Platecarpus
Platecarpus tympaniticus

- Platecarpus coryphaeus
- Platecarpus ictericus
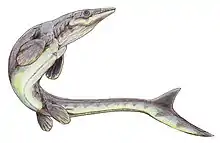
Platecarpus est un genre éteint et fossile des mosasaures ayant vécu durant le Crétacé supérieur (âges Santonien-Campanien), il y a entre 84 et 81 millions d'années avant notre ère, dans ce qui actuellement l'Amérique du Nord, voire peut-être même l'Afrique et la Belgique. Une seule espèce est actuellement reconnue, Platecarpus tympaniticus, décrite par Edward Drinker Cope en 1869, les autres espèces ayant été déclarés comme synonymes ou appartenant à un genre différent. C'est un reptile marin d'une taille mesurant environ 5,5 mètres de long, avec un régime alimentaire s'avérant être piscivore.
Comme d'autres mosasaures, les chercheurs pensait initialement que Platecarpus nageait à la manière d'une anguille, tandis que d'autres suggéraient qu'il nageait davantage comme les requins modernes. Un spécimen exceptionnellement bien conservé de P. tympaniticus montre les traces d'empreintes cutanées, des pigments autour des narines, des bronches et la présence d'une ailette de queue très profilée, montrant que ce genre ainsi que d'autres mosasaures n'avaient pas nécessairement une méthode de nage similaire aux anguilles, mais étaient des nageurs plus puissants et plus rapides.
Découverte et fossiles
Divers squelettes de Platecarpus ont été trouvés dans des gisements du Crétacé, et plus précisément dans des roches datant de la fin du Santonien jusqu'au début du Campanien, dans le membre de Smoky Hill Chalk (en), situé au Kansas, aux États-Unis[2]. Des fossiles supplémentaires attribuées à Platecarpus furent découverts en Afrique et en Belgique, mais leur affiliation au genre sont encore non confirmée et nécessite une réévaluation[3].
Description


Platecarpus possède une longue queue tournée vers le bas avec une grande ailette dessus, des palette natatoires directrices et des mâchoires bordées de dents coniques. L'animal devrait atteindre une taille de 5,4 à 5,67 m de long et devrait peser 440 kg[4] - [5].
La structure crânienne de Platecarpus est unique parmi les mosasaures. Ce genre se caractérise par un crâne court et a moins de dents que tout autres mosasauridés (environ 10 dents dans chaque os dentaires)[N 1]. Le spécimen LACM 128319 préserve la matière dans l'anneau sclérotique qui pourrait éventuellement être la rétine de l'œil. De petites structures dans la rétine, chacune d'environ 2 µm de long et observées par microscopie électronique à balayage, peuvent représenter des mélanosomes rétiniens conservés dans leurs positions d'origine[4].
La trachée est également connu dans LACM 128319, conservé sous forme d'anneaux trachéaux cartilagineux. Seule l'extrémité la plus postérieure du tube trachéal, situé à l'extrémité du cou près de la ceinture pectorale, est connue. La section où les deux bronches sont séparées a également été conservée dans le spécimen, mais a été détruite lors de la fouille. Ceci indique que Platecarpus et d'autres mosasaures avaient deux poumons fonctionnels. Les serpents, qui sont étroitement apparentés aux mosasaures, n'ont qu'un seul poumon fonctionnel, le second étant souvent vestigial ou absent. Contrairement aux lézards terrestres, cependant, les bronches se séparent devant la zone des membres antérieurs plutôt qu'au niveau des membres[4].
Les empreintes cutanées sont connues de Platecarpus, conservées dans LACM 128319 sous forme d'empreintes molles et de matériau phosphaté. Les écailles sur le bout du museau et le sommet du crâne sont de forme quelque peu hexagonale et ne se touchent pas. Les écailles des mâchoires sont plus longues et de forme rhomboïdale, se chevauchant. Les écailles sur le museau indiquent que les narines étaient placées loin devant le crâne à son extrémité et tournées latéralement comme chez la plupart des squamates et archosaures. Les écailles du corps sont toutes de forme rhomboïdale et forment des rangées diagonales étroitement reliées qui se chevauchent sur leurs bords postérieurs. Ils sont généralement de la même taille sur toute la longueur du corps. Les écailles caudales sont plus hautes et plus grandes que celles du reste du corps, bien que celles qui recouvrent la surface inférieure de la queue ressemblent davantage aux écailles corporelles[4].
Les organes internes, peuvent également être conservés dans le spécimen sous forme de zones rougeâtres. L'un est situé dans la cavité thoracique en bas de la cage thoracique, tandis que l'autre est situé dans la partie supérieure de la cavité abdominale juste derrière la cage thoracique. Les zones rougeâtres ont été analysées par spectrométrie de masse et se sont révélées contenir des niveaux élevés de composés à base de fer et de porphyrine. Ces substances sont la preuve de produits de décomposition de l'hémoglobine qui peuvent s'être formés dans les organes lors de leur décomposition. Selon sa position, l'organe de la cavité thoracique est probablement le cœur ou le foie, voire les deux. L'organe de la cavité abdominale peut être un rein, bien qu'il soit dans une position plus antérieure que les reins des varans, les parents vivants parmi les plus proches des mosasaures. La position antérieure des reins peut avoir été une adaptation vers un corps plus profilé, car leur position présumée est similaire à celle des cétacés[4].
Une partie du tube digestif est également préservée et est remplie de restes de poissons de taille moyenne. La forme de ces restes peut décrire la véritable forme de la partie correspondante du tube digestif, très probablement le côlon. La présence d'écailles et d'os non digérés dans le côlon suggère que Platecarpus et d'autres mosasaures ont traité les aliments rapidement et n'ont pas complètement digéré et absorbé tous les aliments dans le tractus gastro-intestinal. Les coprolithes du mosasaure Globidens suggèrent également de faibles taux de digestion et d'absorption car ils contiennent des masses de coquilles de bivalves broyées[4].
Les vertèbres caudales sont fortement inclinées vers le bas. Les vertèbres du pédoncule caudal sont en forme de coin avec des épines neurales plus larges à leurs extrémités qu'elles ne le sont à leurs bases. Cette zone retournée aurait probablement soutenu un ailette similaire aux requins modernes. Deux lobes auraient été présents, un inférieur soutenu par les vertèbres renversées et un supérieur non soutenu. L'ailette de la queue était probablement hypocercale, ce qui signifie que son lobe inférieur est plus long que son lobe supérieur. Cette condition est également observée chez les ichtyosaures et les crocodyliformes du groupe des métriorhynchidés[4].
Classification
Platecarpus était souvent considéré comme le genre le plus courant de mosasaure du dépôt du Smoky Hill Chalk (en) au Kansas, et Platecarpus ictericus était considéré comme l'espèce la plus représentatif. Cependant, les scientifiques le considèrent maintenant comme étant du genre paraphylétique. Par conséquent, certaines espèces ont été réaffectées à leurs propres genres. Le spécimen type de Platecarpus planiforns a été découvert par le professeur B. F. Mudge et a été classé par Edward Drinker Cope comme Clidastes planiforns[2]. En 1898, après une analyse plus approfondie des restes, il est déterminé que ce mosasaure devait être placé dans le genre Platecarpus[6]. Le spécimen holotype subi un autre examen taxonomique en 1967, lorsque le paléontologue Dale Russell détermine que les restes sont trop fragmentaires pour être placés dans n'importe quel genre, et le considère comme un spécimen incertae sedis[7]. En 2011, un nouveau nom générique, Plesioplatecarpus (en), a été érigé par Takuya Konishi et Michael W. Caldwell pour incorporer P. planifrons, qu'ils considère comme distinct de Platecarpus dans une analyse phylogénétique[8]. En 1994, Angolasaurus fut synonymisé avec ce genre[9]. Cependant, de nombreuses études récentes revalident ce genre. De plus, Platecarpus est considéré comme un taxon monotypique, car P. coryphaeus et P. ictericus sont considérés synonymes de l'espèce type, P. tympaniticus[8] - [10].
Le cladogramme ci-dessous suit la topologie la plus résolue d'une analyse de 2011 par Konishi et Caldwell[8] :
| ◄ |
| ||||||||||||
Paléobiologie
Régime alimentaire
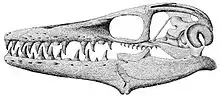
Comparés aux tylosaurinés, les plioplatecarpinés (dont fait partie Platecarpus) ont des dents beaucoup moins robustes, ce qui suggère qu'ils se nourrissaient de proies plus petites et/ou plus molles comme les petits poissons et les calmars[2].
Locomotion
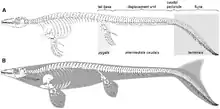
Alors que les chercheurs pensent traditionnellement que les mosasaures se sont propulsés dans l'eau par ondulation latérale de la même manière que les anguilles, la nageoire caudale profonde de Platecarpus suggère qu'il nageait plus comme un requin. Les vertèbres caudales tournées vers le bas de Platecarpus suggèrent qu'il avait une queue en forme de croissant. Au point de la queue où l'ailette se situe, les centra vertébraux sont raccourcis et ressemblent à des disques. Leur taille réduite aurait probablement permis une plus grande flexibilité dans une zone qui aurait subi des contraintes élevées pendant la nage. Les épines neurales de ces vertèbres ont également des rainures pour l'insertion des ligaments inter-épineux et des tissus conjonctifs dorsaux qui auraient aidé au mouvement latéral de la douve. Les ligaments seraient probablement constitués de fibres de collagène qui agissaient comme des ressorts pour ramener la queue dans une position de repos après que l'énergie y ait été stockée. Ces types de ligaments fonctionnent chez certains poissons vivants pour conserver l'énergie lors de la flexion répétitive de la queue. Alors que la nageoire et l'arrière de la queue ondulaient chez Platecarpus, la base de la queue restait stable. Cette forme de mouvement est connue sous le nom de locomotion carangiforme[4].
La structure des écailles de Platecarpus peut avoir été une autre adaptation à un mode de vie marin. La petite taille et la forme similaire de ces écailles sur tout le corps auraient rigidifié le tronc, le rendant plus résistant aux mouvements latéraux. Cette rigidité aurait amélioré l'efficacité hydrodynamique en améliorant l'écoulement de l'eau à travers le corps. Le premier mosasauroïde Vallecillosaurus (en) conserve également des écailles corporelles, mais elles sont plus grandes et de forme plus variée, ce qui suggère que l'animal s'appuyait sur le mouvement ondulatoire de son tronc plutôt que sur sa queue. Plotosaurus, un mosasauridé plus dérivé que Platecarpus, a des écailles encore plus petites couvrant son corps, indiquant qu'il avait une locomotion encore plus efficace dans l'eau[4].
Paléoécologie
L'analyse isotopique sur des spécimens de dents suggère que Platecarpus et Clidastes peuvent être entrés occasionnellement en eau douce, tout comme les serpents marins actuels[11].
Notes et références
Notes
- Une espèce non classée de Plioplatecarpus provenant de la formation de Demopolis Chalk en Alabama montre un nombre inférieur de dents dans ses mâchoires[4].
Références
- (en) E. D. Cope, « On the reptilian orders Pythonomorpha and Streptosauria », Proceedings of the Boston Society of Natural History, vol. 12, , p. 250-266
- (en) Michael J. Everhart, Oceans of Kansas: A Natural History of the Western Interior Seaway, Bloomington, Indiana University Press, , 165-169 p.
- (en) D. Palmer, The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals, Londre, Marshall Editions, (ISBN 978-1-84028-152-1), p. 87
- (en) Johan Lindgren, Michael W. Caldwell, Takuya Konishi et Luis M. Chiappe, « Convergent Evolution in Aquatic Tetrapods: Insights from an Exceptional Fossil Mosasaur », PLOS ONE, vol. 5, no 8, , e11998 (PMID 20711249, PMCID 2918493, DOI 10.1371/journal.pone.0011998
 , Bibcode 2010PLoSO...511998L)
, Bibcode 2010PLoSO...511998L) - (en) Gregory S. Paul, The Princeton Field Guide to Mesozoic Sea Reptiles, Princeton University Press, , 170 p. (ISBN 978-0-69-119380-9, DOI 10.2307/j.ctv2hnkc6h, S2CID 251553177)
- (en) M. J. Everhart, « Linking the Past with the Present: The Mosasaurs of Samuel W. Williston » [archive du ], sur Oceans of Kansas,
- (en) Dale A. Russell, Systematics and morphology of American mosasaurs, vol. 23, New Haven, Bulletin of the Peabody Museum of Natural History, , 240 p. (OCLC 205385, lire en ligne)
- (en) Takuya Konishi et Michael W. Caldwell, « Two new plioplatecarpine (Squamata, Mosasauridae) genera from the Upper Cretaceous of North America, and a global phylogenetic analysis of plioplatecarpines », Journal of Vertebrate Paleontology, vol. 31, no 4, , p. 754-783 (DOI 10.1080/02724634.2011.579023, JSTOR 25835874, S2CID 85972311)
- (de + en) Theagarten Lingham-Soliar, « The mosasaur “Angolasaurus” bocagei (Reptilia: Mosasauridae) from the Turonian of Angola re-interpreted as the earliest member of the genus Platecarpus », Paläontologische Zeitschrift, vol. 68, nos 1-2, , p. 267-282 (DOI 10.1007/BF02989445, S2CID 128963124)
- (en) Caitlin R. Kiernan, « Stratigraphic distribution and habitat segregation of mosasaurs in the Upper Cretaceous of western and central Alabama, with an historical review of alabama mosasaur discoveries », Journal of Vertebrate Paleontology, vol. 22, no 1, , p. 91-103 (DOI 10.1671/0272-4634(2002)022[0091:SDAHSO]2.0.CO;2, S2CID 130280406, lire en ligne)
- (en) L. T. Taylor, R. T. Minzoni, C. A. Suarez, L. A. Gonzalez, L. D. Martin, W. J. Lambert, D. J. Ehret et T. L. Harrell, « Oxygen isotopes from the teeth of Cretaceous marine lizards reveal their migration and consumption of freshwater in the Western Interior Seaway, North America », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 573, , p. 110406 (DOI 10.1016/j.palaeo.2021.110406, Bibcode 2021PPP...573k0406T, S2CID 234799487)
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :