Velociraptor
Vélociraptor

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Classe | Sauropsida |
| Super-ordre | Dinosauria |
| Ordre | Saurischia |
| Sous-ordre | Theropoda |
| Famille | † Dromaeosauridae |
| Sous-famille | † Velociraptorinae |
Espèces de rang inférieur
Velociraptor (littéralement « voleur rapide »), souvent francisé sous le terme vélociraptor ou « raptor », est un genre éteint de petits dinosaures théropodes appartenant à la famille des droméosauridés, ayant vécu durant le Crétacé supérieur dans ce qui est aujourd'hui l'Asie, il y a entre 75 et 71 millions d'années. Deux espèces sont actuellement connues, bien que d'autres aient été attribuées par le passé. L'espèce type, V. mongoliensis, est connue à partir des fossiles découverts dans la formation de Djadokhta en Mongolie. Une deuxième espèce, V. osmolskae, a été décrite en 2008 grâce à des fragments crâniens découverts dans la formation de Bayan Mandahu en Chine.
Mesurant environ 1,5 à 2 m de long pour une masse corporelle comprise entre 15 et 18 kg, Velociraptor est plus petit que d'autres droméosauridés tels que Deinonychus ou Achillobator. Il partage néanmoins bon nombre de caractéristiques anatomiques avec ceux-ci. C'était un carnivore bipède à plumes doté d'une longue queue et d'une grande griffe en forme de faucille sur chaque patte arrière, qui aurait été utilisée pour attaquer et retenir les proies. Velociraptor se distingue des autres droméosauridés par son crâne long et bas, au museau retroussé.
Velociraptor est l'un des dinosaures les plus connus du grand public en raison de son rôle de premier plan dans les films de la saga Jurassic Park. Cependant, Velociraptor est à peu près haut comme une dinde, considérablement plus petit que les reptiles d'environ 2 m de haut et de 90 kg vus dans les romans et les films (qui sont basés sur des membres du genre apparenté Deinonychus). Aujourd'hui, de tous les droméosauridés, Velociraptor est bien connu des paléontologues, plus d'une douzaine de squelettes fossiles ayant été décrits. Un fossile particulièrement célèbre conserve d'ailleurs un Velociraptor figé dans un combat avec un Protoceratops.
Historique des découvertes
Premières descriptions
.jpg.webp)
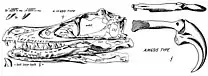
Le , au cours d'une expédition menée aux Flaming Cliffs, situés dans la formation de Djadokhta dans le désert de Gobi, en Mongolie, pour le compte du musée américain d'histoire naturelle, Peter Kaisen découvre le premier fossile de Velociraptor connu des chercheurs. Il s'agit d'un crâne écrasé mais complet, associé à l'une des griffes du deuxième orteil, catalogué AMNH 6515. En 1924, le président du musée, Henry Fairfield Osborn, désigne le crâne et la griffe, qu'il suppose provenir de la main, comme le spécimen holotype de son nouveau genre, qu'il nomme Velociraptor. Le nom générique est dérivé des mots latins velox, « rapide », et raptor, « voleur » ou « pilleur », et fait référence au côté coureur et au régime carnivore de l'animal. Osborn nomme l'espèce type V. mongoliensis en référence au pays d'origine de la découverte[1]. Plus tôt cette année-là, Osborn a déjà mentionné de manière informelle l'animal dans un article de presse populaire, sous le nom « Ovoraptor djadochtari » (distinct de Oviraptor du même nom)[2], pour finalement le changer en V. mongoliensis lors de sa description formelle[1].
Alors que les équipes nord-américaines ont été exclues de la Mongolie communiste pendant la guerre froide, des expéditions de scientifiques soviétiques et polonais, en collaboration avec des collègues mongols, ont récupéré plusieurs autres spécimens de Velociraptor. Le plus célèbre est Fighting Dinosaurs (catalogué MPC-D 100/25, anciennement IGM, GIN ou GI SPS), découvert par une équipe polono-mongole en 1971. Le fossile préserve un Velociraptor en plein combat contre un Protoceratops[3] - [4] - [5]. Il est considéré comme un trésor national de la Mongolie et, en 2000, il a été prêté au musée américain d'histoire naturelle de New York pour une exposition temporaire[6].
Entre 1988 et 1990, une équipe sino-canadienne découvre des restes de Velociraptor dans le nord de la Chine[7]. Des scientifiques américains sont retournés en Mongolie en 1990 à l’occasion d’une expédition mongolo-américaine conjointe dans le désert de Gobi, dirigée par le musée américain d'histoire naturelle et par l'académie des sciences de Mongolie, ce qui a permis de découvrir plusieurs squelettes bien conservés[8] - [9]. Un de ces spécimens, MPC-D 100/980, a été surnommé « Ichabodcraniosaurus » par l'équipe de Mark Norell parce qu’un spécimen assez complet avait été trouvé sans son crâne, une allusion au personnage d'Ichabod Crane, héros de la nouvelle La Légende de Sleepy Hollow de Washington Irving[10]. Alors que Norell et Makovicky le considéraient provisoirement comme un spécimen de V. mongoliensis[8], il a finalement été décrit en 2021 comme étant une nouvelle espèce se nommant Shri devi[11].
Espèces additionnelles
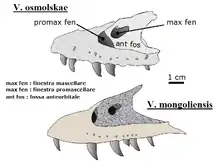
Un os maxillaire et lacrymal, découvert dans la formation de Bayan Mandahu en 1999 par les expéditions sino-belges, s'est avéré appartenir à Velociraptor, mais pas à l'espèce type V. mongoliensis. Pascal Godefroit et ses collègues nomment en 2008 le propriétaire de cet os V. osmolskae, en hommage à la paléontologue polonaise Halszka Osmólska[12]. Cependant, l'étude de 2013 a noté que si « la forme allongée du maxillaire de V. osmolskae est similaire à celle de V. mongoliensis », l'analyse phylogénétique révèle qu'il est plus proche de Linheraptor, ce qui fait de ce dernier un genre paraphylétique. Ainsi, V. osmolskae pourrait ne pas appartenir au genre Velociraptor et nécessite une réévaluation[13].
Les paléontologues Mark A. Norell et Peter J. Makovicky décrivent en 1997 de nouveaux spécimens remarquablement préservés de V. mongoliensis, à savoir MPC-D 100/985, collecté dans la localité de Tugrik Shireh, et MPC-D 100/986, collecté dans la localité de Chimney Buttes, tous deux découverts en 1993. L'équipe mentionne brièvement un autre spécimen, MPC-D 100/982, qui au moment de cette publication restait non décrit[9]. En 1999, Norell et Makovicky fournissent plus d'informations sur l'anatomie de Velociraptor avec des spécimens supplémentaires. Parmi ceux-ci, MPC-D 100/982 est partiellement décrit et compris, et référencé à V. mongoliensis, principalement sur la base de similitudes crâniennes avec le crâne holotype, bien qu'ils déclarent que des différences seraient présentes entre la région pelvienne de ce spécimen et sur d'autres spécimens de Velociraptor. Ce spécimen relativement bien conservé comprenant le crâne, a été découvert et collecté en 1995 dans la localité de Bayn Dzak (plus précisément dans la sous-localité « Volcano »)[8]. Martin Kundrát, dans un résumé de 2004, compare le neurocrâne de MPC-D 100/982 à un autre spécimen de Velociraptor, MPC-D 100/976. Il en conclu que la morphologie globale du premier est plus dérivée que le second, suggérant qu'ils pourraient représenter des taxons distincts[14].

Dans une thèse de maîtrise publiée en 2020, Mark J. Powers décrit en détail MPC-D 100/982, dont il en conclu qu'il représenterait une nouvelle et troisième espèce de Velociraptor. Cette espèce, qu'il nomme V. vadarostrum, diffère principalement des autres espèces de Velociraptor par sa morphologie du maxillaire peu profonde[15]. Powers et ses collègues utilise également en 2020 des analyses morphométriques pour comparer plusieurs maxillaires de droméosauridés et constatent que le maxillaire de MPC-D 100/982 diffère fortement des spécimens référés à Velociraptor. Ils indiquent que ce spécimen, sur la base de ces résultats, représente une espèce différente[16]. En 2021, Powers et son équipe utilisent l'analyse en composantes principales pour séparer les maxillaires des droméosauridés, découvrant notamment que MPC-D 100/982 se situe en dehors de la variabilité intraspécifique de V. mongoliensis, plaidant en faveur d’une espèce distincte. Ils considèrent que V. mongoliensis et cette nouvelle espèce seraient écologiquement séparés en fonction de l'anatomie de leur crâne[17]. L'équipe, dans un autre article publié la même année, renforcent de nouveau la séparation au niveau des espèces, notant que des différences supplémentaires peuvent être trouvées dans les membres postérieurs[18].
Description
Aspect général

Velociraptor est un droméosauridé de taille petite à moyenne, les adultes mesurant entre 1,5 et 2,07 m de long pour environ 50 cm de haut au niveau des hanches[19] - [20], le tout pour un poids estimé entre 15 et 18,3 kg[21].
Des boutons de piquant proéminents, servant de fixation des plumes « d'aile » et indicateur direct d'un revêtement de plumes, ont été signalés sur l'ulna d'un seul spécimen de Velociraptor (IGM 100/981), qui représente un animal d'environ 1,5 m de long et de 15 kg en poids. L'espacement des six boutons conservés suggère que 8 boutons supplémentaires peuvent avoir été présents, donnant un total de 14 boutons de plume qui ont développé de grandes pennes[22]. Cependant, le numéro de spécimen a été corrigé en IGM 100/3503 et son renvoi à Velociraptor peut nécessiter une réévaluation, en attendant une étude plus approfondie[23]. Néanmoins, il existe de solides preuves phylogénétiques d'autres parents de droméosauridés qui indiquent la présence de plumes chez Velociraptor, y compris des droméosauridés tels que Microraptor ou Zhenyuanlong[24] - [25].
Crâne
.png.webp)
.png.webp)
.png.webp)
Le crâne de Velociraptor est plutôt allongé et atteint 23 cm de long. Il est uniquement courbé vers le haut dans la région du museau, concave sur la surface supérieure et convexe sur la surface inférieure. Le museau, qui occupe environ 60 % de toute la longueur du crâne, est particulièrement étroit et principalement formé par les os nasaux, prémaxillaires et maxillaires. Le prémaxillaire est l'os le plus antérieur du crâne, et il est plus long que haut. Alors que son extrémité postérieure rejoint le nasal, le corps principal du prémaxillaire touche le maxillaire. Le maxillaire est de forme presque triangulaire et est le plus grand élément du museau. Sur son centre, ou corps principal, il y a une dépression développant un petit trou de forme ovale à circulaire, appelé fenêtre maxillaire. En face de cette fenêtre se trouvent deux petites ouvertures, appelées fenêtres promaxillaires. Le bord postérieur du maxillaire forme principalement la fenêtre antéorbitaire, l'un des nombreux grands trous du crâne. Le prémaxillaire et le maxillaire ont plusieurs alvéoles dentaires sur leurs surfaces inférieures. Au-dessus du maxillaire et en contact avec le prémaxillaire, il y a l'os nasal. C'est un os étroit et allongé contribuant à la surface supérieure du museau. Ensemble, les os prémaxillaires et nasaux donnent forme à l'ouverture de la narine, qui est relativement grande et circulaire. L'extrémité postérieure de l'os nasal est rejointe par les os frontal et lacrymal[26] - [27].
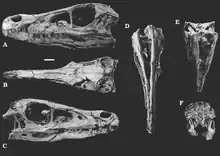
La région postérieure ou antérieure du crâne est constituée des os frontaux, lacrymaux, postorbitaires, jugal, pariétal, bréchet et du quadratojugal. Le frontal est un élément de grande taille, ayant une forme vaguement rectangulaire vu d'en haut. À son extrémité postérieure, cet os est en contact avec le pariétal, et ces éléments constituent les corps principaux du toit crânien. Le lacrymal est un os en forme de « T » et son corps principal est mince et délicat. Son extrémité inférieure rencontre le jugal (souvent appelé pommette), qui est un grand élément de forme sous-triangulaire. Sa bordure inférieure est notamment droite et horizontale. Le postorbitaire est situé juste au-dessus du jugal, un os trapu et fortement en forme de « T ». Dans son ensemble, l'orbite oculaire, formée par le lacrymal, le jugal, le frontal et le postorbital, est grande et de forme presque circulaire, étant plus longue que haute. Vu d'en haut, une paire de grands trous nettement arrondis sont présents près de l'arrière du crâne (les fenêtres temporales), dont les principaux composants sont le postorbitaires et le squamosals. Derrière le jugal, un os en forme de « T » inversé (également observé chez d'autres droméosauridés), connu sous le nom de quadratojugal, s'est développé. Alors que l'extrémité supérieure du quadratojugal rejoint le squamosal (un élément de forme irrégulière), sa face interne rencontre le quadratojugal. Ce dernier est d'une grande importance pour l'articulation avec la mâchoire inférieure. L'os le plus postérieur est l'os occipital et se projette sur le condyle occipital, une protubérance arrondie et bulbeuse qui rencontre la première vertèbre du cou[26] - [27].
La mâchoire inférieure de Velociraptor comprend principalement les os dentaires, spléniaux, angulaires, surangulaires et articulaire. Le dentaire, l'os principal de la mandibule, est un élément très long, faiblement incurvé et étroit qui développe plusieurs alvéoles sur sa surface supérieure. À son extrémité postérieure, il rencontre le surangulaire. Il a un petit trou près de son extrémité postérieure, appelé foramen surangulaire. Les deux os sont les plus gros éléments de la mâchoire inférieure de Velociraptor, contribuant à pratiquement toute sa longueur. Au-dessous d'eux se trouvent les plus petits os spléniaux et anguleux, étroitement articulés les uns aux autres. L'articulaire, l'os le plus en arrière de la mâchoire inférieure, située sur la face interne du surangulaire, est un petit élément qui rejoint le quadratojugal du crâne, permettant ainsi l'articulation avec la mâchoire inférieure. Un trou allongé, presque de forme ovale, s'y développe au centre de la mâchoire inférieure (fenêtre mandibulaire), qui est produit par l'articulation des os dentaires, surangulaires et angulaires[26] - [27].
Les dents de Velociraptor sont assez similaires les uns des autres et ont plusieurs denticules, chacune plus fortement dentelée sur le bord arrière que sur le devant. Le prémaxillaire a quatre alvéoles (ce qui signifie que quatre dents ont été développées) et le maxillaire en a onze. Au dentaire, entre 14 et 15 alvéoles sont présents. Toutes les dents présentes au prémaxillaire sont mal courbées et les deux premières dents sont les plus longues, la seconde ayant une grande taille caractéristique. Les dents du maxillaires sont plus fines, recourbées et, plus particulièrement, l'extrémité inférieure est fortement plus crénelée que la supérieure[26] - [27].
Squelette postcrânien
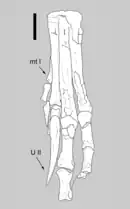
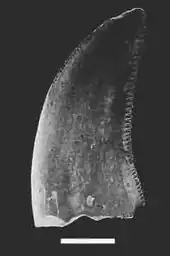
Le bras de Velociraptor est formé par l'humérus, le radius, le cubitus et le manus. Velociraptor, comme les autres droméosauridés, possède une grand main à trois doigts allongés, qui se terminent par des unguals fortement incurvés (os de griffes) qui sont similaires en construction et en flexibilité aux os des ailes des oiseaux actuels. Le deuxième doigt est le plus long des trois doigts présents, tandis que le premier est le plus court. La structure des os du carpe empêche la pronation du poignet et oblige le manus à être tenu avec la surface palmaire tournée vers l'intérieur (médialement) et non vers le bas. L'anatomie de la patte de Velociraptor se compose du métatarse, un grand élément composé de trois os métatarsiens dont le premier est de taille extrêmement réduite, et de quatre doigts qui développent de grands unguals. Le premier doigt, comme chez les autres théropodes, est un petit ergot. Le deuxième doigt, pour lequel a fait la popularité de Velociraptor, est fortement modifié et devrait être maintenu rétracté du sol, ce qui a amené Velociraptor et d'autres droméosauridés à ne marcher que sur leurs troisième et quatrième doigt. Il porte une griffe relativement grande en forme de faucille, typique des droméosauridés et des troodontidés. Cette griffe élargie, qui peut atteindre plus de 6,5 cm de long autour de son bord extérieur, était très probablement un dispositif de prédation utilisé pour retenir les proies en difficulté[9] - [8].
Comme chez les autres droméosauridés, la queue de Velociraptor a des longues projections osseuses appelée prézygapophyses sur les surfaces supérieures des vertèbres, ainsi que des tendons ossifiés en dessous. Les prézygapophyses commencent sur la dixième vertèbre caudale et se s'étendent vers l'avant pour renforcer quatre à dix vertèbres supplémentaires, selon la position dans la queue. Les paléontologues pensaient autrefois que ceux-ci raidissaient complètement la queue, la forçant à agir comme une seule unité en forme de tige. Cependant, au moins un spécimen conserve une série de vertèbres caudales intactes courbées latéralement en forme de « S », ce qui suggère qu'il y avait considérablement plus de flexibilité horizontale que ce qui était généralement admis[9] - [8] - [28].
Classification
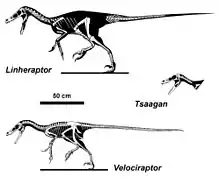
Velociraptor fait partie du clade des Eudromaeosauria, un sous-groupe dérivé de la grande famille des Dromaeosauridae. Il est souvent placé dans la sous-famille, érigé en son nom, des Velociraptorinae. Dans la taxonomie phylogénétique, les Velociraptorinae sont généralement défini comme « tous les droméosaures plus étroitement liés à Velociraptor qu'à Dromaeosaurus ». Cependant, la classification des droméosauridés est très variable. À l'origine, la sous-famille Velociraptorinae a été érigée uniquement pour contenir Velociraptor[3]. D'autres analyses ont souvent inclus d'autres genres, généralement Deinonychus et Saurornitholestes[29], et plus récemment Tsaagan[30]. Plusieurs études publiées au cours des années 2010, y compris des versions étendues des analyses qui ont trouvé un soutien pour les Velociraptorinae, n'ont pas réussi à le résoudre en tant que groupe distinct, mais ont plutôt suggéré qu'il s'agit d'un grade paraphylétique qui aurait donné naissance aux Dromaeosaurinae[31] - [32].
Lorsqu'il a été décrit pour la première fois en 1924, Velociraptor a été placé dans la famille des Megalosauridae, comme c'était le cas avec la plupart des dinosaures carnivores à l'époque[N 1]. Alors que les découvertes de dinosaures se multipliaient, Velociraptor fut plus tard reconnu comme un droméosauridé. Tous les droméosauridés ont également été référés à la famille des Archaeopterygidae par au moins un auteur, ce qui en ferait de Velociraptor un oiseau incapable de voler[33]. D'autres espèces de droméosauridés, dont Deinonychus antirrhopus et Saurornitholestes langstoni, ont parfois été classées dans le genre Velociraptor ; ces espèces étaient alors nommées V. antirrhopus et V. langstoni[19]. Depuis 2008, les seules espèces de Velociraptor actuellement reconnues sont V. mongoliensis[27] - [33] - [34] et V. osmolskae[12]. Cependant, plusieurs études considèrent que V. osmolskae serait apparenté de loin à V. mongoliensis[35] - [36].

Ci-dessous, les résultats de la phylogénie des Eudromaeosauria basés d'après l'analyse phylogénétique menée par James G. Napoli et son équipe en 2021[23] :
Paléobiologie
Plumes

En 2007, Alan H. Turner et ses collègues signalent la présence de six boutons de plume dans l'ulna d'un spécimen référé à Velociraptor (IGM 100/981) de la localité d'Ukhaa Tolgod de la formation de Djadokhta. Turner et ses collègues interprètent la présence de plumes chez Velociraptor comme une preuve contre l'idée que les Maniraptora plus grands et incapables de voler ont perdu leurs plumes secondairement en raison de leur taille corporelle plus grande. En outre, ils notent que les boutons de piquants ne sont presque jamais trouvés chez les espèces d'oiseaux incapables de voler aujourd'hui, et que leur présence dans Velociraptor (présumé avoir été incapable de voler en raison de sa taille relativement grande et de ses membres antérieurs courts) est la preuve que les ancêtres des droméosauridés pouvaient voler, ce qui rend Velociraptor et d'autres grands membres de cette famille secondairement incapables de voler, bien qu'il soit possible que les grandes plumes d'ailes déduites chez les ancêtres de Velociraptor aient eu un but autre que le vol. Les plumes du Velociraptor incapable de voler peuvent avoir été utilisées pour l'affichage, pour couvrir leurs nids pendant la couvaison, ou pour plus de vitesse et de poussée lors de la course sur des pentes inclinées[22]. En raison de la présence d'un autre droméosauridé à Ukhaa Tolgod, Tsaagan, Napoli et l'équipe notent que le renvoi de ce spécimen à Velociraptor ferait actuellement l'objet d'un réexamen[23].
Sens
Les examens de l'endocrâne de Velociraptor indiquent qu'il était capable de détecter et d'entendre une large gamme de fréquences sonores (2 368–3 965 Hz) et pouvait ainsi suivre facilement ses proies. Les examens de l'endocrâne renforcent également la théorie selon laquelle l'animal était un prédateur agile et rapide. Des preuves fossiles suggérant que Velociraptor pratiquait la charogne indiquent également qu'il s'agissait d'un animal opportuniste et d'un prédateur actif, se nourrissant de charognes pendant les périodes de sécheresse ou de famine, s'il était en mauvaise santé ou en fonction de l'âge de l'animal[37].
Alimentation
En 2020, Powers et ses collègues ont réexaminé les maxillaires de plusieurs taxons d'eudroméosaures, concluant que la plupart des eudroméosaures asiatiques et nord-américains étaient séparés par la morphologie du museau et des stratégies écologiques. Ils trouvent que le maxillaire est une référence fiable pour déduire la forme du prémaxillaire et du museau global. Par exemple, la plupart des espèces asiatiques ont des museaux allongés basés sur le maxillaire (à savoir les Velociraptorinae), indiquant une alimentation sélective chez Velociraptor et ses proches parents, comme la capture de petites proies rapides. En revanche, la plupart des eudroméosaures nord-américains, principalement des Dromaeosaurinae, présentent une morphologie du maxillaire robuste et profonde. Cependant, le grand Achillobator est une exception unique aux taxons asiatiques avec son maxillaire profond[38].
Comportement de prédateur
.jpg.webp)
.jpg.webp)
Le fossile Fighting Dinosaurs, découvert en 1971, préserve un V. mongoliensis et un Protoceratops andrewsi en plein combat et fournit des preuves directes d'un comportement de prédateur. Lors du rapport initial, les chercheurs ont émit l'hypothèse que les deux animaux se sont noyés[5]. Cependant, comme les animaux ont été préservés dans d'anciens dépôts de dunes de sable, il en est actuellement admis maintenant que les animaux ont été enterrés dans le sable, soit à partir d'une dune qui s’est effondrée, soit dans une tempête de sable. L'inhumation a dû être extrêmement rapide à en juger par les poses réalistes dans lesquelles les animaux ont été conservés. Des parties du Protoceratops sont manquantes, ce qui a été considéré comme une preuve de comportement de charognards d'autres animaux[39]. Les comparaisons entre les anneaux sclérotiques de Velociraptor, Protoceratops, les oiseaux et reptiles modernes indiquent que Velociraptor peut avoir été nocturne, tandis que Protoceratops peut avoir été cathéméral, actif tout au long de la journée pendant de courts intervalles, suggérant que le combat peut avoir eu lieu au crépuscule ou pendant des périodes de basses conditions lumineuses[40].

La griffe distinctive, sur le deuxième doigt des droméosauridés, a traditionnellement été représentée comme une arme tranchante. Son utilisation présumée étant de couper et d'éventrer des proies[41]. Dans le fossile Fighting Dinosaurs, le Velociraptor se trouve en dessous, avec l'une de ses griffesen forme de faucille apparemment enfoncée dans la gorge de sa proie, tandis que le bec de Protoceratops est serré sur la patte avant droite de son attaquant. Cela suggère que Velociraptor a peut-être utilisé sa griffe pour percer les organes vitaux de la gorge, tels que la veine jugulaire, l’artère carotide ou la trachée plutôt que de couper l'abdomen. Le bord intérieur de la griffe est arrondi et pas exceptionnellement tranchant, ce qui aurait pu empêcher toute sorte d'action coupante ou tranchante, bien que seul le noyau osseux de la griffe soit préservé. L'épaisse paroi abdominale de la peau et des muscles des grandes espèces de proies aurait été difficile à couper sans une surface de coupe spécialisée[39]. L'hypothèse de la coupure a été testée lors d'un documentaire de la BBC sortie en 2005, intitulé The Truth About Killer Dinosaurs (en). Les producteurs du programme ont créé une jambe artificielle de Velociraptor avec une griffe et ont utilisé une poitrine de porc pour simuler la proie du dinosaure. Bien que la griffe ait pénétré la paroi abdominale, elle n'a pas été en mesure de la déchirer, ce qui indique que la griffe n'était pas utilisée pour éventrer une proie[42].
Des restes de Deinonychus, un droméosauridé étroitement apparenté, ont souvent été trouvés dans des groupes de plusieurs individus. Deinonychus a également été trouvé en association avec le grand ornithopode Tenontosaurus, qui a été cité comme preuve de la chasse coopérative en meute[43] - [44]. Cependant, la seule preuve solide d'un comportement social de quelque nature que ce soit chez les droméosauridés provient d'une piste chinoise qui montre six individus d'une grande espèce se déplaçant en groupe[45]. Bien que de nombreux fossiles isolés de Velociraptor aient été trouvés en Mongolie, aucun n'était étroitement associé à d'autres individus[34]. Par conséquent, alors que Velociraptor est généralement décrit comme un chasseur en meute comme dans Jurassic Park, il n'y a que des preuves fossiles limitées pour soutenir cette théorie pour les droméosauridés en général et aucune spécifique au Velociraptor lui-même. Les empreintes de pas des droméosauridés en Chine suggèrent que quelques autres genres ont peut-être chassé en meute, mais aucun exemple concluant de ce type de comportement n'a été trouvé[46] - [47].

En 2011, Denver Fowler et ses collègues suggèrent une nouvelle méthode par laquelle des droméosauridés comme Velociraptor ainsi que des genres apparentés pourraient avoir capturé et retenu des proies. Ce modèle propose que les droméosauridés tuaient leur proie d'une manière très similaire aux rapaces de la famille des accipitridés existants qui sautent sur leur proie, en la coinçant sous leur poids corporel, et le saisissant fermement avec leurs grandes griffes en forme de faucille. Ces chercheurs ont proposé que, comme les accipitridés, les droméosauridés commencerait alors à se nourrir de l'animal lorsqu'il était encore en vie, et que la mort des proies résulterait éventuellement d'une perte de sang et d'une défaillance d'organe. Cette proposition est basée principalement sur des comparaisons entre la morphologie et les proportions des pattes des droméosauridés à plusieurs groupes de rapaces existants avec des comportements de prédateurs connus. Fowler à découvert que les pattes et les jambes des droméosauridés ressemblent le plus à ceux des aigles et des faucons, en particulier en termes d'avoir une deuxième griffe élargie et une amplitude similaire de mouvement de préhension. Le métatarse court et la force de la patte, cependant, auraient été plus similaires à ceux des hiboux. La méthode de prédation serait cohérente avec d'autres aspects de l'anatomie de Velociraptor, tels que leur morphologie inhabituelle de la mâchoire et du bras. Les bras, qui pouvaient exercer beaucoup de force mais étaient probablement recouverts de longues plumes, peuvent avoir été utilisés comme stabilisateurs battants pour l'équilibre au sommet d'une proie en difficulté, avec la queue raide de contrepoids. Les mâchoires, considérées par Fowler et ses collègues comme relativement faibles, auraient été utiles pour les morsures de mouvement de scie à rangée comme le dragon de Komodo moderne, qui a également une morsure faible, pour achever sa proie si les coups de pattes n'étaient pas assez puissants. Ces adaptations prédatrices travaillant ensemble peuvent également avoir des implications sur l'origine du battement d’ailes (en) chez les paraviens[28].
Comportement charognard
En 2010, Hone et ses collègues publient un article sur leur découverte en 2008 de dents qu'ils croyaient être a un Velociraptor près d'un os de la mâchoire marqué par une dent de ce qu'ils croyaient être un Protoceratops dans la formation de Bayan Mandahu. Les auteurs ont conclu que la découverte représentait « une consommation de carcasse à un stade avancé par Velociraptor » car le prédateur aurait mangé d'autres parties d'un Protoceratops fraîchement tué avant de mordre dans la région de la mâchoire[48]. Les preuves ont été considérées comme soutenant l'inférence du fossile Fighting Dinosaurs, selon laquelle Protoceratops faisait partie du régime alimentaire de Velociraptor[49].En 2012, Hone et ses collègues publient un article décrivant un spécimen de Velociraptor avec un os long d'un ptérosaure azhdarchidé dans son intestin. Cela a été interprété comme montrant un comportement charognard[50].
Métabolisme
Velociraptor était un animal à sang chaud dans une certaine mesure, car il avait besoin d'une quantité importante d'énergie pour chasser. Les animaux modernes qui possèdent des manteaux de plumes ou de fourrure, comme Velociraptor, ont tendance à avoir le sang chaud, car ces revêtements fonctionnent comme un isolant. Cependant, les taux de croissance osseuse chez les droméosauridés et certains oiseaux précoces suggèrent un métabolisme plus modéré, par rapport à la plupart des mammifères et oiseaux à sang chaud modernes. Le kiwi est similaire aux droméosauridés dans l'anatomie, le type de plumes, la structure osseuse et même l'anatomie étroite des voies nasales (généralement un indicateur clé du métabolisme). Le kiwi est un oiseau très actif, bien qu’il soit spécialisé, il est incapable de voler. Avec une température corporelle stable et un taux métabolique au repos assez bas, cela fait du kiwi un bon exemple de ce qu’était le métabolisme des oiseaux primitifs et des droméosauridés[33].
Paléopathologies
En 1995, Norell et ses collègues rapportent un crâne de V. mongoliensis portant deux rangées parallèles de petites perforations sur ses os frontaux qui, après un examen plus approfondi, correspondent à l'espacement et à la taille des dents de Velociraptor. Ils suggérent que la blessure avait probablement été infligée par un autre Velociraptor lors d'un combat entre espèces. Étant donné que sa structure osseuse ne montre aucun signe de guérison près des morsures et que l'ensemble du spécimen n'a pas été victime de charognards, cet individu a probablement été tué par cette blessure mortelle[51]. En 2001, Molnar et son équipe notent que ce spécimen est le MPC-D 100/976, originaire de la localité de Tugrik Shireh, où à également été découvert le fossile Fighting Dinosaurs[52].
En 2012, David Hone et son équipe signale un autre spécimen de Velociraptor blessé (MPC-D 100/54, un individu probablement subadultes) trouvé avec les os d'un ptérosaure azhdarchidé dans sa cavité gastrique, portait ou se remettait d'une blessure causé par une fracture des côtes. D'après des preuves sur les os du ptérosaure, qui étaient dépourvus de piqûres ou de déformations dues à la digestion, le Velociraptor est mort peu de temps après, peut-être à cause de la blessure antérieure. Néanmoins, l'équipe a noté que ces côtes cassées montraient des signes de guérison osseuse[50].
Paléoenvironnement
Formation de Bayan Mandahu

Dans les formations de Bayan Mandahu et de Djadokhta, de nombreux fossiles appartenant aux mêmes genres sont présents, mais les espèces diffèrent. Ces variations dans la répartition des espèces peuvent être dues à une barrière naturelle séparant les deux formations, qui sont relativement proches l'une de l'autre géographiquement[12]. Cependant, étant donné l'absence de toute barrière connue qui expliquerait les compositions fauniques spécifiques trouvées dans ces zones, il est plus probable que ces différences indiquent une légère variation dans le temps[53].
V. osmolskae vivait aux côtés de l'ankylosauridé Pinacosaurus mephistocephalus, l’alvarezsauridé Linhenykus, le droméosauridé étroitement apparenté Linheraptor, les oviraptoridés Machairasaurus et Wulatelong, les protocératopsidés Bagaceratops et Protoceratops hellenikorhinus et les troodontidés Linhevenator, Papiliovenator et Philovenator[53]. Les sédiments constituants cette formation indiquent un environnement de dépôt similaire à celui de la formation de Djadochta[54].
Formation de Djadochta

Des spécimens connus de V. mongoliensis ont été découverts dans la formation de Djadochta (également orthographié Djadokhta), dans la province mongole d'Ömnögovi. Il est estimé que cette formation géologique remonte à l'étage du Campanien du Crétacé supérieur, entre 75 et 71 millions d'années avant aujourd'hui. Des sédiments abondants de sables, de grès et de caliche de la formation de Djadochta ont été déposés par des processus éoliens (en) dans des milieux couverts de champs de dunes de sable et parcourus de cours d'eau intermittents, comme l'indique une sédimentation fluviatile très clairsemée, caractérisant un climat semi-aride[55] - [56] - [57].
La formation de Djadochta est séparée en deux membres : celui de Bayn Dzak et de Turgrugyin. V. mongoliensis est identifié dans les deux et représenté par de nombreux spécimens[55]. Le membre de Bayn Dzak abrite l'oviraptoridé Oviraptor, l’ankylosauridé Pinacosaurus grangeri, le protocératopsidé Protoceratops andrewsi et le troodontidé Saurornithoides[1] - [55]. Le membre plus récent de Tugriken Shireh contient l'« oiseau » primitif Elsornis (en), le droméosauridé Mahakala et l’ornithomimidé Aepyornithomimus [58] - [59].
V. mongoliensis a été trouvé dans de nombreuses gisements parmi les plus célèbres et les plus prolifiques de Djadochta. Le spécimen holotype a été découvert sur le site de Flaming Cliffs (sous-gisement du gisement de Bayn Dzak)[1], tandis que le fossile Fighting Dinosaurs a été trouvé sur le gisement de Tugrik Shire (également connue sous le nom de Tugrugeen Shireh ainsi que dans de nombreuses autres orthographes)[4]. Ce dernier est connu pour son exceptionnelle préservation in situ. Sur la base de l'analyse de la structure des dépôts sédimentaires, il a été conclu qu'un grand nombre de spécimens ont été enterrés vivants lors de puissants événements catastrophiques (tels que des avalanches de sable), communs à ces paléoenvironnements[60].
Importance culturelle


Velociraptor est généralement perçu comme un tueur vicieux et rusé en raison de la façon dont il est présenté dans le roman de 1990 Jurassic Park de Michael Crichton, ainsi que de son adaptation cinématographique réalisée par Steven Spielberg, sortie en 1993. Les « raptors » représentés dans Jurassic Park sont en fait calqués sur le droméosauridé étroitement apparenté Deinonychus. Les paléontologues du roman et du film fouillent un squelette dans le Montana, loin de la région est-asiatique où vivait Velociraptor mais caractéristique de la région de Deinonychus[61]. Crichton a rencontré à plusieurs reprises le découvreur de Deinonychus, John Ostrom, à l'université Yale, pour discuter des détails de la gamme possible de comportements et d'apparence de l'animal. Crichton s'est excusé auprès d'Ostrom pour avoir utilisé le nom Velociraptor à la place de Deinonychus, en raison de la tonalité « plus dramatique ». Selon Ostrom, Crichton a déclaré que le Velociraptor du roman repose sur Deinonychus dans presque tous les détails, et que seul le nom a été changé. Les cinéastes de Jurassic Park ont également demandé tous les articles publiés par Ostrom sur Deinonychus pendant la production[62]. Ils ont dépeint les animaux avec la taille, les proportions et la forme du museau de Deinonychus plutôt que de Velociraptor[63] - [64].
La production sur Jurassic Park ayant commencé avant que la découverte du grand droméosauridé Utahraptor ne soit rendue publique, en 1991, Jody Duncan écrit à propos de cette découverte :
« Plus tard, après que nous ayons conçu et construit le raptor, il y a eu la découverte d'un squelette de raptor dans l'Utah qu'ils ont qualifié de « super-slasher ». Ils ont découvert le plus grand Velociraptor à ce jour et il mesurait 1,70 m, tout comme le nôtre. Alors nous l'avons conçu, nous l'avons construit, puis ils l'ont découvert. Cela me dépasse encore[trad 1]. »
— Jony Duncan[63]
Le nom de Spielberg a été brièvement envisagé pour nommer le nouveau dinosaure en échange du financement du travail sur le terrain, mais aucun accord n'a été établi[65].
Jurassic Park et sa suite intitulé Le Monde Perdu sont sortis avant la découverte que les droméosauridés portaient des plumes, de sorte que le Velociraptor dans les deux films est représenté comme écailleux et sans plumes. Pour Jurassic Park 3, les Velociraptor mâle ont reçu des structures en forme de plumes à l'arrière de la tête et du cou, mais ces structures ne ressemblent pas aux plumes que Velociraptor aurait eu en réalité notamment pour des raisons de continuité[66]. La trilogie Jurassic World, servant de suite à Jurassic Park, ignore les plumes de Velociraptor, adhérant aux dessins des précédents films[67]. Cependant, le paravien apparenté Pyroraptor a été emplumé pour Jurassic World : Le Monde d'après, et d'autres changements ont été effectués tels que le raidissement de la queue pour tenir compte des tendons ossifiés et des mains plus semblables de ce que l'on retrouve chez les autres théropodes non aviens[68].
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Velociraptor » (voir la liste des auteurs).
Notes
- Le taxon Megalosauridae, comme Megalosaurus, fonctionnait comme une sorte de taxon « fourre-tout », où de nombreuses espèces non apparentées étaient alors regroupées[1].
Références
- (en) H. F. Osborn, « Three new Theropoda, Protoceratops zone, central Mongolia », American Museum Novitates, no 144, , p. 1-12 (OCLC 40272928, hdl 2246/3223
 , lire en ligne [PDF])
, lire en ligne [PDF]) - (en) H. F. Osborn, « The discovery of an unknown continent », Natural History, vol. 24, no 2, , p. 133-149 (lire en ligne)
- (ru) Rinchen Barsbold, « Carnivorous dinosaurs from the Cretaceous of Mongolia », Transactions of the Joint Soviet-Mongolian Paleontological Expedition, vol. 19, , p. 5-119 (lire en ligne [PDF])
- (en) Zofia Kielan-Jaworowska et Rinchen Barsbold, « Narrative of the Polish-Mongolian Paleontological Expeditions », Paleontologica Polonica, vol. 27, , p. 11 (lire en ligne [PDF])
- (en) Rinchen Barsbold, « Saurornithoididae, a new family of theropod dinosaurs from Central Asia and North America », Paleontologica Polonica, vol. 30, , p. 5-22 (lire en ligne [PDF]).
- (en) American Museum of Natural History, « Fighting Dinosaurs: New Discoveries from Mongolia: Exhibition Highlights » [archive du ], c. 2000 (consulté le ).
- (en) T. Jerzykiewicz, P. J. Currie, D. A. Eberth, P. A. Johnston, E. H. Koster et Jia-Jian Zheng, « Djadokhta Formation correlative strata in Chinese Inner Mongolia: an overview of the stratigraphy, sedimentary geology, and paleontology and comparisons with the type locality in the pre-Altai Gobi », Canadian Journal of Earth Sciences, vol. 30, no 10, , p. 2180-2195 (DOI 10.1139/e93-190, Bibcode 1993CaJES..30.2180J).
- (en) Mark Norell et Peter J. Makovicky, « Important features of the dromaeosaurid skeleton. II: Information from newly collected specimens of Velociraptor mongoliensis », American Museum novitates, vol. 3282, , p. 1-45 (hdl 2246/3025).
- (en) Mark Norell et Peter J. Makovicky, « Important features of the dromaeosaur skeleton : information from a new specimen », American Museum Novitates, vol. 3215, , p. 1-28 (hdl 2246/3557).
- (en) Michael J. Novacek, Dinosaurs of the Flaming Cliffs, New York, Anchor Books, 327 p. (ISBN 0-385-47774-0).
- (en) Alan H. Turner, Shaena Montanari et Mark Norell, « A new dromaeosaurid from the Late Cretaceous Khulsan locality of Mongolia », American Museum Novitates, vol. 3965, , p. 1-48 (DOI 10.1206/3965.1, S2CID 231597229, lire en ligne).
- (en) Pascal Godefroit, Philip J. Currie, Li Hong, Shang Chang Yong et Dong Zhi-Ming, « A new species of Velociraptor (Dinosauria: Dromaeosauridae) from the Upper Cretaceous of northern China », Journal of Vertebrate Paleontology, vol. 28, no 2, , p. 432-438 (DOI 10.1671/0272-4634(2008)28[432:ANSOVD]2.0.CO;2, JSTOR 20490961, S2CID 129414074)
- (en) David C. Evans, Derek W. Larson et Philip J. Currie, « A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America », Naturwissenschaften, vol. 100, no 11, , p. 1041-1049 (PMID 24248432, DOI 10.1007/s00114-013-1107-5, Bibcode 2013NW....100.1041E, S2CID 14978813).
- (en) M. Kundrát, « Two Morphotypes of the Velociraptor Neurocranium » (Conference Abstract), Journal of Morphology, vol. 260, no 3, , p. 305 (PMID 15124236, DOI 10.1002/jmor.10224, S2CID 221869849, lire en ligne [PDF])
- (en) M. J. Powers, « The Evolution of Snout Shape in Eudromaeosaurians and its Ecological Significance » (Master Thesis), Department of Biological Sciences, University of Alberta, (DOI 10.7939/r3-hz8e-5n76, S2CID 235035584, lire en ligne [PDF])
- (en) M. A.Powers, C. Sullivan et P. J. Currie, « Re-examining ratio based premaxillary and maxillary characters in Eudromaeosauria (Dinosauria: Theropoda): Divergent trends in snout morphology between Asian and North American taxa », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 547, no 109704, , p. 109704 (DOI 10.1016/j.palaeo.2020.109704, Bibcode 2020PPP...547j9704P, S2CID 216499705)
- (en) M. J. Powers, M. A. Norell et P. J. Currie, « New shallow snouted species of Velociraptor sheds light on intraspecific variation in Velociraptor mongoliensis and possible niche partitioning between species », Vertebrate Anatomy Morphology Palaeontology, vol. CSVP Online Abstracts, , p. 31 (lire en ligne)
- (en) M. J. Powers, M. A. Norell et P. J. Currie, « Examination of morphological variation across Velociraptor mongoliensis specimens reveals a new species with possible ecomorphological variation in snout dimensions », The Society of Vertebrate Paleontology, vol. Virtual Meeting Conference, , p. 211 (lire en ligne [PDF])
- (en) Gregory S. Paul, Predatory dinosaurs of the world : a complete illustrated guide, New York, Simon & Schuster, (ISBN 978-0-671-61946-6), p. 370
- (en) Alan H. Turner, Diego Paul, Julia A. Clarke, Gregory M. Erickson et Mark Norell, « A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight », Science, vol. 317, no 5843, , p. 1378-1381 (PMID 17823350, DOI 10.1126/science.1144066, JSTOR 20037757, Bibcode 2007Sci...317.1378T, S2CID 2519726, lire en ligne)
- (en) Nicolas E. Campione et David C. Evans, « The accuracy and precision of body mass estimation in non-avian dinosaurs », Biological Review, vol. 95, no 6, , p. 1759-1797 (PMID 32869488, DOI 10.1111/brv.12638, S2CID 221404013).
- (en) Alan H. Turner, Peter J. Makovicky et Mark Norell, « Feather Quill Knobs in the Dinosaur Velociraptor », Science, vol. 317, no 5845, , p. 1721 (PMID 17885130, DOI 10.1126/science.1145076, Bibcode 2007Sci...317.1721T, S2CID 11610649, lire en ligne).
- (en) J. G. Napoli, A. A. Ruebenstahl, B.-A. S. Bhullar, A. H. Turner et M. A. Norell, « A New Dromaeosaurid (Dinosauria: Coelurosauria) from Khulsan, Central Mongolia », American Museum Novitates, no 3982, , p. 1–47 (ISSN 0003-0082, DOI 10.1206/3982.1, hdl 2246/7286 , S2CID 243849373, lire en ligne [PDF]).
- (en) D. W. E. Hone, H. Tischlinger, X. Xu et F. Zhang, « The Extent of the Preserved Feathers on the Four-Winged Dinosaur Microraptor gui under Ultraviolet Light », PLOS ONE, vol. 5, no 2, , e9223 (PMID 20169153, PMCID 2821398, DOI 10.1371/journal.pone.0009223 , Bibcode 2010PLoSO...5.9223H)
- (en) J. Lü et S. L. Brusatte, « A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution », Scientific Reports, vol. 5, no 1, , p. 11775 (PMID 26181289, PMCID 4504142, DOI 10.1038/srep11775 , Bibcode 2015NatSR...511775L)
- (en) Hans-Dieter Sues, « The skull of Velociraptor mongoliensis, a small cretaceous theropod dinosaur from Mongolia », Paläontologische Zeitschrift, vol. 51, nos 3-4, , p. 173-184 (DOI 10.1007/BF02986567, S2CID 128478862)
- (en) Rinchen Barsbold et Halszka Osmólska, « The skull of Velociraptor (Theropoda) from the Late Cretaceous of Mongolia », Acta Palaeontologica Polonica, vol. 44, no 2, , p. 189-219 (résumé, lire en ligne [PDF]).
- (en) Denver W. Fowler, Elizabeth A. Freedman, John B. Scannella et Robert E. Kambic, « The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds », PLOS ONE, vol. 6, no 12, , e28964 (PMID 22194962, PMCID 3237572, DOI 10.1371/journal.pone.0028964 , Bibcode 2011PLoSO...628964F).
- (en) Philip J. Currie, « New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda) », Journal of Vertebrate Paleontology, vol. 15, no 3, , p. 576–591 (DOI 10.1080/02724634.1995.10011250, JSTOR 4523653, S2CID 84544225)
- (en) Mark Norell, James M. Clark, Alan H. Turner et Peter J. Makovicky, « A New Dromaeosaurid Theropod from Ukhaa Tolgod (Ömnögov, Mongolia) », American Museum Novitates, vol. 3545, , p. 1–51 (DOI 10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2, hdl 2246/5823 , S2CID 54705292, lire en ligne)
- (en) Pascal Godefroit, Andrea Cau, Hu Dong-Yu, François Escullié, Wu Wenhao et Gareth Dyke, « A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds », Nature, vol. 498, no 7454, , p. 359-362 (PMID 23719374, DOI 10.1038/nature12168, Bibcode 2013Natur.498..359G, S2CID 4364892, lire en ligne [PDF])
- (en) Robert A. De Palma, David A. Burnham, Larry D. Martin, Peter L. Larson et Robert T. Bakker, « The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation », Paleontological Institute, vol. 14, (DOI 10.17161/paleo.1808.18764 , S2CID 17099603, lire en ligne [PDF])
- (en) Gregory S. Paul, Dinosaurs of the air : the evolution and loss of flight in dinosaurs and birds, Baltimore, Johns Hopkins University Press, (ISBN 978-0-8018-6763-7, lire en ligne)
- (en) Mark A. Norell et Peter J. Makovicky, « Dromaeosauridae », dans David B. Weishampel, Peter Dodson et Halszka Osmólska, The Dinosauria, Berkeley, University of California Press, , 2e éd., 861 p. (ISBN 978-0-520-24209-8), p. 196-209
- (en) Philip J. Currie et David C. Evans, « Cranial Anatomy of New Specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta », The Anatomical Record, vol. 303, no 4, , p. 691-715 (PMID 31497925, DOI 10.1002/ar.24241, S2CID 202002676)
- (en) Steven E. Jasinski, Robert M. Sullivan et Peter Dodson, « New Dromaeosaurid Dinosaur (Theropoda, Dromaeosauridae) from New Mexico and Biodiversity of Dromaeosaurids at the end of the Cretaceous », Scientific Reports, vol. 10, no 1, , p. 5105 (ISSN 2045-2322, PMID 32218481, PMCID 7099077, DOI 10.1038/s41598-020-61480-7 , Bibcode 2020NatSR..10.5105J)
- (en) J. Logan King, Justin S. Sipla, Justin A. Georgi, Amy M. Balanoff et James M. Neenan, « The endocranium and trophic ecology of Velociraptor mongoliensis », Journal of Anatomy, vol. 237, no 5, , p. 861-869 (PMID 32648601, PMCID 7542195, DOI 10.1111/joa.13253 )
- (en) M. A. Powers, C. Sullivan et P. J. Currie, « Re-examining ratio based premaxillary and maxillary characters in Eudromaeosauria (Dinosauria: Theropoda): Divergent trends in snout morphology between Asian and North American taxa », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 547, , p. 109704 (DOI 10.1016/j.palaeo.2020.109704, Bibcode 2020PPP...547j9704P, S2CID 216499705)
- (en) Kenneth Carpenter, « Evidence of predatory behavior by theropod dinosaurs », Gaia, vol. 15, , p. 135-144 (lire en ligne [archive du ] [PDF])
- (en) Lars Schmitz et Ryosuke Motani, « Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology », Science, vol. 332, no 6030, , p. 705-8 (PMID 21493820, DOI 10.1126/science.1200043, Bibcode 2011Sci...332..705S, S2CID 33253407)
- (en) John H. Ostrom, « Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana », Bulletin of the Peabody Museum of Natural History, vol. 30, , p. 1-165 (DOI 10.2307/j.ctvqc6gzx, S2CID 210733704)
- (en) Phillip L. Manning, David Payne, John Pennicott, Paul M. Barrett et Roland A. Ennos, « Dinosaur killer claws or climbing crampons ? », Biology Letters, vol. 2, no 1, , p. 110-112 (PMID 17148340, PMCID 1617199, DOI 10.1098/rsbl.2005.0395 )
- (en) W. Desmond Maxwell et John H. Ostrom, « Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations », Journal of Vertebrate Paleontology, vol. 15, no 4, , p. 707-712 (DOI 10.1080/02724634.1995.10011256, S2CID 84382803, lire en ligne [archive du ])
- (en) Daniel L. Brinkman, Richard L. Cifelli et Nicholas J. Czaplewski, « First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) in the Antlers Formation (Lower Cretaceous: Aptian-Albian) of Oklahoma », Oklahoma Geological Survey Bulletin, vol. 146, , p. 1–27 (lire en ligne [PDF])
- (en) Rihui Li, Martin G. Lockley, Peter J. Makovicky, Masaki Matsukawa, Mark A. Norell, Jerald D. Harris et Mingwei Liu, « Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China », Naturwissenschaften, vol. 95, no 3, , p. 185-191 (DOI 10.1007/s00114-007-0310-7, Bibcode 2008NW.....95..185L, S2CID 16380823)
- (en) Brian Switek, « Dinosaurs behaving badly: Did velociraptors hunt in packs ? », The Guardian, (lire en ligne )
- (en) John Long et Peter Schouten, Feathered Dinosaurs: The Origin of Birds., Oxford University Press, (ISBN 978-0-19-537266-3), Page 21
- (en) Matt Walker, « Fossil find shows Velociraptor eating another dinosaur », BBC Earth News, (lire en ligne)
- (en) David Hone, Jonah Choiniere, Corwin Sullivan, Xing Xu, Michael Pittman et Qingwei Tan, « New evidence for a trophic relationship between the dinosaurs Velociraptor and Protoceratops », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 291, vol. 291, nos 3-4, , p. 488-492 (DOI 10.1016/j.palaeo.2010.03.028, Bibcode 2010PPP...291..488H, S2CID 55857507)
- (en) David Hone, Takanobu Tsuihiji, Mahito Watabe et Khishigjaw Tsogtbaatr, « Pterosaurs as a food source for small dromaeosaurs », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 331-332, , p. 27-30 (DOI 10.1016/j.palaeo.2012.02.021, Bibcode 2012PPP...331...27H, S2CID 85997141)
- (en) Mark Norell, Discovering dinosaurs, Knopf Inc., (ISBN 9780520225015, lire en ligne), p. 43
- (en) Kenneth Carpenter, Michael William Skrepnick et Darren H. Tanke, Mesozoic vertebrate life, Bloomington (Indiana), Indiana University Press, (ISBN 9780253339072, OCLC 248649755), « Theropod Paleopathology: A Literature Survey », p. 337-363
- (en) Nicholas R. Longrich, Philip J. Currie et Dong Zhi-Ming, « A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia », Palaeontology, vol. 53, no 5, , p. 945-960 (DOI 10.1111/j.1475-4983.2010.00968.x , S2CID 128767044).
- (en) D. A. Eberth, « Depositional environments and facies transitions of dinosaur-bearing Upper Cretaceous redbeds at Bayan Mandahu (Inner Mongolia, People's Republic of China) », Canadian Science Publishing, vol. 30, no 10, , p. 2196-2213 (DOI 10.1139/e93-191, Bibcode 1993CaJES..30.2196E, S2CID 129911684).
- (en) Demberelyin Dashzeveg, Lowell Dingus, David B. Loope, Carl C. Swisher III, Togtokh Dulam et Mark R. Sweeney, « New Stratigraphic Subdivision, Depositional Environment, and Age Estimate for the Upper Cretaceous Djadokhta Formation, Southern Ulan Nur Basin, Mongolia », American Museum Novitates, no 3498, , p. 1-31 (DOI 10.1206/0003-0082(2005)498[0001:NSSDEA]2.0.CO;2, hdl 2246/5667 , S2CID 55836458, lire en ligne [PDF])
- (en) Kevin Padian, Encyclopedia of dinosaurs, San Diego : Academic Press, (ISBN 978-0-12-226810-6), p. 188-191
- (en) Lowell Dingus, David B. Loope, Dėmbėrėliĭn Dashzėvėg, Carl Celso Swisher, Chuluun Minjin, Michael J. Novacek et Mark Norell, « The geology of Ukhaa Tolgod (Djadokhta Formation, Upper Cretaceous, Nemegt Basin, Mongolia) », American Museum Novitates, no 3616, , p. 1-40 (DOI 10.1206/442.1, hdl 2246/5916 , S2CID 129735494, lire en ligne).
- (en) Alan H. Turner, Diego Pol, Julia A. Clark, Gregory M. Erickson et Mark Norell, « A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight », Science, vol. 317, no 5843, , p. 1378-1381 (PMID 17823350, DOI 10.1126/science.1144066, Bibcode 2007Sci...317.1378T, S2CID 2519726)
- (en) Tsogtbaatar Chinzorig, Yoshitsugu Kobayashi, Khishigjav Tsogtbaatar, Philip J. Currie, Mahito Watabe et Rinchen Barsbold, « First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia », Scientific Reports, vol. 7, no 1, , p. 5835 (PMID 28724887, PMCID 5517598, DOI 10.1038/s41598-017-05272-6 , Bibcode 2017NatSR...7.5835C)
- (en) Mototaka Saneyoshi, Mahito Watabe, Shigeru Suzuki et Khishigjav Tsogtbaatar, « Trace fossils on dinosaur bones from Upper Cretaceous eolian deposits in Mongolia: Taphonomic interpretation of paleoecosystems in ancient desert environments », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 311, nos 1-2, , p. 38-47 (DOI 10.1016/j.palaeo.2011.07.024, Bibcode 2011PPP...311...38S, lire en ligne [PDF])
- (en) Michael Crichton, Jurassic Park : a novel, New York : Knopf : Distributed by Random House, , 399 p. (ISBN 978-0-394-58816-2, lire en ligne), Page 117.
- (en) Mike Cummings, « Yale’s legacy in ‘Jurassic World’ », YaleNews, (lire en ligne).
- (en) J. Duncan, The Winston Effect. London: Titan Books (ISBN 978-1-84576-365-7), Page 171
- (en) R. T. Bakker, Raptor Red (ISBN 978-0-553-57561-3), Page 4.
- (en) Brook Adams, « Director Loses Utahraptor Name Game », Deseret News, (lire en ligne).
- (en) Michael Dhar, « T. Rex at 20: How ‘Jurassic Park’ science has evolved », The Washington Post, (lire en ligne)
- (en) Susana Polo, « Jurassic World explains its featherless dinos while poking fun at blockbusters », Polygon, (lire en ligne)
- (en) Amit Katwala, « The Real Story Behind Jurassic World Dominion’s Dino Feathers », Wired, (lire en ligne)
Citations originales
- « Later, after we had designed and built the raptor, there was a discovery of a raptor skeleton in Utah, which they labeled 'super-slasher.' They had uncovered the largest Velociraptor to date and it measured five-and-a-half-feet tall, just like ours. So we designed it, we built it, and then they discovered it. That still boggles my mind. »
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :
- Global Biodiversity Information Facility
- (cs + en) BioLib
- (en) Paleobiology Database
- (mul + en) iNaturalist
- (en) Interim Register of Marine and Nonmarine Genera
- Notices dans des dictionnaires ou encyclopédies généralistes :