Arisierpeton
Arisierpeton simplex
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Clade | Amniota |
| Sous-classe | Synapsida |
| Clade | † Caseasauria |
| Famille | † Caseidae |
Reisz, 2019
Reisz, 2019
Arisierpeton est un genre éteint de synapside Caseidae ayant vécu durant le Permien inférieur (Artinskien inférieur) dans ce qui est aujourd’hui le sud de l’Oklahoma, États-Unis. Il n’est connu que par des fragments de mâchoires et trois vertèbres isolées retrouvées dans des dépôts de remplissage de fissures karstiques appartenant à un ancien environnement de montagne[1] - [2].
Étymologie
Le nom de genre honore Giuseppe Alberto Arisi qui a identifié le matériel et l’a rendu disponible pour la recherche. L’épithète spécifique se réfère à la morphologie plus simple de la denture marginale comparée à celle de la plupart des autres caséidés permiens[1]
Description
Les éléments connus d’Arisierpeton se composent de trois prémaxillaires droits, un fragment postérieur d’un maxillaire gauche portant trois dents et un fragment de racine, un fragment antérieur de maxillaire droit incluant trois dents et deux fragments de racine, un autre fragment de maxillaire droit avec deux dents complètes, deux fragments de racine et une couronne dentaire non éruptée, un dentaire droit avec sept dents complètes ou presque complètes et 5 dents fragmentaires, et un dentaire gauche portant huit dents plus un petit fragment d’une dent antérieure. Trois vertèbres dorsales postérieures de caséidé retrouvées dans la même poche fossilifère lui sont également attribuées de façon incertaine[1].
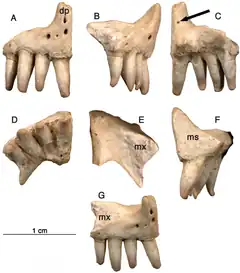
Le prémaxillaire

Bien que les paléontologues ne disposent que de peu de restes d’Arisierpeton, ceux-ci montrent des ressemblances et des différences avec les autres caséidés. Parmi les caractères communs aux autres caséidés, le prémaxillaire possède un processus dorsal penché vers l’avant indiquant que le museau était incliné antérieurement, et la denture marginale n'est pas recourbée, mais légèrement courbée médialement. Contrairement à la condition observée chez la plupart des caséidés, Arisierpeton possède quatre dents prémaxillaires. Les autres membres du groupe n’ont en général que deux ou trois dents prémaxillaires, la première dent étant souvent la plus grosse de la denture marginale. Chez Arisierpeton, les dents prémaxillaires sont toutes de taille similaire. Dans ces deux caractéristiques, nombre et taille des dents, Arisierpeton se distingue facilement des autres caséidés en général et d’Oromycter en particulier, lequel est un autre caséidé provenant de la même localité. Arisierpeton partage avec le caséidé Français Euromycter la présence de quatre dents sur les prémaxillaires. Toutefois, chez Euromycter la première dent du prémaxillaire est plus grande que les autres dents du même os, et est également la plus grande dent de toute la rangée dentaire. La majorité des dents des trois prémaxillaires connus d’Arisierpeton ont leur extrémité distale endommagée. Il est toutefois évident que ces dents se rétrécissent clairement vers la terminaison de la couronne. Cependant, toutes les dents conservées montrent également qu’elles sont quelque peu spatulées vers l’extrémité de la couronne et ont peu de chances d’avoir une extrémité pointue. Leur forme légèrement spatulée montre que Arisierpeton partagent cette condition avec Oromycter, tandis que les caséasaures basaux comme Eocasea et les Eothyrididae ont des dents simples, coniques, légèrement recourbées. De plus, chez les autres caséidés plus dérivés, la morphologie spatulée des dents est la règle, avec des couronnes souvent élargies antéropostérieurement et pourvues à leur extrémité de denticules bien marqués plus ou moins nombreux. Des trois prémaxillaires connus d’Arisierpeton, une seule dent y est complète et montre que la dent est tricuspide, avec deux petites cuspides accessoires qui sont séparées de la pointe centrale de la dent par des crêtes qui courent le long de la couronne. Arisierpeton diffère ainsi d’Oromycter, chez qui les dents spatulées possèdent une extrémité distale large et arrondie, sans cuspides accessoires[1].
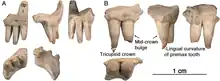
Le maxillaire
Les trois maxillaires sont tous fragmentaires et sont notamment dépourvus du processus antérodorsal. Un seul d’entre eux a conservé la base de ce processus. Celui-ci se distingue par la morphologie de sa bordure antérieure. Chez Oromycter, il n’y a pas d’encoche maxillaire antérieure pour le plateau osseux qui est associée aux narines externes fortement élargies de tous les autres caséidés. Contrairement à Oromycter, chez Arisierpeton, la base préservée de la bordure nasale du processus antérodorsal est large et arrondie, suggérant qu’un plateau maxillaire antérieur peut avoir été présent sur le maxillaire complet. Il n’est pas possible de déterminer avec certitude si ce plateau était présent chez Arisierpeton, mais la forme du maxillaire dans cette région suggère qu’il pourrait y en avoir un. Même si un plateau pleinement développé n’était pas présent chez Arisierpeton, la morphologie du maxillaire représente une condition plus dérivée que celle observée chez Oromycter, chez qui le bord antérieur du processus antérodorsal est une simple crête acérée, et le plateau osseux de la narine externe est limitée à l’os lacrymal.
Les dents maxillaires présentent la morphologie typique des caséidés en ayant des couronnes incurvées lingualement, avec peu ou pas de recourbure. La dent maxillaire la mieux conservée a une morphologie superficielle similaire à celle du prémaxillaire, avec des cannelures verticales à la surface de l’émail. La plupart des dents ont toutefois perdu le bout de leur couronne, ce qui rend difficile de déterminer si elles étaient également tricuspides. Cependant, la disposition des cannelures suggère qu’il y avait également un cône central et éventuellement deux cuspides accessoires, ou au moins des cuspides accessoires naissantes[1] - [3].
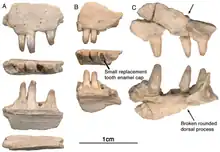
Le dentaire
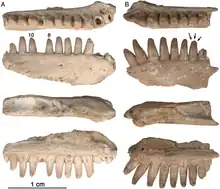
Les dentaires d’Arisierpeton présentent la même configuration générale que celle des autres caséidés en ayant un bord dorsal se courbant ventralement près de la symphyse et formant un angle aigu avec le bord ventral de l’os. Il en résulte un os dentaire sensiblement plus mince près de la symphyse que dans le reste de l’os. Les dents de ces dentaires sont identiques à celles des maxillaires. Comme chez tous les caséidés, les dents antérieures du dentaire se penchent vers l’avant. Les trois premières dents inclinées vers l’avant sont les plus grandes de la série. L’apex de chaque dent porte des carènes antérieures et postérieures, avec des cuspides accessoires peu marquées associées aux cannelures antérieures du cône central de la dent. Ces dents semblent également avoir un cône central légèrement incliné vers l’arrière. Les dents du dentaire sont plus fines que celles d’Oromycter et ont des épaules linguales plus modestes à la base de la couronne[1].
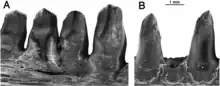
Dans l’ensemble, il semble que la denture d’Arisierpeton montre quelques modifications dans la forme des dents et le contour de la couronne par rapport à l’état amniote primitif observé chez le caséidé basal Eocasea et chez les caséasaures Eothyrididae. Les dents présentent peu ou pas de recourbure, mais ont plutôt une courbure médiale ou linguale apicale. Les couronnes, lorsqu’elles sont conservées, présentent des cannelures et des carènes occasionnelles, qui sont parfois suffisamment bien développées pour la formation d’un terminus tricuspide, rappelant quelque peu la condition observée chez Cotylorhynchus. Cependant, le type de renflement du côté lingual de la dent sous la couronne observée chez Cotylorhynchus et les caséidés géologiquement plus jeunes n’est développé que modestement chez Arisierpeton[1].
Les vertèbres
Une série de trois vertèbres dorsales postérieures incomplètes a été trouvée dans la même poche fossilifère que les fragments de mâchoires, mais leur association avec l’holotype et d’autres spécimens référés est provisoire. Elles sont principalement représentées par leur centra, une ayant toutefois conservée son épine neurale. Elles appartiennent clairement à un petit caséidé. Cependant, leur affectation à Arisierpeton est provisoire et basée sur leur cooccurrence et sur leur petite taille, et il n’y a pas de caractéristiques diagnostiques reconnaissables sur les centra en dessous du niveau de la famille[1].
Phylogénie
Le matériel disponible d’Arisierpeton est trop fragmentaire pour permettre une analyse phylogénétique. Il semble toutefois être un caséidé basal tout comme Oromycter. À en juger par les caractères de sa denture et de son maxillaire, Arisierpeton semble un peu plus dérivé que Oromycter mais plus basal que les autres caséidés[1].
Age et paléoenvironnement
Tous les fossiles d’Arisierpeton proviennent de la carrière de Dolese Brothers à Richards Spur, dans le Comté de Comanche, en Oklahoma[1]. Ce site très particulier, exploité depuis plus d’un siècle, représente un remplissage de sédiments fossilifères du Permien inférieur dans une série de fissures karstiques de calcaire et de dolomie de l’Ordovicien[2] - [1]. Les spéléothèmes intimement associés au site indiquent que Richards Spur est un système de grottes, suggérant un environnement de conservation distinct de ceux des localités plus typique des plaines deltaïques et fluviales du Permien inférieur. La composition faunique unique de Richards Spur, et l’absence de taxons franchement aquatiques à cet endroit, ont conduit à suggérer qu’il représente une localité de montagne. Ce réseau de fissures contient l’une des faunes terrestres du Paléozoïque la plus diversifiée actuellement connue, avec une quarantaine de taxons de tétrapodes (dont plus d’une trentaine ont été nommés) de tailles petites à moyennes. L’assemblage conservé dans ces grottes est surtout composé de parareptiles, d’eureptiles, et de temnospondyles dissorophoïdes, tandis que les restes de synapsides y sont beaucoup plus rares (incluant une espèce indéterminée de Dimetrodon[4])[2]. L’analyse isotopique du rapport U-Pb de plusieurs spéléothèmes isolés, considérés comme des stalactites en raison de leur forme, a donné des âges de 289,2 ± 0,68 Ma, 286,7 ± 2,9 Ma, et 286,2 ± 0,2 Ma, correspondant à la partie inférieure de l’étage Artinskien du Permien inférieur. Ces datations indiquent que la durée minimale et maximale possible de la période de formation active des grottes était respectivement de 2,1 Ma et 6,1 Ma. La présence de spéléothèmes indique également que les grottes étaient hydrologiquement actives. De plus, de nombreux spécimens fossiles se trouvent enfermés dans des blocs de calcite grossièrement cristalline, qui doivent avoir précipité autour des restes d’animaux submergés. Au Permien inférieur, Richards Spur était situé dans le sud-ouest de la partie équatoriale de la Pangée. D’après les informations climatiques obtenues à partir des spéléothèmes, Richards Spur aurait connu des conditions météorologiques de mousson, et donc une forte saisonnalité, alternant entre périodes d’aridité et périodes de fortes précipitations. Il s’agit d’un paléoclimat comparable aux conditions qui ont été proposées pour les localités de plaines du sud-ouest équatorial de la Pangée, via l’analyse des paléosols, ainsi que des plantes fossiles[2].
Formation des grottes et origines des fossiles qu’elles contiennent
Les fissures de Richards Spur sont localisées dans le calcaire d’Arbuckle, lequel s’est déposé durant l’Ordovicien. Par la suite, le calcaire a été soulevé et est devenu subaérien pendant le Carbonifère supérieur et le Permien. Ce soulèvement a redressé les strates calcaires originellement horizontale dans une position quasi verticale. Une condition qui a facilité l’infiltration de l’eau le long des plans de stratification, et la dissolution du calcaire ordovicien, pour finalement développé un système de grottes. L’assemblage de vertébrés retrouvés dans le remplissage de ces grottes, indique que ces dernières étaient ouvertes vers la surface pendant le Permien inférieur. Au Crétacé, le calcaire d'Arbuckle était submergé par la Voie maritime intérieure de l’Ouest et une formation sableuse recouvrait le calcaire. Après le retrait de cette mer, le calcaire d’Arbuckle a été érodé, exhumant les fissures calcaires et leur remplissage depuis au moins le Pléistocène. Les passages qui composent le système de grottes sont généralement petits, mesurant environ 40 à 60 cm de large, et sont principalement parallèles aux strates verticales du calcaire ordovicien. De nombreux passages se ramifient et rencontrent d'autres parties du système de grottes, et de grands passages occasionnels se produisent. Bien que la profondeur totale des grottes ne soit pas connue, seul le remplissage des 25 premiers mètres contient des fossiles. Les ossements retrouvés dans ces grottes sont souvent désarticulés, bien que les restes articulés n’y soient pas rares. Cela implique au moins deux scénarios sur la façon dont les fossiles s’y sont déposés. De nombreux animaux morts à l'extérieur des grottes ont probablement été désarticulés à la surface, puis emportés par les pluies jusque dans les conduits verticaux des grottes qui étaient probablement des points bas naturels à la surface. Le deuxième mode de dépôt implique que des animaux entiers pénètrent dans les grottes. Pendant les périodes de pluie de mousson, les animaux morts à la surface auraient pu être emportés dans les grottes avant d’avoir été désarticulés par la décomposition et/ou la prédation. Alternativement, les grottes de Richards Spur ont pu jouer aussi le rôle de pièges naturels. Les animaux vivant autour du système de grottes y tombant à l’occasion ou, dans certains cas, étant emportés vivants[2].
La majorité des fossiles de Richards Spur ont été imprégnés d’hydrocarbures provenant des champs pétrolifères sous-jacents du schiste de Woodford. Par conséquent, ces os sont de couleur brun foncé à noir et sont très faciles à voir dans les sédiments de la grotte. A Richards Spur, les fossiles les plus communs ainsi préservés appartiennent au petit Amphibamidae Doleserpeton et au petit Captorhinidae Captorhinus aguti. On y a trouvé aussi le petit caséidé Oromycter. Ce type de fossiles a été collecté pendant plus de 90 ans. A la fin du 20e siècle, Richards Spur a livré une nouvelle grotte dont les fossiles sont dépourvus d’imprégnation d’hydrocarbures. Ainsi, contrairement à la grande majorité des fossiles de Richards Spur de couleur brun foncé ou noir, les fossiles de cette nouvelle grotte étaient de couleur crème, presque de couleur blanche. La faune préservée dans ce système de grotte comprend de nombreux restes désarticulés du Trematopidae Acheloma et du grand captorhinide Captorhinus magnus. Les spécimens d’Arisierpeton sont dérivés de cet assemblage d'os blancs aux couleurs distinctes. Malgré des différences dans la composition des taxons les plus abondants, il existe des similitudes entre les deux types de communautés, le Dissorophidae Cacops et le captorhinide Captorhinus aguti étant présents dans les deux assemblages, mais étant plus rares dans l’assemblage "os blanc". La seule différence majeure est la rareté apparente du petit amphibamide Doleserpeton dans l'assemblage "os blanc". Le caséidé Oromycter est connu de l’assemblage "os noirs", tandis que Arisierpeton est connu de l’assemblage "os blancs". Deux caséidés sont donc présents à Richards Spur, mais ceux-ci proviennent de deux remplissages de grottes qui étaient probablement séparés spatialement d'au moins 300 m. Il n’est pas possible de déterminer s’ils étaient d'âge contemporain ou séparés stratigraphiquement ou temporellement, car le remplissage des grottes est éjecté lors des activités d’extraction de la carrière. Ce qui rend impossible de savoir exactement de quelles parties des grottes les fossiles et les spéléothèmes sont obtenus[1].
Liens externes
- Ressources relatives au vivant :
Notes et références
- (en) R.R. Reisz, « A small caseid synapsid, Arisierpeton simplex gen. et sp. nov., from the early Permian of Oklahoma, with a discussion of synapsid diversity at the classic Richards Spur locality », PeerJ 7:e6615, , p. 1-20 (DOI 10.7717/peerj.6615, lire en ligne)
- (en) M.J. MacDougall, N.J. Tabor, J. Woodhead, A.R. Daoust et R.R. Reisz, « The unique preservational environment of the Early Permian (Cisuralian) fossiliferous cave deposits of the Richards Spur locality, Oklahoma », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 475, , p. 1-11 (DOI 10.1016/j.palaeo.2017.02.019)
- (en) R.R. Reisz, « Oromycter, a new caseid from the lower Permian of Oklahoma », Journal of Vertebrate Paleontology, vol. 25(4), , p. 905-910 (DOI 10.1671/0272-4634(2005)025[0905:OANCFT]2.0.CO;2)
- (en) K.S. Brink, M.J. MacDougall et R.R. Reisz, « Dimetrodon (Synapsida: Sphenacodontidae) from the cave system at Richards Spur, OK, USA, and a comparison of Early Permian-aged vertebrate paleoassemblages », TheScience of Nature, vol. 106(2), (DOI 10.1007/s00114-018-1598-1)