Caseopsis
Caseopsis agilis

Caseopsis est un genre éteint et fossile de synapsides Caseidae ayant vécu à la fin du Permien inférieur (Kungurien) ou au début du Permien moyen (Roadien) dans ce qui est aujourd’hui le Texas, États-Unis. Le genre n’est représenté que par son espèce type, Caseopsis agilis, qui fut nommée en 1962 par Everett C. Olson à partir d’un squelette fragmentaire découvert dans la formation de San Angelo, dans le Comté de Knox au Texas.
Présentation
Des os des membres trouvés dans des strates légèrement plus jeune lui sont également attribués de façon incertaine[1] - [2].
Description
Caseopsis est un caséidé de taille moyenne connu par des restes très fragmentaires. L’holotype FMNH UR 253 est représenté par une partie du crâne et des mâchoires, une vertèbre lombaire, des fragments d’autres vertèbres, plusieurs côtes, une partie de la scapula gauche, un radius, un ulna, le bassin, un fémur, des parties du tibia et de la fibula, et quelques os du pied. Il fut découvert dans un niveau de mudstone rouge sableux, à la base de la partie supérieure de la formation de San Angelo, dans le comté de Knox au Texas. D’après Olson, le squelette devait être presque complet au moment du dépôt, mais une décomposition chimique des os après l’enfouissement et l’érosion en ont détruit une partie considérable avant la découverte. Un second squelette découvert dans la même région du Texas provient d’un niveau stratigraphique légèrement plus récent composé de mudstones rouges situés au sommet de la formation de San Angelo[N 1]. Ce spécimen, un peu plus grand que l’holotype, est plus fragmentaire et est composé d’une extrémité proximale de radius, d’un tibia, d’une fibula, d’un fragment d’extrémité distale de fémur, et des fragments de côtes. Il est attribué à Caseopsis cf. agilis car ses maigres vestiges ne permettent pas de le placer ou de le distinguer de l’espèce type avec certitude[1] - [2].
D’après Olson, le crâne serait représenté par plusieurs fragments mais seul un maxillaire incomplet a été figuré[1] - [2]. Celui-ci suggère que le crâne devait être très petit comparativement au reste du corps, comme c’est le cas chez de nombreux autres caséidés. Ce maxillaire a conservé six dents plus ou moins complètes, lesquelles sont pourvues de trois petites cuspides[2]. Le squelette post-crânien se distinguerait principalement par des éléments squelettiques légèrement construits. Les os des membres en particulier sont décrits comme longs et minces, et la lame de l’ilion est haute. Les surfaces articulaires des os des membres sont petites par rapport à celles d’autres caséidés de taille comparable. Ainsi, pour Olson, la construction légère de l’animal et la hauteur des lames de ses ceintures indique que Caseopsis était vraisemblablement adaptés à une locomotion un peu plus rapide que celle des autres caséidés contemporains[1] - [2]. Cependant, Romano et Nicosia ont démontré que les os des membres de C. agilis n’étaient pas tous d’une gracilité excessive et qu’ils montraient, dans plusieurs cas, une plus grande robustesse en comparaison de la plupart des autres caséidés[3]. Le radius notamment est proportionnellement plus robuste que celui de Cotylorhynchus qui est pourtant un caséidé nettement plus gros[4]. Caseopsis possédait ainsi des membres antérieurs plus robustes que les membres postérieurs comme cela semble avoir été le cas chez quelques autres caséidés. Pour Romano, il est également possible que Caseopsis représente l’un des stades juvéniles du genre Cotylorhynchus. Une hypothèse difficile à tester en raison notamment du manque de séries ontogénétiques suffisamment complètes chez les grands caséidés[4]. Si Caseopsis est bien un genre valide, alors la présence de membres antérieurs robustes serait indépendante de la nécessité de supporter une taille corporelle énorme. Cela signifierait que ce n’est que par la suite que la structure a été exaptée par des caséides plus dérivées et plus gros, pour des raisons de support ou pour une fonction spécifique particulière des membres antérieurs[4].
Répartition stratigraphique
L’âge de la formation de San Angelo a fait l’objet de nombreuses interprétations, celles-ci lui attribuant alternativement un âge Cisuralien terminal (Kungurien) et/ou Guadalupien basal (Roadien) [5]. La formation de San Angelo surmonte le Groupe de Clear Fork et est recouverte par la formation de Blaine. D’après Spencer G. Lucas et d’autres auteurs, des fusulines trouvées dans une intercalation marine de la formation de San Angelo, ainsi que des ammonoïdes présentes à la base de la formation de Blaine sus-jacente, indiquaient un âge Kungurien. De plus, selon ces auteurs, la base de la formation de San Andres, située plus à l’ouest et considérée comme un équivalent latéral de la formation de Blaine, se trouve dans la zone de conodontes de Neostreptognathodus prayi, la seconde des trois biozones de conodontes du Kungurien. La base de la formation de Blaine appartiendrait donc à cette biozone du Kungurien, ce qui suggère que la formation de San Angelo sous-jacente et Caseopsis seraient légèrement plus anciens que la zone de conodonte à N. prayi avec un âge Kungurien inférieur[6] - [7] - [8]. Cependant, Michel Laurin et Robert W. Hook ont fait valoir que l’intercalation marine à fusulines citée plus haut n’appartient pas à la formation de San Angelo dans laquelle elle fut incluse par erreur, et ne peut pas être utilisée pour dater cette dernière. Le nom de formation de San Angelo a été incorrectement appliqué à une grande variété de roches dans divers bassins sédimentaires situés dans l’ouest du Texas, alors que la Formation de San Angelo est limitée au plateau oriental et est exclusivement continentale et dépourvue de fossiles marins[5]. Par ailleurs, la révision taxonomique des ammonoïdes de la base de la formation de Blaine indique un âge Roadien plutôt que Kungurien[N 2] - [5]. La formation de San Angelo a quant à elle livré une flore fossile dominée par les les conifères voltziens, un assemblage plutôt caractéristique du Guadalupien et du Lopingien. Ainsi, selon Laurin et Hook la formation de San Angelo pourrait dater du Kungurien terminal ou du Roadien basal, ou plus probablement pourrait chevaucher la limite Kungurien/Roadien[5].
Paléoenvironnements
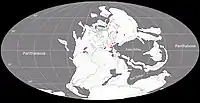
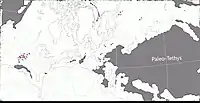
Au Permien, la plupart des terres émergées étaient réunies en un seul supercontinent, la Pangée. Celui-ci avait alors grossièrement la forme d’un C : ses parties nord (la Laurasia) et sud (le Gondwana) étaient reliés à l'ouest mais séparés à l'est par une très vaste baie océanique - la mer de Téthys[9]. Un long chapelet de microcontinents, regroupés sous le nom de Cimmérie, divisait la Téthys en deux : la Paleo-Tethys au nord, et la Neo-Tethys au sud[10]. La formation de San Angelo correspond à des sédiments fluviatiles et/ou éoliens déposés dans une vaste plaine deltaïque parsemée de lacs et de lagunes. Cette plaine côtière était bordée à l’ouest par une mer qui occupait ce qui est maintenant le Golfe du Mexique et la partie la plus méridionale de l'Amérique du Nord. Les cours d’eau alimentant le delta se dirigeaient depuis des reliefs modestes situés plus à l’est et correspondant aux soulèvements ancestraux des montagnes Ouachita, Arbuckle et Wichita. Le climat était subtropical avec des pluies modérées et saisonnières. Il existait une mousson d’été ainsi qu’une saison sèche hivernale. La mousson était relativement faible en raison de la taille limitée de la mer et du faible différentiel entre les températures d’été et d’hiver. La présence d’évaporites indique une aridité importante interrompue par des inondations saisonnières[11] - [1] - [12] - [13] - [14] - [8].
La formation de San Angelo est composée à sa base de grès dur, vert, gris et bruns, et de conglomérats fins, non fossilifères. La partie centrale de la formation est principalement constituée de mudstones rouges interprétés comme des boues argileuses et limoneuses déposées dans les plaines côtières lors d’épisodes d’inondations périodiques. Ces mudstones rouges sont intercalés d’un mince niveau de grès vert, de mudstones sableux, et d’évaporites. Ceux-ci correspondraient à un empiètement mineur et de courte durée des estuaires, des lagunes, et des mers très peu profondes sur la partie terrestre du delta. Les caséidés Angelosaurus dolani et Caseoides sanangeloensis sont présents dans les mudstones rouges de cette partie de la formation[N 3]. La partie supérieure de la formation de San Angelo est caractérisée par la prépondérance des sédiments grossiers tels que grès et conglomérats, mais incluant aussi à sa base des mudstones sableux et à son sommet des mudstones rouges purs. D’après Olson, ces sédiments furent déposés par des rivières plus larges et aux courants plus puissants que celles de la partie centrale de la formation[1]. Cependant, en Oklahoma, les strates équivalentes à la formation de San Angelo, qui étaient elles aussi considérées comme des dépôts fluviaux-deltaïques et fluviaux côtiers, ont été réinterprétées comme étant d’origine éolienne[15]. La partie supérieure non sommitale de la formation de San Angelo est caractérisée par l’absence du caséidé Angelosaurus et l’abondance de Cotylorhynchus hanckoki. Caseopsis est un composant mineur de cette faune qui comprend, entre autres[N 4], le sphénacodontidé Dimetrodon angelensis, les Captorhinidae Rothianiscus multidonta, et Kahneria seltina, et le dvinosaure Tupilakosauridae Slaugenhopia[11] - [1]. Quelques pistes de tétrapodes indiquent également la présence de paréiasauromorpha nycteroleteridés (ichnotaxon Pachypes ollieri)[16], dont un squelette partiel est connu dans les dépôts légèrement plus récents de la formation de Chickasha en Oklahoma[17] - [16]. Dans les mudstones rouges du sommet de la formation, le spécimen Caseopsis cf. agilis a été trouvé avec un autre caséidé, Angelosaurus greeni[1]. Une flore inhabituelle a été retrouvée dans les chenaux de la partie supérieure de la formation de San Angelo. Elle est dominée par les gymnospermes et est remarquable par sa composition unique incluant à la fois des taxons typiques du Permien inférieur tels Walchia ou Culmitzschia mais aussi des formes qui n’étaient jusque-là connus que dans le Permien moyen ou supérieur comme diverses espèces d’Ulmannia, Pseudovoltzia liebeana, et le taxon d’affinité incertaine Taeniopteris eckardtii, ou dans les strates du Mésozoïque comme la bennettitale Podozamites et le cycadidé putatif Dioonitocarpidium. Le reste de la flore est représentée par le ginkgoale Dicranophyllum, la cordaitale Cordaites, et l’équisetale cf. Neocalamites[18] - [19].
Phylogénie
En 1968 Olson suggérait que Caseopsis était le seul représentant connu d’une ligné particulière de caséidés aux membres graciles[2]. Cette hypothèse a été invalidé en 2015 par une analyse cladistique des caséidés réalisée par Romano et Nicosia dans laquelle Caseopsis est identifié comme étant proche de caséidés aux proportions plutôt robustes par rapport à leur taille. Dans cette étude, Caseopsis agilis est le groupe frère de “Casea” nicholsi, ces deux taxons appartenant à un clade incluant également Caseoides sanangeloensis[3].
Ci-dessous, le cladogramme des Caseidae publié par Romano et Nicosia en 2015[3].
Une étude phylogénétique publiée en 2022 par Ralf Werneburg et des collègues indique que Caseopsis pourrait former un clade avec Ruthenosaurus et que ce clade formerait une polytomie avec l’espèce Cotylorhynchus romeri, ainsi qu’avec un clade incluant Alierasaurus, les deux autres espèces de Cotylorhynchus et Lalieudorhynchus[20].
Ci-dessous, le cladogramme publié par Werneburg et des collègues en 2022[21].
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- Ce niveau de mudstones rouges fut originellement considéré par Olson (1962) comme appartenant à la base de la formation de Flowerpot. En 1974, Gary E. Smith a redéfini ces mudstones comme formant le sommet de la formation de San Angelo
- Pris individuellement les sept genres d’ammonoïdes de cette faune ont une distribution temporelle s’étendant du Cisuralien au Lopingien : trois sont connus dans les gisements du Cisuralien et du Guadalupien, deux s'étendent du Cisuralien au Lopingien, et deux sont enregistrés dans les roches du Guadalupien au Lopingien inférieur. Cependant, le Roadien (= base du Guadalupien) est le seul étage durant lequel les sept genres coexistent.
- De ces niveaux proviennent également les genres très fragmentaires Steppesaurus et Mastersonia. En 1953, Olson et Beerbower classèrent d’abord Steppesaurus parmi les pélycosaures Sphenacodontidae. Puis en 1962 Olson le considéra comme un thérapside Phtinosuchidae. Dans le même article Olson décrivit Mastersonia qu’il considérait comme un thérapside « éodinocéphalien ». Dans une courte note publiée en 1995, Sidor et Hopson ont rejeté la nature thérapside de ces deux taxons, leurs restes appartenant selon eux à des pélycosaures. Laurin et Hook (2022) souligne toutefois la nécessité de réétudier ces spécimens avec les méthodes modernes.
- Olson et Beerbower (1953), puis Olson (1962), ont nommés d’autres taxons fragmentaires de la partie supérieure de la formation de San Angelo qu’ils attribuèrent à divers groupes de thérapsides, comme Knoxosaurus, Gorgodon, Dimacrodon, Eosyodon, Driveria, et Tappenosaurus. En 1995 Sidor et Hopson ont réinterprété les fossiles de tous ces taxons comme des restes de pélycosaures caséidés et sphénacodontidés plus ou moins déterminables. Cependant, une nouvelle étude de ces spécimens par des méthodes modernes serait nécessaire (Laurin et Hook, 2022).
Références
- (en) E.C. Olson, « Late Permian terrestrial vertebrates, U.S.A and U.S.S.R. », Transactions of the American Philosophical Society, New Series, vol. 52, , p. 1-224 (DOI 10.2307/1005904)
- (en) E.C. Olson, « The family Caseidae », Fieldianna (Geology), vol. 17, , p. 225–349
- (en) M. Romano et U. Nicosia, « Cladistic analysis of Caseidae (Caseasauria, Synapsida): using the gap-weighting method to include taxa based on incomplete specimens », Palaeontology, vol. 58, no 6, , p. 1109–1130 (DOI 10.1111/pala.12197)
- (en) M. Romano, « Long bone scaling of caseid synapsids: a combined morphometric and cladistic approach », Lethaia, vol. 50, no 4, , p. 511-526 (DOI 10.1111/let.12207)
- (en) M. Laurin et R.W. Hook, « The age of North America’s youngest Paleozoic continental vertebrates : a review of data from the Middle Permian Pease River (Texas) and El Reno (Oklahoma) Groups », BSGF – Earth Sciences Bulletin, vol. 193, no 10, , p. 1-30 (DOI 10.1051/bsgf/2022007)
- (en) S.G. Lucas, « Global Permian tetrapod biostratigraphy and biochronology », dans Lucas, S.G., Cassinis, G. and Schneider, J.W., Non-Marine Permian Biostratigraphy and Biochronology, London, Geological Society, Special Publications, 265, (ISBN 978-1-86239-206-9), p. 65–93
- (en) S.G. Lucas et V.K. Golubev, « Age and duration of Olson’s Gap, a global hiatus in the Permian tetrapod fossil record », Permophiles, vol. 67, , p. 20- 23 (lire en ligne)
- (en) J.W. Schneider, S.G. Lucas, F. Scholze, S. Voigt, L. Marchetti, H. Klein, S. Opluštil, R. Werneburg, V.K. Golubev, J.E. Barrick, T. Nemyrovska, A. Ronchi, M.O. Day, V.V. Silantiev, R. Rößler, H. Saber, U. Linnemann, V. Zharinova et S-Z. Shen, « Late Paleozoic–early Mesozoic continental biostratigraphy — Links to the Standard Global Chronostratigraphic Scale », Palaeoworld, vol. 29, no 2, , p. 186-238 (DOI 10.1016/j.palwor.2019.09.001)
- (en) S. McLoughlin, « The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism », Australian Journal of Botany, vol. 49, , p. 271-300 (DOI 10.1071/BT00023)
- (en) A.M.C. Şengör, « Tectonics of the Tethysides: orogenic collage development in a collisional setting », Annual Review of Earth and Planetary Sciences, vol. 15, , p. 214-244 (DOI 10.1146/annurev.ea.15.050187.001241)
- (en) E.C. Olson et J.R. Beerbower, « The San Angelo formation, Permian of Texas and its vertebrates », The Journal of Geology, vol. 61, no 5, , p. 384-423
- (en) G.E. Smith, « Depositional Systems, San Angelo Formation (Permian), North Central Texas – Facies Control of Red-Bed Copper Mineralization », The University of Texas at Austin Bureau of Economic Geology Report of Investigation, vol. 80, , p. 1-74 (DOI 10.23867/RI0080D)
- (en) T.S. Kemp, « Pelycosaurs », dans Kemp, T.S., Mammal-like reptiles and the origin of Mammals, London, Academic Press, (ISBN 978-0124041202), p. 73
- (en) M.J. Soreghan, M.M. Swift et G.S. Soreghan, « Provenance of Permian eolian and related strata in the North American midcontinent: Tectonic and climatic controls on sediment dispersal in western tropical Pangea », dans Ingersoll, R.V., Lawton, T.F., and Graham, S.A., Tectonics, Sedimentary Basins, and Provenance: A Celebration of William R. Dickinson’s Career, Boulder, Geological Society of America Special Paper 540, (ISBN 978-0813725406, DOI 10.1130/2018.2540(28)), p. 661-688
- (en) T.M. Foster, G.S. Soreghan, M.J. Soreghan, K.C. Benison et R.D. Elmore, « Climatic and paleogeographic significance of eolian sediment in the Middle Permian Dog Creek Shale (Midcontinent U.S.) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 402, , p. 12-29 (DOI 10.1016/j.palaeo.2014.02.031)
- (en) L. Marchetti, S. Voigt, E. Mujal, S.G. Lucas, H. Francischini, J. Fortuny et V.L. Santucci, « Extending the footprint record of Pareiasauromorpha to the Cisuralian : earlier appearance and wider palaeobiogeography of the group », Papers in Palaeontology, vol. 7, no 3, , p. 1297-1319 (DOI 10.1002/spp2.1342)
- (en) R.R. Reisz et M. Laurin, « The reptile Macroleter: First vertebrate evidence for correlation of Upper Permian continental strata of North America and Russia », Geological Society of America Bulletin, vol. 113, no 9, , p. 1229-1233 (DOI 10.1130/0016-7606(2001)113<1229:TRMFVE>2.0.CO;2)
- (en) W.A. DiMichele, S.H. Mamay, D.S. Chaney, R.W. Hook et W.J. Nelson, « An Early Permian flora with Late Permian and Mesozoic affinities from north-central Texas », Journal of Paleontology, vol. 75, no 2, , p. 449-460 (DOI 10.1666/0022-3360(2001)075˂0449:AEPFWL˃2.0.CO;2)
- (en) W.A. DiMichele, N.J. Tabor, D.S. Chaney et W.J. Nelson, « From wetlands to wet spots: Environmental tracking and the fate of Carboniferous elements in Early Permian tropical floras », dans Greb, S.F. & DiMichele, W.A., Wetlands through time, Boulder, The Geological Society of America Special Paper 399, (DOI 10.1130/2006.2399(11)), p. 223-248
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2)
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2).