Lalieudorhynchus
Lalieudorhynchus gandi

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Clade | Amniota |
| Sous-classe | Synapsida |
| Clade | † Caseasauria |
| Famille | † Caseidae |
Werneburg et al., 2022
Werneburg et al., 2022
Lalieudorhynchus est un genre éteint de synapsides Caseidae ayant vécu au Guadalupien (= Permien moyen) dans ce qui est aujourd’hui le sud de la France. Le genre n’est connu que par son espèce type, Lalieudorhynchus gandi, qui fut nommée en 2022 par Ralf Werneburg, Frederik Spindler, Jocelyn Falconnet, Jean-Sébastien Steyer, Monique Vianey-Liaud, et Joerg W. Schneider. Lalieudorhynchus est représenté par un squelette postcrânien partiel découvert dans le bassin de Lodève dans le centre du département de l’Hérault, en région Occitanie. Il appartient à un individu mesurant environ 3,75 m de longueur. Le degré d’ossification de ces os indique toutefois qu’il s’agissait d’un juvénile tardif ou d’un jeune adulte encore en cours de croissance. D’après la structure interne de ses os, Lalieudorhynchus est interprété comme un animal semi aquatique qui aurait pu avoir un mode de vie similaire à celui des hippopotames, passant une partie de son temps dans l’eau mais retournant sur terre pour se nourrir. Il est géologiquement l’un des plus jeunes représentants connus des caséidés. L’analyse phylogénétique proposée par Werneburg et des collègues a identifié Lalieudorhynchus comme un caséidé dérivé étroitement apparenté à l’espèce nord-américaine “Cotylorhynchus” hancocki[1].
Étymologie
Le nom de genre est une combinaison de La Lieude situé à proximité de la localité type, et de « rhynchus », la forme latinisée du grec « rhynchos » (le nez) parfois utilisé pour nommer les caséidés. L’épithète spécifique rend hommage à Georges Gand qui a travaillé sur le bassin de Lodève pendant des décennies, notamment sur la dalle à empreintes de La Lieude, et a co-organisé et promu les campagnes d’excavations de ce caséidé[1].
Description
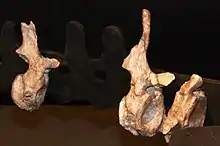
Lalieudorhynchus est représenté par un squelette post-crânien partiel et désarticulé mais bien conservé. L’holotype, représenté par une série d’ossements catalogués UM-LIE 02-37, UM-LIE 39-41, UM-LIE 45 et UM-LIE 47, se compose d’une dizaine de vertèbres (dorsales, sacrales et caudales), une quinzaine de côtes, un scapulo-coracoïde droit complet de 50 cm de long, la branche dorsale de l’ilion gauche, les fémurs gauche et droit mesurant 35,5 cm de long, et plusieurs os du pied (un astragale, deux éléments tarsiens et cinq phalanges). La longueur totale du corps de ce spécimen est estimée à 3,75 m. L’holotype de Lalieudorhynchus montre un mélange de caractères matures et immatures dans l’ensemble de son squelette indiquant que ce spécimen était un juvénile tardif ou un jeune adulte encore en croissance au moment de sa mort[1]. Le crâne n’est pas connu mais comme chez ses plus proches cousins, il devait probablement être très petit comparé à la taille du corps, triangulaire en vue dorsale, et se terminer antérieurement par un museau incliné vers l’avant et pourvu de très grandes narines externes. La forme de ses côtes indique que Lalieudorhynchus avait une cage thoracique en forme de tonneau comme chez les autres caséidés dérivés[1]. Celle-ci devait abriter des intestins volumineux suggérant que l’animal devait se nourrir d’une grande quantité de végétaux à faible valeur nutritive[2].
Lalieudorhynchus est caractérisé par plusieurs apomorphies. Les épines neurales des vertèbres sacrées et des vertèbres caudales antérieures ont une section transversale avec un processus en forme de quille très mince vers l’avant, qui commence au-dessus des prézygapophyses[N 1] et remonte tout le long du bord vertical jusqu’au sommet de l’épine neurale. Les épines neurales des vertèbres dorsales et caudales ont leur extrémité dorsale mince au lieu de montrer un épaississement latéral. La première côte sacrée a une extrémité distale étroite. Le scapulo-coracoïde possède une fosse sur le processus triceps du métacoracoïde (l’un des trois os constituant le scapulo-coracoïde avec le procoracoïde et la scapula). Le pied se caractérise par un très grand tarse distal 1 de même largeur que l’astragale, avec presque tous les côtés légèrement concaves[1].
Lalieudorhynchus se distingue également par une combinaison unique de caractères. Comme d’autres caséidés, il diffère de Ruthenosaurus par ses épines neurales droites au lieu d’être inclinées vers l’avant. Les vertèbres caudales médianes ont des centra (ou corps vertébraux) relativement longs, allongés sous leur postzygapophyses[N 2], mais avec des arcs neuraux bas, contrairement à Alierasaurus. Les trois vertèbres sacrées et la vertèbre caudale la plus antérieure ont un centrum court et transversalement très large comme chez Ruthenosaurus, alors que ces même centra sont beaucoup plus étroit chez Cotylorhynchus romeri. L’épine neurale de la première vertèbre sacrée et de la première vertèbre caudale est très allongée dorsalement comme chez “Cotylorhynchus” hancocki, ce qui n’est pas le cas chez C. romeri et Ruthenosaurus. Un hyposphene[N 3] est présent sous les postzygapophyses des vertèbres dorsales et caudales, un caractère signalé jusqu’ici que chez “C.” hancocki. Un foramen supraglénoïde est présent, s’ouvrant latéralement dans la fosse supraglénoïde et médialement dans la partie dorsale de la fosse sous-scapulaire comme chez “C.” hancocki. La lame scapulaire à une diaphyse large, beaucoup plus large que celle d’Alierasaurus. La bordure antéro-médiane de la scapula est bombée par la présence d’un processus scapulaire légèrement arrondi, un caractère partagé avec “C.” hancocki, “C.” bransoni et “Angelosaurus” romeri, mais pas avec C. romeri et Alierasaurus. Deux parties de la fosse glénoïde ont un angle d’environ 130° comme chez “C.” bransoni. Le fémur possède un condyle postérieur occupant une position beaucoup plus distale que le condyle antérieur, contrairement à Ruthenosaurus. La zone poplitée du fémur de Lalieudorhynchus est relativement large avec des sillons robustes, et est beaucoup plus grande et profonde que chez C. romeri. La tête proximale de l’os est plus massive dorso-ventralement que chez C. romeri. Le fémur possède également un grand trochanter interne prononcé et un petit quatrième trochanter dans sa moitié distale, un caractère partagé avec “C.” hancocki, Angelosaurus romeri et A. greeni, et qui diffère de C. romeri, Angelosaurus dolani, Casea broilii, et Ruthenosaurus. La fosse intercondylienne du fémur est très large, inférant un condyle postéro-dorsal étroit, contrairement à C. romeri et Ruthenosaurus. L’astragale est presque aussi large que long contrairement à la plupart des autres caséidés, mais est très similaire à celui de “C.” hancocki. Le métatarse I est robuste et élargi comme chez Alierasaurus. Les phalanges sont courtes et larges. Elles sont plus courtes que chez Alierasaurus[1].
Paléobiologie
L’histologie osseuse d’une côte de Lalieudorhynchus a révélé un os à la structure très spongieuse, au cortex extrêmement fin, et à l’absence de canal médullaire distinct. Ces caractéristiques, également signalés chez d’autres grands caséidés tel Cotylorhynchus, suggèreraient un mode de vie semi aquatique[3] - [1]. Cette hypothèse est toutefois contestée par Kenneth Angielczyk et Christian Kammerer, ainsi que par Robert Reisz et des collègues à partir de données paléontologiques et taphonomiques combinées à l’absence chez ces grands caséidés d’adaptations morphologiques à un mode vie aquatique. Ces auteurs n’apportent toutefois pas encore d’explications alternatives concernant la structure osseuse interne des grands caséidés[4] - [5]. Werneburg et des collègues estiment que Lalieudorhynchus et les grands caséidés en général, aurait pu avoir un mode de vie semi aquatique comparable à celui des hippopotames, passant le plus clair de leur temps dans l’eau, pratiquant une sorte de marche subaquatique plutôt que la nage, et possiblement revenant sur terre pour se nourrir de plantes terrestres. Cependant, les paléontologues ignorent si Lalieudorhynchus se nourrissait de plantes terrestres et/ou aquatiques. Des plantes fossiles associées au squelette de Lalieudorhynchus sont identifiés comme des formes terrestres adaptées à un climat saisonnier sec, alors que les plantes aquatiques ne sont pas présentes. Ces dernières sont toutefois rarement conservées dans les sites du Permien. L’analyse sédimentaire de la localité type de Lalieudorhynchus indiquent l’existence de plusieurs habitats aquatiques potentiels. Des lits argilo-calcaires partiellement limoneux de 15 cm à 1,8 m d’épaisseur proviennent de la sédimentation en suspension dans une masse d’eau stagnante après de fortes inondations. Quelques restes d’arthropodes aquatiques adaptés aux eaux stagnantes temporaires comme les conchostracés et les Triopsidae ont notamment été trouvé dans ces niveaux. Un autre habitat possible correspond à des chenaux de rivière atteignant 3,5 m à 5 m de profondeur dans la formation de La Lieude. La présence d’enracinement à plusieurs niveaux indique que ces chenaux ne devaient pas être remplis d’eau courante toute l’année à cause du climat saisonnier de l’époque. Pendant la saison sèche il est cependant probable que des masses d’eau stagnante aient pu exister dans des chenaux abandonnés, des lacs ou des étangs, permettant la survie d’animaux plus aquatiques comme les dvinosaures tupilakosauridés, dont un spécimen était associé au squelette de Lalieudorhynchus[1].
Découverte et taphonomie
Les premiers restes de Lalieudorhynchus furent découverts en 2001 par Joerg W. Schneider et Frank Körner dans le ruisseau du Salagou lors d’une cartographie géologique de terrain dans le bassin de Lodève. D’autres éléments furent récupérés lors de fouilles effectuées entre 2004 et 2008 totalisant une cinquantaine d’ossements. La gamme de taille des os découverts indique qu’ils proviennent d’un seul individu. La présence d’ossements d’un seul et même individu réparti sporadiquement dans une séquence de 1,4 m d’épaisseur (avec une concentration d’os dans une couche de 40 cm) au sein d’horizons à la lithologie différente pourrait s’expliquer par une décomposition de la carcasse dans une zone végétalisée (comme le suggèrent les nombreux restes de plantes) alternativement exondée et inondée. Les ossements semblent avoir été remaniés[N 4] et redéposés plusieurs fois mais sur une distance très courte et sur une période de temps très brève car les os sont très bien conservés. Ils ne montrent pas d’usure dû au transport par l’eau et ne sont pas fracturés par une exposition prolongée à un climat fortement saisonnier[1].
Répartition géographique et stratigraphique
L’holotype de Lalieudorhynchus provient de la partie supérieure de la Formation de la Lieude, dans le bassin de Lodève situé dans le département de l’Hérault, en Occitanie. Le squelette désarticulé fut découvert dans des strates situées environ 140 m au-dessus de la base de la formation (qui atteint au total 175 m d’épaisseur). Aucune datation radiométrique n’est disponible pour la formation de La Lieude. Un âge Lopingien (= Permien supérieur) lui fut attribué par la biostratigraphie des insectes, ainsi qu’à partir des âges radiométriques et des taux de sédimentations calculés pour la formation du Salagou sous-jacente. Des données magnétostratigraphiques et paléontologiques suggèrent plus probablement un âge Guadalupien (= Permien moyen). La magnétostratigraphie indique que la partie inférieure de la formation de La Lieude ne serait pas plus jeune que l’Inversion d’Illawarra, un évènement géomagnétique global daté du Wordien moyen il y a 266,66 ± 0,76 millions d’années. Par conséquent, cette partie de la formation aurait probablement un âge Roadien supérieur – Wordien inférieur, tandis que la partie supérieure aurait un âge minimal Wordien supérieur – Capitanien inférieur. La présence dans la partie inférieure de la formation de La Lieude de l’ichnogenre Brontopus indique également un âge Guadalupien. Découvert initialement à La Lieude, Brontopus a depuis été trouvé dans la formation d’Abrahamskraal en Afrique du Sud qui est datée radiométriquement du Wordien et du Capitanien. Les auteurs présumés des traces de Brontopus, les thérapsides dinocéphales, concordent également avec un âge Guadalupien de la formation de La Lieude car les ossements de ces animaux, découverts en Afrique australe, en Afrique de l'Est, ainsi qu’en Russie, en Chine, et au Brésil, sont exclusivement connus dans les dépôts du Guadalupien[1]. L’âge Wordien-Capitanien de Lalieudorhynchus fait de lui l’un des derniers caséidés connus. Avec les genres Ennatosaurus et Alierasaurus, il confirme la persistance des caséidés durant le Guadalupien au moins en Europe[1].
Paléoenvironnements
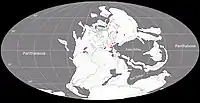
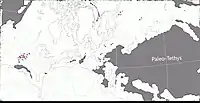
Au Guadalupien, la plupart des terres émergées étaient réunies en un seul supercontinent, la Pangée. Celui-ci avait alors grossièrement la forme d’un C : ses parties nord (la Laurasia) et sud (le Gondwana) étaient reliés à l'ouest mais séparés à l’est par la très vaste mer de Téthys[6]. Un long chapelet de microcontinents, regroupés sous le nom de Cimmérie, divisait la Téthys en deux : la Paleo-Tethys au nord, et la Neo-Tethys au sud[7] Le bassin de Lodève était situé dans la ceinture équatoriale de l’époque, au niveau du 10e parallèle nord, et par rapport aux rivages de la Téthys, se trouvait environ à 400 km à l'intérieur des terres[8] - [9]. Des monts Hercyniens, à la topographie inconnue, séparaient le bassin de Lodève de la Téthys[9]. A cette époque, le climat très humide qui caractérise habituellement le climat équatorial avait été remplacé par une extention du climat tropical plus sec (à deux saisons : sèche et humide) vers les régions proches de l’équateur[8] - [10].
La formation de La Lieude est représentée par des conglomérats, des grès à grain fin à grossier, partiellement caillouteux, des siltites rouge-brun, partiellement argileuses ou sableuses fines, et des claystones limoneux. Ces roches correspondent à des sables, des graviers, et des galets charriés par des rivières, et à des limons fluviatiles de plaines inondables, déposés dans un système fluvial en tresse. Sont également présentes plusieurs strates correspondant à des coulées de laves torrentielles qui témoignent de l’existence de très fortes précipitations pendant la saison humide. La plupart des ossements de Lalieudorhynchus proviennent de ces horizons de laves torrentielles et quelques-uns de coulées de boue. Ces niveaux torrentiels contiennent également d’abondants restes de végétaux représentés par de nombreuses feuilles lancéolées de 5 à 11 cm de longueur appartenant probablement à Plagiozamites, quelques restes de coniférophytes de type Podozamites, des fragments de type Supaia, quelques troncs d’arbres atteignant 2 m de long et 15 à 20 cm de large, et des axes végétaux plus petits[1].
En dehors de Lalieudorhynchus, la partie supérieure de la Formation de la Lieude n’a livrée qu’une colonne vertébrale d’un temnospondyle tupilakosauridé, des arthropodes d’eau douce (conchostracés et triopsidés), des ailes d’insectes (odonata), et des empreintes de pas difficilement déterminables[1]. D’autres éléments de la faune de la formation de La Lieude sont présents sur la dalle à empreintes de La Lieude, située vers la base de la formation, et qui expose de nombreuses pistes de tétrapodes. De nombreuses ichnoespèces[N 5] de tétrapodes ont été nommé en provenance de ce site classé. La révision taxonomique et morphologique de ces empreintes a distingué quatre ichnoespèces valides et identifié leurs auteurs probables: Brontopus giganteus et B. antecursor, qui représentent très probablement des thérapsides dinocéphales (respectivement un Tapinocephalia et un Anteosauridae), Merifontichnus thalerius, qui correspond à des empreintes d’un eureptile Captorhinidae du clade des Moradisaurinae, et Pachypes ollieri, qui appartiendrait à un pareiasauromorpha Nycteroleteridae[11] - [12] - [13].
Phylogénie
L’analyse phylogénétique réalisée par Werneburg et des collègues a identifié Lalieudorhynchus gandi comme l’un des caséidés les plus dérivés et le taxon frère de l’espèce nord-américaine “Cotylorhynchus” hancocki. Ces deux taxons forment un clade caractérisé par la présence d’un hyposphene, ainsi que par la présence et la position du foramen supraglénoïdien. Ce clade forme avec “Cotylorhynchus” bransoni un clade apical caractérisé par des postzygapophyses rapprochées. Cette analyse suggère également que les genres Angelosaurus et Cotylorhynchus (chacun composé de trois espèces) seraient paraphylétiques, les taxons autres que leur espèce type pouvant appartenir à des genres différents[1].
Ci-dessous le cladogramme publié par Werneburg et des collègues en 2022.
Liens externes
Notes et références
Notes
- La prézygapophyse est un processus sur la face antérieure de l’arc neural d’une vertèbre qui s’articule avec la postzygapophyse de la vertèbre située devant elle.
- La postzygapophyse est un processus sur la face postérieure de l’arc neural d’une vertèbre qui s’articule avec la prézygapophyse de la vertèbre située derrière elle.
- L’hyposphene est un élément d’une articulation intervertébrale supplémentaire qui renforçait et stabilisait davantage la colonne vertébrale.
- Un fossile remanié est un fossile qui a été exhumé par l'érosion de la couche sédimentaire dans laquelle il a été déposé à l'origine, et qui est ensuite redéposé dans un dépôt sédimentaire plus jeune.
- Une même ichnoespèce peut avoir été laissée par plusieurs espèces biologiques étroitement apparentées qui partagent une anatomie très similaire de la main et du pied.
Références
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2)
- (en) E.C. Olson, « The family Caseidae », Fieldianna (Geology), vol. 17, , p. 225–349
- (en) M. Lambertz, C.D. Shelton, F. Spindler et S.F. Perry, « A caseian point for the evolution of a diaphragm homologue among the earliest synapsids », Annals of the New York Academy of Sciences, no 1385, , p. 1-18 (DOI 10.1111/nyas.13264)
- (en) K.D. Angielczyk et C.F. Kammerer, « Non-Mammalian synapsids : the deep roots of the mammalian family tree », dans Zachos, F.E. & Asher, R.J., Handbook of Zoology : Mammalian Evolution, Diversity and Systematics, Berlin, de Gruyter, (ISBN 978-3-11-027590-2), p. 138-139
- (en) R.R. Reisz, D. Scott et S.P. Modesto, « Cranial Anatomy of the Caseid Synapsid Cotylorhynchus romeri, a Large Terrestrial Herbivore From the Lower Permian of Oklahoma, U.S.A », Frontiers in Earth Science, no 10, , p. 1-19 (DOI 10.3389/feart.2022.847560)
- (en) S. McLoughlin, « The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism », Australian Journal of Botany, vol. 49, , p. 271-300 (DOI 10.1071/BT00023)
- (en) A.M.C. Şengör, « Tectonics of the Tethysides: orogenic collage development in a collisional setting », Annual Review of Earth and Planetary Sciences, no 15, , p. 214-244 (DOI 10.1146/annurev.ea.15.050187.001241)
- (en) J.W. Schneider, F. Körner, M. Roscher et U. Kroner, « Permian climate development in the northern peri-Tethys area – The Lodève basin, French Massif Central, compared in a European and global context », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 240, , p. 161-183 (DOI 10.1016/j.palaeo.2006.03.057)
- (en) L.A. Michel, N.J. Tabor, I.P. Montañez, M. Schmitz et V.I. Davydov, « Chronostratigraphy and paleoclimatology of the Lodève Basin, France: evidence for a pan-tropical aridification event across the Carboniferous-Permian boundary », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 430, , p. 118-131 (DOI 10.1016/j.palaeo.2015.03.020)
- (en) T.S. Kemp, « The origin and early radiation of the therapsid mammal-like reptiles : a palaeobiological hypothesis », Journal of Evolutionary Biology, vol. 19(4), , p. 1231-1247 (DOI 10.1111/j.1420-9101.2005.01076.x)
- (en) L. Marchetti, H. Klein, M. Buchwitz, A. Ronchi, R.M.H. Smith, W.J. De Klerk, L. Sciscio et G.H. Groenewald, « Permian-Triassic vertebrate footprints from South Africa: Ichnotaxonomy, producers and biostratigraphy through two major faunal crises », Gondwana Research, vol. 72, , p. 139-168 (DOI 10.1016/j.gr.2019.03.009)
- (en) L. Marchetti, « New occurrences of tetrapod ichnotaxa from the Permian Orobic Basin (Northern Italy) and critical discussion of the age of the ichnoassociation », Papers in Palaeontology, vol. 2, no 3, , p. 363-386 (DOI 10.1002/spp2.1045)
- (en) L. Marchetti, S. Voigt, E. Mujal, S.G. Lucas, H. Francischini, J. Fortuny et V.L. Santucci, « Extending the footprint record of Pareiasauromorpha to the Cisuralian : earlier appearance and wider palaeobiogeography of the group », Papers in Palaeontology, no 7(3), , p. 1297-1319 (DOI 10.1002/spp2.1342)