nodosauridae) from the Smoky Hill Chalk (Upper Cretaceous) of western Kansas », Kansas Academy of Science, vol. 108, , p. 15-21 (DOI 10.1660/0022-8443(2005)108[0015:ANNSDN]2.0.CO;2, S2CID 86252250, lire en ligne)
- (en) Iyad S. Zalmout et Hakam Mustafa, « A selachian fauna from the Late Cretaceous of Jordan », Abhath Al-Yarmouk, vol. 10, no 2B, , p. 377-343 (lire en ligne)
- (en) Andrew Retzler, Mark A. Wilson et Yoav Avnib, « Chondrichthyans from the Menuha Formation (Late Cretaceous: Santonian–Early Campanian) of the Makhtesh Ramon region, southern Israel », Cretaceous Research, vol. 40, , p. 81-89 (DOI 10.1016/j.cretres.2012.05.009, S2CID 128754632)
- (en) Stephen L. Cumbaa, Kenshu Shimada et Todd D. Cook, « Mid-Cenomanian vertebrate faunas of the Western Interior Seaway of North America and their evolutionary, paleobiogeographical, and paleoecological implications », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 295, nos 1-2, , p. 199-214 (DOI 10.1016/j.palaeo.2010.05.038, Bibcode 2010PPP...295..199C, S2CID 129427657)
- (en) Alison M. Murray et Todd D. Cook, « Overview of the Late Cretaceous fishes of the Northern Western Interior Seaway », Cretaceous Period: Biotic Diversity and Biogeography, vol. 71, , p. 255-261 (lire en ligne [archive du ], consulté le )
- (en) Oliver Friedrich, Richard D. Norris et Jochen Erbacher, « Evolution of middle to Late Cretaceous oceans – A 55 m.y. record of Earth's temperature and carbon cycle », Geology, vol. 40, no 2, , p. 107-110 (DOI 10.1130/G32701.1, Bibcode 2012Geo....40..107F, S2CID 131578034)
- (en + fr) Nathalie Bardet, Alexandra Houssaye, Jean-Claude Rage et Xabier Pereda Suberbiola, « La radiation cénomano-turonienne (Crétacé supérieur) des squamates (Reptilia) marins : le rôle de la Téthys méditerranéenne », Bulletin de la Société Géologique de France, vol. 179, no 6, , p. 605-622 (DOI 10.2113/gssgfbull.179.6.605, S2CID 140669678, lire en ligne)
- (en) In Brief, « Submarine eruption bled Earth's oceans of oxygen » [archive du ], New Scientist, (consulté le )
- (en) L. Barry Albright III, David D. Gillette et Alan L. Titus, « Plesiosaurs from the Upper Cretaceous (Cenomanian-Turonian) tropic shale of southern Utah, Part 1: New records of the pliosaur Brachauchenius lucasi », Journal of Vertebrate Paleontology, vol. 27, no 1, , p. 31-40 (DOI 10.1671/0272-4634(2007)27[31:PFTUCC]2.0.CO;2, JSTOR 4524665, S2CID 128994726, lire en ligne)
- (en) Elizabeth L. Nicholls et Anthony P. Russel, « Paleobiogeography of the Cretaceous Western Interior Seaway of North America: the vertebrate evidence », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 79, nos 1–2, , p. 149-169 (DOI 10.1016/0031-0182(90)90110-S, Bibcode 1990PPP....79..149N, S2CID 129177959)
- (en) Référence Fossilworks : Cretoxyrhina
- (en) Boris Ekrt, Martin Košt'ák, Martin Mazuch, Silke Voigt et Frank Wiese, « New records of teleosts from the Late Turonian (Late Cretaceous) of the Bohemian Cretaceous Basin (Czech Republic) », Cretaceous Research, vol. 29, no 4, , p. 659-673 (DOI 10.1016/j.cretres.2008.01.013, S2CID 129497814)
- (en) Mikael Siverson, « Lamniform sharks of the Mid Cretaceous Alinga Formation and Beedagong Claystone, Western Australia », Palaeontology, vol. 39, no 4, , p. 813-849 (lire en ligne [PDF])
- (en) Annie P. McIntosh, Kenshu Shimada et Michael J. Everhart, « Late Cretaceous Marine Vertebrate Fauna from the Fairport Chalk Member of the Carlile Shale in Southern Ellis County, Kansas, U.S.A. », Transactions of the Kansas Academy of Science, vol. 119, no 2, , p. 222-230 (DOI 10.1660/062.119.0214, S2CID 88452573)
- (en) Christopher Gallardo, Kenshu Shimada et Bruce A. Shumacher, « A New Late Cretaceous Marine Vertebrate Assemblage from the Lincoln Limestone Member of the Greenhorn Limestone in Southeastern Colorado », Transactions of the Kansas Academy of Science, vol. 115, nos 3-4, , p. 107-116 (DOI 10.1660/062.115.0303, S2CID 86258761)
- (en) Nikita V. Zelenkov, Alexander O. Averianov et Evgeny V. Popov, « An Ichthyornis-like bird from the earliest Late Cretaceous (Cenomanian) of European Russia », Cretaceous Research, vol. 75, no 1, , p. 94-100 (DOI 10.1016/j.cretres.2017.03.011, S2CID 132491078)
- (en) Andrei V. Panteleyev, Evgenii V. Popov et Alexander O. Averianov, « New record of Hesperornis rossicus (Aves, Hesperornithiformes) in the Campanian of Saratov Province, Russia », Paleontological Research, vol. 8, no 2, , p. 115-122 (DOI 10.2517/prpsj.8.115 , S2CID 54659655)
- (en) Akihiro Misaki, Hideo Kadota et Haruyoshi Maeda, « Discovery of mid-Cretaceous ammonoids from the Aridagawa area, Wakayama, southwest Japan », Paleontological Research, vol. 12, no 1, , p. 19-26 (DOI 10.2517/1342-8144(2008)12[19:DOMAFT]2.0.CO;2, S2CID 129158175)
- (en) Mikael Siverson et Marcin Machalski, « Late late Albian (Early Cretaceous) shark teeth from Annopol, Pol », Alcheringa: An Australasian Journal of Palaeontology, vol. 41, no 4, , p. 433-463 (DOI 10.1080/03115518.2017.1282981, S2CID 133123002)
- (en) Samuel Giersch, Eberhard Frey, Wolfgang Stinnesbeck et Arturo H. Gonzalez, « Fossil fish assemblages of northeastern Mexico: New evidence of Mid Cretaceous Actinopterygian radiation », 6th Meeting of the European Association of Vertebrate Paleontologists, , p. 43-45 (lire en ligne [PDF])
- (en) Naoshu Kitamura, « Features and paleoecological significance of the shark fauna from the Upper Cretaceous Hinoshima Formation, Himenoura Group, Southwest Japan », Paleontological Research, vol. 23, no 2, , p. 110-130 (DOI 10.2517/2014PR020, S2CID 129036582, lire en ligne [PDF])
- (en) Octávio MateusI, Louis L. Jacobs, Anne S. Schulp, Michael J. Polcyn, Tatiana S. Tavares, André Buta Neto, Maria Luísa Morais et Miguel T. Antunes, « Angolatitan adamastor, a new sauropod dinosaur and the first record from Angola », Anais da Academia Brasileira de Ciências, vol. 83, no 1, , p. 221-233 (PMID 21437383, DOI 10.1590/S0001-37652011000100012 , S2CID 24000235, lire en ligne [PDF])
- (en) Guillaume Guinot et Jorge D. Carrillo-Briceno, « Lamniform sharks from the Cenomanian (Upper Cretaceous) of Venezuela », Cretaceous Research, vol. 82, no 2018, , p. 1-20 (DOI 10.1016/j.cretres.2017.09.021, S2CID 135126387, lire en ligne [PDF])
- (en + es + de) Oliver Hampe, « Considerations on a Brachauchenius skeleton (Pliosauroidea) from the lower Paja Formation (late Barremian) of Villa de Leyva area (Colombia) », Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe, vol. 8, , p. 37-51 (DOI 10.1002/mmng.200410003 , S2CID 73535826)
- (en) B. A. Schumacher, K. Carpenter et M. J. Everhart, « A new Cretaceous Pliosaurid (Reptilia, Plesiosauria) from the Carlile Shale (middle Turonian) of Russell County, Kansas », Journal of Vertebrate Paleontology, vol. 33, no 3, , p. 613-628 (DOI 10.1080/02724634.2013.722576, JSTOR 42568544, S2CID 130165209, lire en ligne )
- (en) Kenshu Shimada et Tracy K. Ystesund, « A dolichosaurid lizard, Coniasaurus cf. C. crassidens, from the Upper Cretaceous Carlile Shale in Russell County, Kansas », Transactions of the Kansas Academy of Science, vol. 110, nos 3-4, , p. 236-242 (DOI 10.1660/0022-8443(2007)110[236:ADLCCC]2.0.CO;2, S2CID 86027583)
- (en) Boris Ekrt, Martin Košťák, Martin Mazuch et J. Valiceck, « Short note on new records of late Turonian (Upper Cretaceous) marine reptile remains from the Upohlavy quarry (NW Bohemia, Czech Republic) », Bulletin of the Czech Geological Survey, vol. 76, no 2, , p. 101-106 (lire en ligne )
- (en) Michael Everhart, « Rapid evolution, diversification and distribution of mosasaurs (Reptilia; Squamata) prior to the KT Boundary », Tate 2005 11th Annual Symposium in Paleontology and Geology, (lire en ligne )
- (en) Corinne Myers et Bruce Lieberman, « Sharks that pass in the night: using Geographical Information Systems to investigate competition in the Cretaceous Western Interior Seaway », Proceedings of the Royal Society B, vol. 278, no 1706, , p. 681-689 (PMID 20843852, PMCID 3030853, DOI 10.1098/rspb.2010.1617, lire en ligne [PDF])
Citations originales
- « The remarkable thing about this specimen is that the vertebral column, though of cartilaginous material, was almost complete, and that the large number of 250 teeth were in position. When Chas. R. Eastman, of Harvard, described this specimen, it proved so complete as to destroy nearly thirty synonyms used to name the animal, and derived from many teeth found in former times. »
Voir aussi
Articles connexes
Liens externes
- Riley Black, « Quand les requins se nourrissaient de dinosaures » [archive du ] [html], sur National Geographic,
- (en) « Dossier d'information sur Cretoxyrhina mantelli » [archive du ] [html], sur National Geographic
- (en + fr) Mike Everhart, « Les Grands Requins de la Mer Intérieure de l'Ouest » [archive du ] [html], sur Oceans of Kansas
- Ressources relatives au vivant :