Cardabiodon
Cardabiodon (littéralement « dent de Cardabia ») est un genre éteint de grands requins lamniformes ayant vécu durant le Crétacé supérieur (âges Cénomanien et Turonien), il y a entre 95 et 91 millions d'années. Il fait partie des Cardabiodontidae, une famille de lamniformes caractérisée par des structures dentaires spécifiques. Le genre compte deux espèces, Cardabiodon ricki et Cardabiodon venator, dont les restes fossiles ont été découverts en Australie, en Amérique du Nord, en Angleterre et au Kazakhstan. Il s'agissait probablement d'un requin antitropical qui habitait les océans tempérés néritiques et hauturiers situés entre 40° et 60° de paléolatitude, comme le requin-taupe commun actuel.
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Classe | Chondrichthyes |
| Infra-classe | Euselachii |
| Super-ordre | Selachimorpha |
| Ordre | Lamniformes |
| Famille | † Cardabiodontidae |
Espèces de rang inférieur
- † C. ricki (type) Siverson, 1999
- † C. venator Siverson & Lindgren, 2005
- † Pseudoisurus tomosus Glückman, 1957
- † Cretolamna woodwardi Williamsen et al., 1993
Mesurant jusqu'à 5,5 m de longueur et d'un profil plutôt trapu, Cardabiodon figurait parmi les plus grands requins de son temps. Il était probablement un prédateur au sommet de son écosystème et utilisait probablement ses grandes dents robustes et ses capacités de nage rapide pour s'attaquer à une variété de proies, notamment des plésiosaures, des ichtyosaures et d'autres gros poissons.
Découverte et dénomination
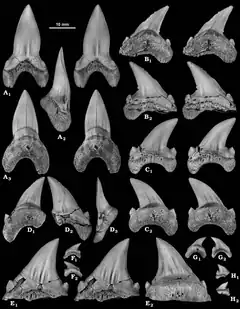
Cardabiodon a été décrit en 1999 par le paléontologue Mikael Siversson (d) à partir de fossiles découverts dans le bassin sud de Carnarvon du Gearle Siltstone, à Cardabia (en), une ferme d'élevage de bétail située en Australie-Occidentale[1]. Ce site est daté de l'âge Cénomanien du Crétacé supérieur, il y a entre 96 et 94,7 millions d'années[2]. Ce spécimen, catalogué WAM 96.4.45, consistait en un ensemble d'une centaine de dents et de quinze vertèbres et représentait un requin subadulte. Des fouilles ultérieures sur le même site ont collecté quatre autres dents appartenant au même spécimen[3].
Siversson a reconnu le spécimen comme holotype d'un nouveau genre et lui a attribué le nom scientifique Cardabiodon ricki. Le nom générique Cardabiodon combine le nom du lieu originaire de la trouvaille, Cardabia, et le terme en grec ancien ὀδών / odṓn « dent », le tout signifiant « dent de Cardabia ». L'épithète spécifique de l'espèce type, ricki, rend hommage à Rick French, le propriétaire de la ferme de Cardabia[1]. En 2005, la deuxième espèce, C. venator, a été décrite à partir de spécimens types comptant un total de 37 dents, collectées sur un site du membre Fairport des schistes de Carlile, situés dans le Montana, une formation géologique datée de 92 à 91 millions d'années. L'épithète spécifique de cette espèce, venator, est un mot latin signifiant « chasseur », ce qui fait référence à la position présumée de C. venator en tant que superprédateur, ou en tout cas proche du sommet de la chaine alimentaire[2].
Avant la description de Cardabiodon par Siverson, ses dents avaient été décrites sous deux noms désormais synonymes. Une description avait été faite en 1957 par le paléontologue soviétique Leonid Glickman, dans laquelle il décrivait le taxon Pseudoisurus tomosus, sur la base de quatre dents provenant de l'oblast de Saratov, en Russie européenne[4]. Plus tôt, en 1894, Arthur Smith Woodward avait décrit un ensemble de vingt-cinq dents datant du Crétacé, découvertes autour de Maidstone, en Angleterre, sous le nom 'Lamna appendiculata' (Cretolamna appendiculata)[5]. En 1977, le paléontologue Jaques Herman a estimé que ce dernier représentait une nouvelle espèce, qu'il a nommée Cretolamna woodwardi[6]. En 1996, Siverson a fusionné Cretolamna woodwardi et Pseudoisurus tomosus, jugeant que leurs spécimens types référencés représentaient la même espèce[7]. En 1999, Siverson a réévalué la taxonomie de Pseudoisurus tomosus, en concluant que ses syntypes avaient été perdus, ce qui en faisait un nomen dubium. Il constatait également que Cretolamna woodwardi représentait en fait le mélange de deux genres distincts de requins, ce qui semblait être également le cas de Pseudoisurus tomosus. Le premier a été renommé Cardabiodon ricki (qu'il avait déjà décrit en utilisant le spécimen WAM 96.4.45), et le second a été renommé Dwardius woodwardi[1].
Description
Morphologie
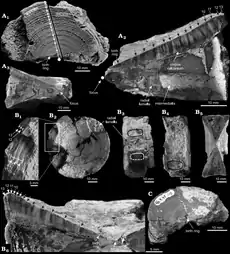
C. ricki aurait mesuré jusqu'à 5,5 m de longueur, d'après la comparaison de ses vertèbres avec celles du grand requin blanc moderne, tandis que C. venator aurait atteint 5 m d'après la taille de ses dents[3], ce qui fait de Cardabiodon l'un des plus grands requins connus[1]. Le registre fossile de ce taxon est très mince : il comprend des dents, des vertèbres et des écailles. Cette rareté est usuelle car le squelette cartilagineux des requins se fossilise difficilement. Les vertèbres peuvent parfois se conserver quand elles sont durcies par calcification[8].
La peau de Cardabiodon est recouverte d'écailles placoïdes émailoïdes en forme de larme, revêtues de 6 à 8 rainures parallèles, qui possèdent chacune une structure appelée kneel. Chez un individu mesurant 2,5 à 3,8 m de long, ces écailles varieraient de 0,3 à 0,7 mm dans les dimensions maximales. Les écailles avec rainures et kneels permettent de réduire la trainée hydrodynamique et d'augmenter la vitesse de nage, ce qui est un trait fréquent chez les lamniformes. Cardabiodon était donc probablement capable de nager vite et de traquer des proies rapides[8].
Le corps et les lamelles radiales, les deux couches les plus externes d'une vertèbre chez les lamniformes, permettent d'estimer le poids de l'animal en fonction de sa longueur, selon une relation mathématique établie sur la base des requins actuels. Les vertèbres de Cardabiodon sont de constitution robuste, plus épaisses que celles d'autres grands requins appartenant au même groupe, ce qui suggère que Cardabiodon était à longueur égale significativement plus lourd et plus trapu, bien qu'aucune estimation de son poids n'ait encore été publiée. Sa colonne vertébrale est rigide et en forme de fuseau, ce qui serait peu flexible mais efficace pour une nage rapide[3].
Denture
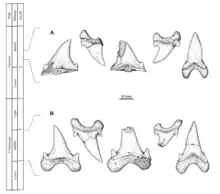
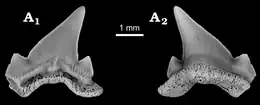
Cardabiodon possède les caractéristiques dentaires de base d'un requin lamniforme[3]. Sa denture est dite hétérodonte, ce qui signifie que la mâchoire contient des dents différenciées[1]. Les dents de Cardabiodon sont caractérisées par des racines fortement bilabiales, des couronnes robustes, qui sont soit presque symétriquement dressées, soit incurvées de manière distale, des cols de dents visibles et larges (bourlette), des bords tranchants non dentelés et des mini-cuspides latérales (petites cuspides émaillées qui apparaissent à la base de la couronne de la dent)[1] - [2] - [8] - [9]. La face linguale, c'est-à-dire la face des dents tournée vers la langue, présente une racine massivement convexe avec de petits pores nutritifs[9]. La face labiale de la couronne, c'est-à-dire tournée vers l'extérieur, présente une protubérance[10].
Les dents de Cardabiodon sont de taille moyenne par rapport à la taille de son corps[8] - [11]. Les plus grands exemplaires de dents documentés chez C. ricki proviennent de l'holotype[N 1]. Ses dents antérieures mesurent jusqu'à 3,67 cm de hauteur, tandis que ses premières dents postérieures et inférieures mesurent jusqu'à 3,98 cm de hauteur. Les dents de C. venator sont légèrement plus petites, la plus grande connue étant une dent antérieure mesurant 3,26 cm de hauteur, mais elles sont toutefois bien plus épaisses. C. venator a également des mini-cuspides latérales nettement plus courtes que C. ricki[2].
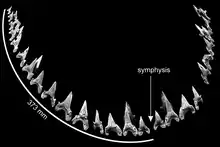
Siverson a reconstitué en 1999 la formule dentaire de C. ricki sur la base des 104 dents de l'holotype[1]. Il l'a révisée en 2015 après avoir conclu qu'il avait surestimé le nombre de dents manquantes dans la mandibule[3]. La formule dentaire proposée est S2.A2.I1.LP14 | S1.A2.I1.LP12[N 2] - [3] - [8]. Cela signifie que Cardabiodon possède quatre types de dents, à savoir (d'avant en arrière) : symphysaires, antérieures, intermédiaires et latéropostérieures. La denture de Cardabiodon serait unique parmi les requins. Sur la mandibule, la taille des dents augmenterait brusquement et culminerait à la première dent latéropostérieure, ce qui s'éloignerait de la structure dentaire générique des Lamniformes, où la taille des dents diminue progressivement d'avant vers l'arrière (à l'exception des petites dents symphysaires et intermédiaires)[3]. Les dents inférieures seraient également plus larges que les dents de la mâchoire supérieure, contrairement aux autres lamniformes où c'est l'inverse[8]. L'arc de morsure de la mandibule de l'holotype de C. ricki a été estimé à 74,6 cm[3].
Une autre formule dentaire a été proposée à partir d'un ensemble de dents isolées, identifiées comme Cardabiodon sp. et trouvées dans les schistes de Carlile, au Kansas, à savoir S1?.A2.I1.LP6 | S1?.A2.I1?.LP8. Cette dernière formule se veut plus cohérente avec la structure dentaire générique des lamniformes, tandis que celle de Siverson est mise en doute en raison de ses fortes divergences avec les autres représentants du groupe. Les deux formules restent hypothétiques puisqu'elles se fondent toutes deux sur un ensemble de dents isolées[8].
Classification et évolution
Parmi les requins lamniformes, dont il partage les caractères communs vertébraux et dentaires, Cardabiodon possède une structure dentaire unique. Pour cette raison, Siverson a créé la famille des Cardabiodontidae, où il l'avait placé en compagnie d'un requin fossile du Cénozoïque, Parotodus[1]. Cependant, il est revenu par la suite sur ce choix, notamment en raison du gros écart temporel entre les deux taxons (environ 60 millions d'années)[2]. Un autre requin du Crétacé supérieur, Dwardius, dont les dents ressemblent beaucoup à celles de Cardabiodon, peut être placé chez les Cardabiodontidae[13], à moins qu'il s'agisse du même taxon[10]. Siverson a confirmé l'existence de deux genres distincts en notant des tendances évolutives contraires, telles que la diminution progressive des mini-cuspides latérales chez Cardabiodon au fil du temps, contre l'augmentation progressive de celles-ci chez Dwardius, mais a admis la proximité des deux taxons[11]. La position phylogénétique précise des Cardabiodontidae parmi les Lamniformes reste à établir[2].
Les chercheurs ont découvert qu'une dent de C. ricki, collectée dans des gisements de l'Alberta et datée d'environ 93 millions d'années, présentait des caractéristiques habituellement trouvées chez C. venator, suggérant une possible relation anagénétique entre les deux espèces[10] - [3].
Bien que les Cardabiodontidae ne contiennent à ce jour que les deux genres Cardabiodon et Dwardius[13], plusieurs fossiles non décrits datés de périodes antérieures pourraient être attribués à cette famille. Le plus notable d'entre eux comprend un ensemble de dents d'un même individu et de très grandes vertèbres datées de l'âge Albien, vers 105 millions d'années, provenant de la formation de Toolebuc (en), au Queensland, en Australie, et qui appartiennent à un spécimen dont la longueur est estimée entre 8 et 9 m[14]. Dans une discussion sur un blog, Siverson note que ses dents sont moins robustes que celles d'autres grands requins du Crétacé supérieur, tels que Cretoxyrhina. Il suggère une tendance évolutive des Cardabiodontidae marquée par une réduction de la taille et de la longueur du corps, une réduction du nombre de dents, mais une augmentation de la robustesse des dents (y compris de la racine dentaire) et un élargissement de la couronne, avec une réduction des mini-cuspides latérales[12].
Paléobiologie
Comme tous les lamniformes, Cardabiodon développait chaque année un anneau de croissance dans ses vertèbres, ce qui permet aux paléontologues de calculer l'âge d'un spécimen à sa mort. Cependant, on n'a pu faire que peu de ces mesures en raison de la rareté des vertèbres bien conservées. La croissance de C. ricki a été calculée sur la base des vertèbres de trois individus préadultes et d'une dent isolée de nouveau-né[3]. Par rapport aux données connues sur Cretoxyrhina mantelli et Archaeolamna kopingensis, les vertèbres de C. ricki suggèrent que sa longueur totale à la naissance était inférieure à celle des deux autres requins, entre 41 et 76 cm. Il grandissait à un rythme moyen de 29,9 cm par an, intermédiaire entre le rythme nettement plus rapide de Cretoxyrhina mantelli, de 40,7 cm par an, et le rythme plus lent d'Archaeolamna kopingensis, de 6,2 cm par an[15]. Les trois vertèbres fossiles présentent treize anneaux de croissance, ce qui signifie que les individus ont vécu jusqu'à l'âge de treize ans. Cependant, un taux de croissance plus lent que Cretoxyrhina mantelli suggère une durée de vie potentielle supérieure à treize ans, les trois spécimens étudiés ayant eu une mort prématurée. Autour du cinquième au septième anneau de croissance chez les trois spécimens, on constate un changement de rythme de croissance, suggérant que C. ricki aurait atteint la maturité sexuelle vers l'âge de cinq à sept ans[3].
Paléoécologie
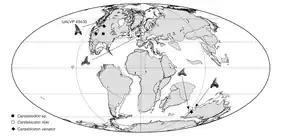
Distribution antitropicale
Des fossiles de Cardabiodon ont été trouvés au Canada, dans le Midwest des États-Unis, en Angleterre, en Australie-Occidentale[3], et au Kazakhstan[1]. On attribue à Cardabiodon une répartition antitropicale, car ces gisements se situaient tous à l'époque entre 40° et 60° de latitude, à l'image de l'aire de répartition du requin-taupe commun actuel, qui ne vit qu'entre 30° et 60° de latitude. Une étude de 2010 confirme que les plages de température dans ces deux aires étaient presque les mêmes. L'étude révèle également que les fossiles de Cardabiodon trouvés les plus hauts en latitude datent de juste avant la période la plus chaude du Crétacé supérieur, qui commence il y a environ 93 millions d'années, phase connue sous le nom d'optimum thermique du Cénomanien-Turonien (en). Ceci suggère une extension de la distribution du requin vers le nord due à la hausse des températures[10].
La température de surface de la mer où vivait Cardabiodon est estimée entre 17,5 et 24,2 °C[10]. Cardabiodon aurait néanmoins pu se déplacer d'un hémisphère à l'autre en empruntant des eaux plus profondes et donc plus froides. On l'observe chez les requins pèlerins actuels, une autre espèce antitropicale, qui migre entre les hémisphères en suivant des chemins plus frais dans les eaux tropicales profondes, où les températures sont similaires à la plage de températures normale du requin pèlerin. Bien qu'aucun fossile de Cardabiodon n'ait été trouvé en dehors de son aire de répartition présumée, cette proposition est étayée par la présence de dents de Cardabiodon dans des dépôts en eau profonde dans les latitudes les plus basses de son aire de répartition[10].
Habitat
Cardabiodon vivait probablement en haute mer, mais dans des eaux modérément profondes, comme les mers intérieures ou la zone néritique. La majorité de ses fossiles connus proviennent d'Amérique du Nord[3], où les fossiles de C. venator ont été trouvés principalement dans la moitié sud de la voie maritime intérieure de l'Ouest, qui avait un climat tempéré un peu plus chaud que la moitié nord[16]. Cardabiodon a traversé l'optimum climatique du Cénomanien-Turonien[10], qui a entraîné une modification de l'environnement et l'apparition et la radiation évolutive de nouvelles formes comme les mosasaures[16].
Les sites du Cénomanien de la voie maritime intérieure de l'Ouest ont livré de nombreux fossiles de vertébrés marins, qu'un superprédateur comme Cardabiodon aurait pu attaquer. Ceux-ci incluent de nombreux requins, notamment des lamniformes comme Cretodus, Cretalamna, Protolamna (en), Cretoxyrhina et Squalicorax, ainsi que des hybodontes comme l'énigmatique Ptychodus et Hybodus, de grands poissons osseux, tels que Protosphyraena, Pachyrhizodus, Enchodus et Xiphactinus, des proto-oiseaux marins comme Pasquiaornis (en) et Ichthyornis, des reptiles marins tels que les élasmosauridés et les plésiosaures polycotylidés, le pliosaure Brachauchenius, les tortues marines protostegidés et des squamates aquatiques tels que Coniasaurus crassidens[16].
Le site de Gearle Siltstone, en Australie-Occidentale, était principalement dominé par Cretalamna, mais d'autres requins tels que Squalicorax, Archaeolamna, Paraisurus (en), Notorhynchus (en), Leptostyrax (en) et Carcharias étaient présents, ainsi que des pliosaures dont a trouvé des restes fragmentaires[17]. Dans d'autres parties de l'Australie-Occidentale vivaient au Cénomanien des ichthyosaures, comme Platypterygius[18].
Les faunes benthiques étaient rares à cette époque, en particulier dans les eaux plus septentrionales habitées par Cardabiodon. Cela était principalement dû à des niveaux d'oxygène plus faibles[16], causés par l'évènement du Cénomanien-Turonien, qui a conduit à l'extinction de pas moins de 27 % des invertébrés marins[19].
Zone de naissance
Comme de nombreux requins actuels, Cardabiodon utilisait des zones d'alevinage pour donner naissance à ses petits, de préférence dans des eaux peu profondes qui offrent une protection naturelle contre les prédateurs. Une zone des schistes de Carlile, dans le Montana, a été identifiée comme site d'alevinage en raison de l'abondance des fossiles de juvéniles de Cardabiodon[2]. D'autres sites de la voie maritime intérieure de l'Ouest, notamment la formation de Kaskapau (en), dans le nord-ouest de l'Alberta, et la formation de Greenhorn, dans le centre du Kansas, ont également livré des fossiles de juvéniles[20]. Ceux-ci étant largement absents ailleurs, cela a conduit à suggérer que la voie maritime intérieure de l'Ouest dans son ensemble aurait pu servir de zone d'alevinage pour Cardabiodon[3].
Notes et références
Notes
- Sur un blog, Siverson affirme que les plus grandes dents de Cardabiodon trouvées sont jusqu'à 15 % plus grandes que celles de l'holotype[12]. Cependant, ces éléments n'ont pas été officiellement publiés dans la littérature scientifique.
- Ceci est basé sur l'interprétation de Dickerson et al. (2012), qui utilise une terminologie plus courante[8].
Références
- (en) Mikael Siverson, « A new large lamniform shark from the uppermost Gearle Siltstone (Cenomanian, Late Cretaceous) of Western Australia », Transactions of the Royal Society of Edinburgh: Earth Sciences, vol. 90, no 1, , p. 49-66 (DOI 10.1017/S0263593300002509, S2CID 131195702)
- (en) Mikael Siverson et Johan Lindgren, « Late Cretaceous sharks Cretoxyrhina and Cardabiodon from Montana, USA », Acta Palaeontologica Polonica, vol. 50, no 2, , p. 301-314 (résumé, lire en ligne [PDF])
- (en) Michael G. Newbrey, Mikael Siverson, Todd D. Cook, Allison M. Fotheringham et Rebecca L. Sanchez, « Vertebral Morphology, Dentition, Age, Growth, and Ecology of the Large Lamniform Shark Cardabiodon ricki », Acta Palaeontologica Polonica, vol. 60, no 4, , p. 877-897 (DOI 10.4202/app.2012.0047
 , S2CID 56258221)
, S2CID 56258221) - (ru) Leonid S. Glickman, « Genetic relations of the Lamnidae and Odontaspidae and new genera of lamnids from the Upper Cretaceous », Trudy Geologicheskogo Muzeia Akademiia Nauk SSSR, vol. 1, , p. 110-117
- (en) Arthur S. Wooward, « Notes on the sharks' teeth from British Cretaceous formations », Proceedings of the Geologists' Association, vol. 13, no 6, , p. 190-200 (DOI 10.1016/S0016-7878(94)80009-4, S2CID 130309655, lire en ligne [PDF])
- Jaques Herman, Les Sélaciens des terrains néocrétacés & paléocènes de Belgique & des contrées limitrophes : éléments d'une biostratigraphie intercontinentale, Bruxelles, Service géologique de Belgique, , 401 p. (OCLC 612294375)
- (en) Mikael Siverson, « Lamniform sharks of the mid Cretaceous Alinga Formation and Beedagong Claystone, Western Australia », Palaeontology, vol. 39, no 4, , p. 813-849 (résumé, lire en ligne [PDF])
- (en) Ashley A. Dickerson, Kenshu Shimada, Brian Reilly et Cynthia R. Rigsby, « New data on the Late Cretaceous cardabiodontid lamniform shark based on an associated specimen from Kansas », Transactions of the Kansas Academy of Science, vol. 115, nos 3-4, , p. 125-134 (DOI 10.1660/062.115.0305, S2CID 83864321)
- (en) Alexander D. Meglei, Kenshu Shimada et James I. Kirkland, « Fossil Vertebrates from the Middle Graneros Shale (Upper Cretaceous: Middle Cenomanian) in Southeastern Nebraska », Transactions of the Kansas Academy of Science, vol. 116, nos 3-4, , p. 129-136 (DOI 10.1660/062.116.0304, S2CID 83671316)
- (en) Todd D. Cook, Mark V. H. Wilson et Michael G. Newbrey, « The first record of the large Cretaceous lamniform shark Cardabiodon ricki from North America and a new empirical test for its presumed antitropical distribution », Journal of Vertebrate Paleontology, vol. 30, no 3, , p. 643-649 (DOI 10.1080/02724631003758052, JSTOR 40666186, S2CID 128489655)
- (en) Mikael Siverson, David J. Ward, Johan Lindgren et L. Scott Kelley, « Mid-Cretaceous Cretoxyrhina (Elasmobranchii) from Mangyshlak, Kazakhstan and Texas, USA », Alcheringa: An Australasian Journal of Palaeontology, vol. 37, no 1, , p. 87-104 (DOI 10.1080/03115518.2012.709440, S2CID 129785545)
- (en) Mikael Siverson, « Cardabiodon size, HELP! », sur The Fossil Forum, (consulté le )
- (en) Mikael Siverson et Marcin Machalski, « Late late Albian (Early Cretaceous) shark teeth from Annopol, Poland », Alcheringa: An Australasian Journal of Palaeontology, vol. 41, no 4, , p. 433-463 (DOI 10.1080/03115518.2017.1282981, S2CID 133123002)
- (en) Lamniform Sharks: 110 Million Years of Ocean Supremacy, Mikael Siverson () Royal Tyrrell Museum of Palaeontology.
- (en) Joseph A. Fredrickson, Joshua E. Cohen et Jeff L. Berry, « Ontogeny and life history of a large lamniform shark from the Early Cretaceous of North America », Cretaceous Research, vol. 59, no 1, , p. 272-277 (DOI 10.1016/j.cretres.2015.11.007, S2CID 131249276)
- (en) Stephen L. Cumbaa, Kenshu Shimada et Todd D. Cook, « Mid-Cenomanian vertebrate faunas of the Western Interior Seaway of North America and their evolutionary, paleobiogeographical, and paleoecological implications », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 295, nos 1-2, , p. 199-214 (DOI 10.1016/j.palaeo.2010.05.038, Bibcode 2010PPP...295..199C, S2CID 129427657)
- (en) Mikael Siverson, « Sharks from the mid-Cretaceous Gearle Siltstone, Southern Carnarvon Basin, Western Australia », Journal of Vertebrate Paleontology, vol. 17, no 3, , p. 453-465 (DOI 10.1080/02724634.1997.10010995, JSTOR 4523829, S2CID 129572662)
- (en) Benjamin Kear, « Cretaceous marine reptiles of Australia: a review of taxonomy and distribution », Cretaceous Research, vol. 24, no 2003, , p. 277-303 (DOI 10.1016/S0195-6671(03)00046-6, S2CID 128619215, lire en ligne [PDF])
- (en) In Brief, « Submarine eruption bled Earth's oceans of oxygen » [archive du ], New Scientist, (consulté le )
- (en) Todd D. Cook, Mark V.H. Wilson, Alison M. Murray, A. Guy Plint, Michael G. Newbrey et Michael J. Everhart, « A high latitude euselachian assemblage from the early Turonian of Alberta, Canada », Journal of Systematic Palaeontology, vol. 11, no 5, , p. 555-587 (DOI 10.1080/14772019.2012.707990, S2CID 129551937, lire en ligne)
Voir aussi
Articles connexes
Liens externes
- (en) Mikael Siverson, « The rise of super predatory sharks » , sur Western Australian Museum
- Ressources relatives au vivant :