Eocasea
Eocasea martini
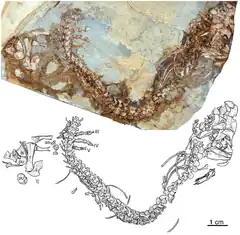
Eocasea est un genre éteint de synapsides Caseidae ayant vécu au Carbonifère supérieur dans ce qui est aujourd’hui le Kansas, États-Unis. Le genre n’est représenté que par son espèce type, Eocasea martini, qui fut nommée en 2014 par Robert Reisz et Jorg Fröbisch. L’holotype et unique spécimen connu est représenté par un squelette incomplet incluant un crâne partiel, une colonne vertébrale articulée presque complète et un membre postérieur appartenant à un individu juvénile. Il est le premier caseidé datant du Carbonifère à avoir été décrit. Les caractères de sa denture et de son squelette suggèrent qu’il était carnivore, ou plus probablement insectivore étant donné sa petite taille. Eocasea démontre ainsi que les grands caseidés herbivores du Permien ont évolué à partir d’espèces prédatrices de petites tailles[1]. Toutefois, il n’est pas impossible que la denture d’insectivore d’Eocasea soit un caractère juvénile qui aurait été remplacé à l’âge adulte par une denture d’omnivore, comme cela a été mis en évidence chez le caséidé Martensius du Permien d’Allemagne[2].
Étymologie
Le nom générique se compose du Grec eos l’aube, et de Casea et fait référence au fait qu'il s'agit du plus ancien membre connu du clade. L'épithète spécifique est un hommage au Dr. Larry Dean Martin, le collecteur original du spécimen holotypique[1].
Description
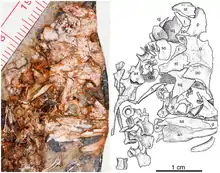
L’holotype est un individu juvénile incluant le tiers postérieur du crâne et de la mandibule droite, une colonne vertébrale presque complète (comprenant les vertèbres cervicales, dorsales, sacrées, et une partie des vertèbres caudales), la ceinture pelvienne, et le membre postérieur droit presque complet. La longueur de l’animal est de 12,5 cm queue non comprise. Bien que fragmentaire et très écrasé, le crâne montre plusieurs caractères typiques des caséidés tels qu’une grande fosse temporale latérale bordée par un squamosal postérieurement étroit, un quadratojugal contribuant au bord ventral de la fosse temporale, un grand postorbitaire avec une large surface dorsale contribuant significativement au toit crânien, et la présence sur le toit crânien de petites dépressions arrondies distribuées de façons aléatoires. Le fragment préservé du maxillaire portait à l’origine quelques dents mais celles-ci furent détruites lors du dégagement à l’acide nécessaire à la libération du spécimen de sa gangue calcaire. Le spécimen non préparé montrait la présence d’une denture conique simple, sans aucun signe de cuspides sur la couronne, ni même le type d’expansion mésio-distale de la couronne présente chez Oromycter. La mâchoire inférieure n'est que partiellement conservée et exposée en vue latérale. La partie postérieure du dentaire est exposée en vue latérale, son bord dorsal étant recouvert par le maxillaire. Le dentaire montre un long processus postérieur qui s’étend le long du bord dorsal de la mâchoire inférieure et couvre une partie du surangulaire dans la région de la faible éminence coronoïde. Chez les autres caséidés le bord dorsal de l’éminence coronoïde est occupé par le coronoïde et le surangulaire. L’angulaire, bien qu’un peu écrasé, semble être plus haut que le surangulaire, une condition qui rappelle plus Eothyris que Cotylorhynchus. L’os articulaire est un petit os à l’extrémité postérieure de la mandibule, et n’a pas de processus rétroarticulaire, comme chez tous les autres caséidés[1].
Le squelette postcrânien est caractérisé par la présence de 27 vertèbres présacrées et deux vertèbres sacrées, une condition très proche de la condition plésiomorphe (26 présacrées et deux sacrées) pour les synapsides et tous les amniotes. Les autres caséidés ont un nombre plus réduit de vertèbres présacrées (24–26) et un nombre plus élevé de vertèbres sacrées (3–4). Tout le long de la partie cervicale et présacrale de la colonne, les vertèbres ont de simples épines neurales en forme de lame. Elles sont bien développées antéropostérieurement, mais courtes dorsoventralement. Les vertèbres caudales ont des épines neurales très réduites. Toutes les vertèbres cervicales, dorsales et présacrales ont des processus transversaux relativement courts, contrastant fortement avec les autres caséidés, où les processus transversaux s’étendent bien latéralement. Contrairement aux grands caséidés herbivores, les côtes dorsales d’Eocasea ne sont pas épaissies et ne forment pas une cage thoracique expansée dorsolatéralement en forme de tonneau. Chez la plupart des caséidés, les côtes lombaires ont tendance à fusionner avec les vertèbres et restent massives et grandes, de section similaire au reste des côtes dorsales, s'étendant loin latéralement. Chez Eocasea, les trois dernières côtes, celles juste avant le sacrum, sont petites et plus délicates que les autres côtes dorsales, une condition similaire à celle observée chez les synapsides non herbivores. Contrairement aux autres caséidés, qui possède trois ou quatre côtes sacrées, Eocasea n’a que deux côtes sacrées, la première étant significativement plus large distalement que la seconde. La ceinture scapulaire et la patte antérieure ne sont pas conservées, mais la ceinture pelvienne et le membre postérieur droit sont présents. L'ilium a la morphologie typique des caséidés avec une lame iliaque haute avec des rainures verticales et des crêtes distales. La lame est centrée directement au-dessus de l'acétabulum, avec des processus antérieurs modestes et légèrement plus grands, similaires à ceux observés chez Ruthenosaurus et la plupart des autres caséidés. Comme chez les autres caséidés, le fémur, le tibia et la fibula sont massivement construits, mais ne montrent aucun caractère particulier, peut être en raison de la nature juvénile du spécimen. Le pied possède un 1er métatarsien long et massif, de longueur presque égale aux 4e et 5e de la série. Les phalanges conservées ont une longueur approximativement égale, et les phalanges terminales portant des griffes sont grandes, avec un tubercule adducteur massif. La formule phalangienne[N 1] est 2-3-4-5-4 comme celle de Casea broilii et de Martensius, représentant la condition plésiomorphe des amniotes. Tous les autres caséidés dont la formule phalangienne est connue montre une réduction du nombre de phalanges dans le pied[1] - [2].
Répartition géographique et stratigraphique

L’holotype d’Eocasea martini fut découvert dans la carrière Hamilton, près de Hamilton au Kansas, dans le comté de Greenwood. Il provient de la Formation de Calhoun Shale du groupe de Shawnee, datée de l’étage local Virgilien[1], lequel correspond à l’étage Gzhélien de l’échelle stratigraphique internationale[3]. La carrière Hamilton est connue pour être un Konservat-Lagerstätte (un dépôt sédimentaire avec une extraordinaire préservation des fossiles) correspondant à une paléovallée influencée par les marées, où le dépôt rapide de sédiments a préservé une flore et une faune inhabituellement diversifiées. La faune de vertébrés est essentiellement aquatique et est dominée par des centaines de poissons acanthodiens superbement préservés et de nombreux amphibiens dissorophoïdes. Seuls quelques amniotes, tous de petits carnivores terrestres, ont été retrouvés. Outre Eocasea, il s’agit de l’araeoscélidien Spinoaequalis, du Varanopidae Archaeovenator, et du Captorhinidae Euconcordia. Les restes de tous ces amniotes sont représentés par de petits squelettes ne montrant aucun signe de prédation ou de nécrophagie, mais quelques signes de transport par l’eau. Ces spécimens ont probablement été emportés dans la paléovallée[1].
Phylogénie
En 2020, une analyse phylogénétique publiée par Berman et des collègues a identifiée Eocasea comme le plus basal des Caseidae[2].
Ci-dessous, les deux cladogrammes des Caseidae publié par Berman et des collègues en 2020[2].
Par contre, il n'est pas présent dans le cladogramme suivant publié en 2022. Ci-dessous, le cladogramme publié par Werneburg et des collègues en 2022[4].
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- La formule phalangienne correspond au nombre de phalanges constituant chaque doigt des mains et des pieds des tétrapodes. Elle s’énumère en partant du doigt I (correspondant chez l’homme au pouce et au gros orteil) jusqu’au doigt V (l’équivalent de l’auriculaire et du petit orteil).
Références
- (en) R.R. Reisz et J. Fröbisch, « Oldest caseid synapsid from the late Pennsylvanian of Kansas, and the evolution of herbivory in terrestrial vertebrates », PLoS One, vol. 9 (4): e94518, (PMID 24739998, PMCID 3989228, DOI 10.1371/journal.pone.0094518)
- (en) D.S. Berman, H.C. Maddin, A.C. Henrici, S.S. Sumida, D. Scott et R.R. Reisz, « New primitive Caseid (Synapsida, Caseasauria) from the Early Permian of Germany », Annals of Carnegie Museum, vol. 86(1), , p. 43-75 (DOI 10.2992/007.086.0103)
- (en) V. Davydov, D. Korn et M.D. Schmitz, « The Carboniferous Period », dans Gradstein, FM., Ogg, J.G., & Schmitz, M.B., Ogg, G.M., The Geologic Time Scale 2012, Elsevier BV, (ISBN 978-0-44-459390-0), p. 605
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2).