Angelosaurus
Angelosaurus est un genre éteint de synapsides Caseidae herbivore ayant vécu à la fin du Permien inférieur (Kungurien) et au début du Permien moyen (Roadien) dans ce qui est aujourd’hui le Texas et l’Oklahoma aux États-Unis. Comme les autres caséidés herbivores il avait une petite tête, un grand corps en forme de tonneau, une longue queue, et des membres massifs. Angelosaurus diffère des autres caséidés par l’extrême massivité de ses os, en particulier ceux des membres, lesquels montrent un fort développement des crêtes, des processus, et des rugosités pour la fixation des muscles et des tendons. Par rapport à la taille de son corps, les membres d’Angelosaurus étaient plus courts et plus larges que ceux des autres caséidés. Les phalanges unguéales ressemblaient davantage à des sabots plutôt qu’à des griffes. Les quelques éléments crâniens connus montrent que le crâne était court et plus robuste que celui des autres représentants du groupe. Angelosaurus se distingue également par ses dents bulbeuses aux couronnes plus courtes et plus larges que celles des autres caséidés. Leur morphologie et le fort taux d’usure qu’elles présentent suggère un régime alimentaire différent de celui des autres grands caséidés herbivores, et devait être à base de plantes particulièrement coriaces. Une étude publiée en 2022 suggère que le genre pourrait être paraphylétique, deux des trois espèces pouvant appartenir à un ou des genres distincts.

Étymologie
Le nom de genre Angelosaurus réfère à la formation de San Angelo au Texas, où cet animal fut découvert pour la première fois. Il se complète du suffixe saurus signifiant lézard.
Espèces
Angelosaurus n’est connu que par des squelettes incomplets et des ossements isolés. Le genre est représenté par trois espèces, la plus grande, Angelosaurus greeni, atteignant une taille comparable à celle de Cotylorhynchus romeri[1]. A. romeri était l’espèce la plus petite et A. dolani était de taille intermédiaire. Cependant, la différence de taille n’est pas un critère taxonomique pertinent car le nombre de spécimens connus est trop limité pour connaître la taille maximale atteinte par chaque espèce[2]. Ces trois espèces diffèrent essentiellement par des caractéristiques de l’humérus, du fémur, et du bassin, qui sont les seuls éléments disponibles permettant des comparaisons. Chez l’espèce type Angelosaurus dolani, l’humérus est très massif, aussi long que large avec une diaphyse quasi inexistante. Chez A. romeri, l’humérus est moins massif avec une diaphyse très courte mais bien distincte. Chez A. romeri et A. dolani, le trochanter interne du fémur est situé plus ou moins à mi-chemin entre les extrémités proximale et distale de la fosse intertrochantérienne alors que chez A. greeni il est situé près de l’extrémité distale de la fosse. A. romeri se distingue par la proéminence du quatrième trochanter. Chez A. romeri et A. greeni, le condyle externe du fémur est le plus grand des deux et est bien séparé du condyle interne. Chez A. dolani c’est le condyle interne qui est le plus développé mais la séparation avec le condyle externe est moins nette, probablement en raison de la présence d’une couverture cartilagineuse étendue. Le bassin d’A. dolani est caractérisé à la fois par la massivité des connexions vertébrales et la forte fusion des côtes sacrales formant une seule plaque distalement, mais aussi par la faible ossification de la lame dorsale de l’ilion et de la symphyse pubienne. Chez A. romeri il n’y a pratiquement pas de fusion des côtes sacrales. Le sacrum d’A. dolani compte trois vertèbres dont seules les deux premières sont fusionnées. Chez A. romeri, la symphyse pubienne est fortement ossifiée et le sacrum compte quatre vertèbres dont trois d’entre elles au moins sont fusionnées chez l’holotype[3] - [2] - [1]. Une analyse cladistique réalisée par Werneburg et des collègues indique cependant que les espèces A. greeni et A. romeri pourraient ne pas appartenir au genre Angelosaurus, tous ces taxons nécessitant une redescription détaillée[4].
Angelosaurus dolani
L’espèce type Angelosaurus dolani fut érigée en 1953 par Everett C. Olson et James R. Beerbower à partir d’un squelette partiel (l’holotype FMNH UR 149) trouvé dans les mudstones rouges constituant la partie centrale de la formation de San Angelo (Groupe de Pease River), dans le Comté de Knox au Texas. Cette espèce fut nommée en l’honneur de Wayne Dolan, dont la coopération a permis de poursuivre les travaux sur le gisement de Little Crotton Creek, d’où provient l’holotype, situé sur le terrain du ranch MacFayden, près de la ville de Benjamin[3]. Le squelette d’A. dolani était en grande partie articulé. Les trois premières vertèbres présacrées, le sacrum, et 24 vertèbres caudales étant en connexion anatomique, de même que le bassin, les fémurs, le tibia et la fibula gauche, et la majeure partie du pied gauche. Quatre autres vertèbres, l’humérus droit, des éléments de la main droite, des débris de côtes, deux fragments de dentaire et de maxillaire mal conservés et aux dents cassées, étaient dispersés antérieurement au bassin[3]. Quelques vertèbres d’un second spécimen furent également signalées du même gisement[2] - [1], à moins d’une centaine de mètres du premier site[5].
A. dolani est caractérisé par un squelette lourd et massif, aux membres plus courts et plus larges que ceux des autres caséidés. L’humérus est l’un des éléments les plus distinctif du squelette par ses proportions extrêmement massives. Il n’y a pratiquement pas de diaphyse, et la largeur de l’extrémité proximale est presque égale à la longueur totale de l’os. La crête delto-pectorale est très développée et porte un processus proéminent, noueux et rugueux pour l'insertion d'une partie de la musculature pectorale. Le long de la marge antérieure de la partie proximale de l'humérus, il y a une forte crête pour l'insertion du muscle deltoïde. Le condyle radial de l’humérus se trouve loin en avant sur la surface ventrale et distale, et ses proportions suggèrent que la tête du radius était massive. Le fémur est très massif avec une extrémité proximale profonde et lourde. Le trochanter interne est exceptionnellement solide et a une surface rugueuse. De façon caractéristique, il s’étend très près de l’extrémité proximale de la fosse intertrochantérienne (une large surface concave située sur la face ventrale de l’extrémité proximale du fémur et où s’insérait le muscle puboischiofemoralis externus). Le quatrième trochanter, en revanche, est faible. Les condyles distaux du fémur sont volumineux mais pas fortement différenciés, probablement en raison de la présence d'une calotte cartilagineuse. Une forte crête passe proximalement de la fosse intercondylienne le long de la marge dorsale du fémur et continue sur la zone où la partie fémoro-tibiale du triceps a vraisemblablement pris son origine. Tout comme le fémur, le tibia et la fibula sont caractérisés par leurs extrémités formées d’os non fini, suggérant la présence d’une couverture cartilagineuse importante. Cette particularité, qui d’ordinaire est un signe d’immaturité squelettique, est étonnante car l’holotype d’A. dolani appartient à un animal adulte comme l’indique l’ossification complète de diverses autres parties du squelette[3]. Du pied gauche, relativement grand par rapport à la taille du corps, les tarses furent retrouvés articulés mais les métatarses et les phalanges étaient quelque peu déplacés. De sorte qu'Olson et Beerbower eurent quelques difficultés à déterminer la formule phalangienne[3]. Après réexamen du matériel découvert, Olson évalua plus tard que la formule phalangienne du pied était 2-2-2-3-3[1]. Les phalanges unguéales se distinguent de celles des autres caséidés en étant courtes, larges, et lisses, et ressemblent à des sabots plutôt qu’à des griffes[1] - [6]. Quelques éléments de la main droite montrent que les phalanges étaient larges et courtes. Le bassin est caractérisé par un pubis et un ischion formant une large plaque aplatie, avec une symphyse faible sur la plus grande partie de sa longueur. L’ilium, qui est court et large, montre quelques indications de l’élévation dorsale caractéristique des caséidés, mais le sommet semble incomplet, peut-être cartilagineux. L’acetabulum est proportionnellement très large et profond. Le sacrum est composé de trois vertèbres, la première et la seconde étant soudées. Les extrémités distales des trois côtes sacrées sont fusionnées pour former une large plaque, qui s'insère sur l'ilium. Les vertèbres thoraciques possèdent des apophyses transverses relativement longues et il semble y avoir eu une petite facette ventrale, indiquant que les côtes étaient à double tête. Les épines neurales, bien que mal conservées, semblent avoir été plutôt courtes. Deux fragments de mâchoires supérieure et inférieure indiquent que le crâne était plutôt petit par rapport à la taille du corps, comme c’est le cas chez les autres caséidés dérivés. Les deux morceaux de dentaire et de maxillaire portent quelques dents qui, bien que cassées, avaient une section ovoïde et étaient assez espacées, indiquant que leur nombre total devait être réduit[3].
Angelosaurus greeni
Angelosaurus greeni fut nommée en 1962 par Everett C. Olson à partir d’un squelette très fragmentaire et divers éléments isolés trouvés dans les mudstones rouges du sommet de la formation de San Angelo[N 1], dans le Comté de Knox au Texas. L’espèce honore J. Green du ranch Alexander, dont la coopération et l’aide ont permis pendant plusieurs années d'explorer et de collecter en profondeur les affleurements fossilifères des terres du ranch Alexander[2]. Il s’agit de l’espèce la plus grande du genre avec des dimensions comparable à celle du caséidé Cotylorhynchus romeri. Elle est aussi l’espèce la moins bien connue. Les vestiges retrouvés étaient fragmentaires, en grande partie à cause d'une importante décomposition chimique après l'enfouissement. Chez l’holotype, cette décomposition a altéré et détruit de nombreux éléments d’un squelette originellement plus complet comme l’indiquait la présence de nombreux fragments difficilement identifiable, mais aussi la présence sur le sol du contour indistinct de certains os entièrement dissous[2] - [5]. L’holotype (FMNH UR 257) est composé d’un fémur, de quelques vertèbres, et des fragments de côtes. Les paratypes incluent un morceau d’extrémité distale d’un humérus, des fragments de bassin, des éléments distaux des membres, et des fragments indéterminés, et un humérus aplati et à l’extrémité distale mal conservée, tous provenant du même horizon général et de la même localité que le type[2] - [1].
Cette espèce est principalement caractérisée par la morphologie de son fémur. L’emplacement du trochanter interne est très caractéristique en étant situé près de l'extrémité distale de la fosse intertrochantérienne. Alors qu’il est situé près de la partie médiane ou proximale de cette fosse chez les deux autres espèces. Ce trochanter interne est apparemment fusionné avec le quatrième trochanter. Il n’y a pas de crête de l’adducteur. Le condyle distal antérieur (interne) est petit et est nettement séparé du postérieur par une rainure profonde. Enfin, la diaphyse du fémur est très courte[2].
Angelosaurus romeri
Angelosaurus romeri fut nommée en 1962 par Everett C. Olson et Herbert Barghusen à partir d’un squelette partiel et de nombreux autres éléments isolés trouvés dans la carrière d’Oméga, dans le Comté de Kingfisher en Oklahoma. Ses restes furent initialement décrits comme provenant de la partie centrale de la formation de Flowerpot[5]. Plus tard, Olson a corrigé cette attribution en précisant que ces restes appartiennent à une langue de la formation de Chickasha (Groupe d’El Reno) dont les dépôts s’imbriquent par endroit avec ceux des formations de Flowerpot, de Blaine, et de Dog Creek[7] - [1]. Bien qu’Olson et Barghusen ne donne aucune information sur le nom de l’espèce, celui-ci fait probablement référence au paléontologue américain Alfred Sherwood Romer. Une autre espèce de caséidé portait déjà son nom, Cotylorhynchus romeri. A. romeri est l’espèce du genre la mieux connue, et la seule pour laquelle on dispose d’éléments de diverses parties du crâne. C’est la plus petite espèce du genre et elle diffère de ces cousines par un développement modéré de la massivité des membres et une faible augmentation de la robustesse des vertèbres, lesquelles ne peuvent être distinguées qu’avec difficulté de celles de Cotylorhynchus[1].
L’holotype (FMNH UR 827) est composé du bassin, du fémur droit, de 16 vertèbres présacrées dont cinq avec leurs côtes associées, 3 sacrées, et 4 caudales dont deux associées avec des chevrons. Sont également connues deux vertèbres sacrées et 2 présacrées formant une série articulée (FMNH UR 828), le côté droit d’un bassin dépourvu de la plus grande partie de l’ischion (FMNH UR 844), un pubis droit (FMNH UR 845), une interclavicule (FMNH UR 846), deux côtes dorsale antérieure (FMNH UR 847 et 850), deux côtes dorsale médiane (FMNH UR 848 et 849), une côte dorsale postérieure (FMNH UR 851), une côte cervicale (FMNH UR 852), quatre vertèbres caudales antérieures (FMNH UR 853), et un fragment de prémaxillaire avec deux dents (FMNH UR 854)[5].
Les fouilles ultérieures dans la carrière d’Oméga ont permis de découvrir de nombreux ossements supplémentaires, dont plusieurs éléments inédits du squelette. Ce matériel additionnel inclus un squelette partiel composé du sacrum, 4 vertèbres sacrées, 3 présacrées, 18 caudales et des os des orteils (FMNH UR 904), un scapulocoracoïde droit et un humérus droit (FMNH UR 907), 20 vertèbres présacrées (FMNH UR 908), un scapulocoracoïde gauche (FMNH UR 909), un tibia mal conservé (FMNH UR 911, attribué avec doute à ce taxon), 4 os des orteils (FMNH UR 914), 3 vertèbres présacrées avec des côtes (FMNH UR 916), un fémur gauche de grande taille (FMNH UR 917), un fragment de maxillaire avec 2 dents (FMNH UR 926), une boîte crânienne et une partie du palais (FMNH UR 927), une côte de grande taille (FMNH UR 928), un ptérygoïde (FMNH UR 931), une côte dorsale antérieure (FMNH UR 932), une clavicule (FMNH UR 933), la troisième côte présacrée (FMNH UR 940), une côte dorsale antérieure (FMNH UR 941), une dent isolée (FMNH UR 942), une paire de mâchoires inférieures articulées ayant conservées leurs rangées de dents (FMNH UR 943), 2 côtes dorsales médiane (FMNH UR 944), plusieurs côtes dorsales antérieures (FMNH UR 945), 20 vertèbres caudales avec des côtes sur les plus antérieures (FMNH UR 971), un pubis (FMNH UR 978), ilia et ischia d’individus juvéniles (FMNH UR 979), et un bassin gauche et droit (FMNH UR 980)[7] - [1].
Aucun crâne complet d’Angelosaurus n’a encore été découvert. L’espèce A. romeri fournit la meilleure image de la morphologie crânienne chez ce genre. Le crâne est représenté par un prémaxillaire incomplet portant deux dents, un fragment de maxillaire portant deux dents, une boîte crânienne, un ptérygoïde, et une paire de mâchoires inférieures portant une denture quasi complète. Tous ces éléments sont cohérents entre eux en indiquant un crâne assez court, robuste et lourd, avec des dents fortes et courtes. L’apparence du crâne en vue latérale n’est pas bien connue mais le prémaxillaire montre que les narines externes étaient grandes, et le fragment de maxillaire montre l'existence d'une grande orbite. Ces deux fragments suggèrent que les éléments du crâne étaient assez massifs par rapport à ceux de Cotylorhynchus de taille comparable. Le crâne d’A. romeri devait ressembler à une version un peu plus solide et robuste du schéma de base des autres caséidés dérivés. La paire de mâchoires inférieures montre que le dentaire est court et lourd avec une symphyse forte. Il y a un plateau moyennement fort et large à la marge ventrale de la symphyse[1].
La denture d’A. romeri se démarque très clairement de celle des autres caséidés par leur morphologie bulbeuse avec des couronnes très courtes et larges. La connaissance de la denture supérieure est limitée à quatre dents, deux étant portées par un prémaxillaire incomplet et deux autres provenant d’un fragment de maxillaire. La première dent prémaxillaire est très différente de celle des autres genres de caséidés, en étant épaisse à la base et se rétrécissant en une terminaison émoussée de la couronne. La deuxième dent est de forme similaire, mais beaucoup plus petite. Ni l'une ni l'autre ne montre de cuspides. Les deux dents maxillaires ont une structure générale similaire à celle des autres caséidés dérivés en ayant une base à section circulaire, suivi à mis hauteur de la dent d’un élargissement de la couronne, laquelle se termine ensuite par un amincissement labio-lingual donnant à l’extrémité supérieure de la couronne une morphologie spatulée. Elles diffèrent toutefois de celles des autres caséidés en étant plus courtes et plus émoussées, la couronne se terminant par un bord aplati portant cinq petites cuspides longitudinales. La paire de mâchoires inférieures conserve une denture beaucoup plus complète. Les trois dents antérieures sont très distinctives. Elles sont très courtes et émoussées avec une partie médiane très élargie. Elle ne montre pas de cuspides. Les dents plus postérieures sont courtes et ressemblent assez étroitement aux supérieures. Les cinq cuspides sur la marge dorsale aplatie des couronnes sont très distinctes. Il y a 11 dents sur le côté droit et 12 sur le gauche, en comptant comme présentes celles qui sont en cours de remplacement. Apparemment, 12 est le nombre correct, ce qui est le nombre de dents le plus bas parmi les caséidés d’Amérique du Nord. Seul le genre Russe Ennatosaurus en possède moins avec seulement 10 dents par demi mâchoires. Les dents antérieures des mandibules d’A. romeri sont relativement usées. Certaines des dents les plus postérieures sont usées et d'autres non. Les cuspides s’effacent rapidement avec l'usure et les étapes finales produisent une couronne en forme de bouton dont la surface est formée par la zone bulbeuse et arrondie de la partie centrale de la dent. La robustesse de ces dents, associée à l’usure importante qu’elles présentent, indiquent que le régime alimentaire d’A. romeri devait inclure des végétaux plus coriaces que ceux dont se nourrissait la plupart des autres caséidés herbivores[1].
Le squelette postcrânien d’A. romeri diffère principalement de celui des deux autres espèces du genre par des caractéristiques du bassin, du fémur, et de l’humérus. L’humérus, bien que large et lourd avec des arêtes et des processus fortement développés, diffère de celui d’A. dolani par ses proportions moins massives et la présence d’une diaphyse bien distincte quoique relativement courte[1]. Le bassin est caractérisé par un pubis et un ischion formant une large plaque un peu en forme de selle avec une symphyse médiane fortement ossifiée alors qu'elle était cartilagineuse chez A. dolani. Sur l’holotype, le sacrum ne compte que trois vertèbres avec trois côtes sacrales associées[5]. Toutefois, le paratype FMNH UR 904 possède lui quatre côtes sacrales indiquant la présence de quatre vertèbres dans le sacrum de ce spécimen. Il est probable que quatre soit le nombre normal de vertèbres sacrées chez cette espèce, le sacrum de l’holotype étant en fait incomplet[1]. Sur l’holotype d’A. romeri, les trois vertèbres sacrées sont soudées alors que chez A. dolani, seules les deux premières sont fusionnées[5]. Les côtes sacrales sont en forme de lame et sont presque complètement séparées les unes des autres au niveau de l’articulation avec l’ilium. Tout au plus est observé une légère fusion entre la deuxième et la troisième côte sacrale[5] - [1]. Le fémur est relativement petit et fortement ossifié. Il est caractérisé par la position du trochanter interne, lequel est situé à peu près à mi-chemin entre les limites proximale et distale de la fosse intertrochantérienne. Le quatrième trochanter est bien développé et s’élève légèrement au-dessus de la crête de l’adducteur. Le condyle distal antérieur est peu profond et plat et est bien séparé du condyle postérieur. Ce dernier est plus grand et porte une surface bien définie pour l'articulation avec la fibula. L'espace poplité est proéminent[5].
Répartition stratigraphique
L’âge de la formation de San Angelo a fait l’objet de nombreuses interprétations, celles-ci lui attribuant alternativement un âge Cisuralien terminal (Kungurien) et/ou Guadalupien basal (Roadien)[8]. La formation de San Angelo surmonte le Groupe de Clear Fork et est recouverte par la formation de Blaine. D’après Lucas et d’autres auteurs, des fusulines trouvées dans une intercalation marine de la formation de San Angelo, ainsi que des ammonoïdes présentes à la base de la formation de Blaine sus-jacente, indiquaient un âge Kungurien. De plus, selon ces auteurs, la base de la formation de San Andres, située plus à l’ouest et considérée comme un équivalent latéral de la formation de Blaine, se trouve dans la zone de conodontes de Neostreptognathodus prayi, la seconde des trois biozones de conodontes du Kungurien. La base de la formation de Blaine appartiendrait donc à cette biozone du Kungurien, ce qui suggère que la formation de San Angelo sous-jacente et Angelosaurus seraient légèrement plus anciens que la zone de conodonte à N. prayi avec un âge Kungurien inférieur[9] - [10] - [11]. Cependant, Michel Laurin et Robert W. Hook ont fait valoir que l’intercalation marine à fusulines citée plus haut n’appartient pas à la formation de San Angelo dans laquelle elle fut incluse par erreur, et ne peut pas être utilisée pour dater cette dernière. Le nom de formation de San Angelo a été incorrectement appliqué à une grande variété de roches dans divers bassins sédimentaires situés dans l’ouest du Texas, alors que la Formation de San Angelo est limitée au plateau oriental et est exclusivement continentale et dépourvue de fossiles marins[8]. Par ailleurs, la révision taxonomique des ammonoïdes de la base de la formation de Blaine indique un âge Roadien plutôt que Kungurien[N 2] - [8]. La formation de San Angelo a quant à elle livré une flore fossile dominée par les conifères Voltziens, un assemblage plutôt caractéristique du Guadalupien et du Lopingien. Ainsi, selon Laurin et Hook la formation de San Angelo pourrait dater du Kungurien terminal ou du Roadien basal, ou plus probablement pourrait chevaucher la limite Kungurien/Roadien[8].
Avec une position stratigraphique équivalente à la partie centrale de la formation de Flowerpot, la formation de Chickasha contient la faune de tétrapodes la plus récente du Permien d’Amérique du Nord (incluant A. romeri)[7] - [8]. Sur la base de sa composition faunique, un âge Guadalupien (= Permien moyen) lui fut plusieurs fois assigné. Cette attribution repose principalement sur la présence dans la faune de Chickasha du parareptile Macroleter qui n’était connu que dans l’assemblage de Mezen en Russie européenne, lequel contient une faune de thérapsides non mammaliens typique du Guadalupien, mais aussi le caséidé Ennatosaurus[12]. Un autre indice possible d’un âge Guadalupien est la présence dans la faune de Chickasha du plus grand représentant connu des Varanopidae, Watongia, dont les grandes dimensions seraient le signe d’une augmentation de taille tardive dans l’évolution de ce clade, lequel aurait pris la place de prédateur du sommet en l’absence des Sphenacodontidae[13]. Toutefois, la présence du genre Macroleter à la fois en Russie et en Oklahoma ne garantit pas que l’assemblage de Mezen et la formation de Chickasha soient contemporains car durant le Permien divers genres de tétrapodes avaient une large distribution temporelle, tels Dimetrodon et Diplocaulus[14]. Autre exemple, le varanopidé Mesenosaurus, initialement connu dans l’assemblage Guadalupien de Mezen, est également représenté par une espèce distincte dans un gisement de l’Oklahoma daté radiométriquement du milieu du Permien inférieur (Artinskien), soit un écart d’âge d’au moins 20 millions d’années avec l’espèce Russe[14]. Ainsi, l’âge de la formation de Chickasha peut difficilement être évaluée à partir de la faune qu’elle contient. Certains auteurs lui ont assigné un âge Kungurien[9] - [11]. Cependant, des données magnétostratigraphiques suggèrent que la formation de Chickasha date probablement du Roadien inférieur[8]. Cet âge est également cohérent avec la position stratigraphique de la formation de Chickasha qui occupe une position légèrement plus élevée que celle de la formation de San Angelo datée de la limite Kungurien-Roadien[8].
Paléoenvironnements
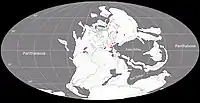
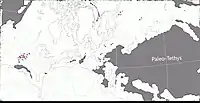
Au Permien, la plupart des terres émergées étaient réunies en un seul supercontinent, la Pangée. Celui-ci avait alors grossièrement la forme d’un C : ses parties nord (la Laurasia) et sud (le Gondwana) étaient reliés à l'ouest mais séparés à l'est par une très vaste baie océanique - la mer de Téthys[15]. Un long chapelet de microcontinents, regroupés sous le nom de Cimmérie, divisait la Téthys en deux : la Paleo-Tethys au nord, et la Neo-Tethys au sud[16]. Les formations de San Angelo et de Chickasha correspondent principalement à des sédiments fluviatiles (et éoliens) déposés dans une vaste plaine deltaïque parsemée de lacs et de lagunes. Cette plaine côtière était bordée à l’ouest par une mer qui occupait ce qui est maintenant le Golfe du Mexique et la partie la plus méridionale de l'Amérique du Nord. Les cours d’eau alimentant le delta se dirigeaient depuis des reliefs modestes situés plus à l’est et correspondant aux soulèvements ancestraux des montagnes Ouachita, Arbuckle et Wichita. Le climat était subtropical avec des pluies modérées et saisonnières. Il existait une mousson d’été ainsi qu’une saison sèche hivernale. La mousson était relativement faible, en raison de la taille limitée de la mer et du faible différentiel entre les températures d’été et d’hiver. La présence d’évaporites indique une aridité importante interrompue par des inondations saisonnières[3] - [2] - [17] - [18] - [11].
La formation de San Angelo est composée à sa base de grès dur, vert, gris et bruns, et de conglomérats fins, non fossilifères. La partie centrale de la formation est principalement constituée de mudstones rouges interprétés comme des boues argileuses et limoneuses déposées dans les plaines côtières lors d’épisodes d’inondations périodiques. Le mode de conservation du spécimen type d’Angelosaurus dolani, avec la partie postérieure du squelette articulée et la partie antérieure incomplète et disloquée, suggère que l’animal a été piégé dans des sédiments boueux puis dévoré par des carnivores qui ont dispersés les parties exposées. Le caséidé Caseoides est également présent dans cette partie de la formation. Ces mudstones rouges sont intercalés d’un mince niveau de grès vert, de mudstones sableux, et d’évaporites. Ceux-ci correspondraient à un empiètement mineur et de courte durée des estuaires, des lagunes, et des mers très peu profondes sur la partie terrestre du delta. La partie supérieure de la formation de San Angelo est caractérisée par la prépondérance des sédiments grossiers tels que grès et conglomérats, mais incluant aussi à sa base des mudstones sableux et à son sommet des mudstones rouges purs. D’après Olson, ces sédiments furent déposés par des rivières plus larges et aux courants plus puissants que celles de la partie centrale de la formation. Cependant, en Oklahoma, les strates équivalentes à la formation de San Angelo, qui étaient elles aussi considérées comme des dépôts fluviaux-deltaïques et fluviaux côtiers, ont été réinterprétées comme étant d’origine éolienne[19]. La partie supérieure non sommitale de la formation de San Angelo est caractérisée par l’absence du genre Angelosaurus et l’abondance du caséidé Cotylorhynchus hancocki, lequel est associé, entre autres, à Caseopsis agilis, au Sphenacodontidae Dimetrodon angelensis, et aux Captorhinidae Rothianiscus multidonta, et Kahneria seltina. Tout au sommet de la formation de San Angelo, les mudstones rouges sont à nouveau le faciès sédimentaire dominant. Le genre Angelosaurus est à nouveau présent (avec l’espèce A. greeni), accompagné par Caseoides cf. agilis, R. multidonta et K. seltina[3] - [2] - [1].
Dans l’Oklahoma, la formation de Chickasha correspond à la partie centrale de la formation de Flowerpot dans laquelle elle s’intercale localement. Les sédiments qui la composent sont variés et comportent des shales rouges, des grès, des mudstones, des conglomérats, et des évaporites, déposés dans les plaines inondables et les chenaux bordant la mer et les lagunes côtières. Dans la carrière d’Oméga, tous les fossiles proviennent de grès, de mudstones et de conglomérats durs et siliceux, disposés en lentilles. Ils correspondent à des dépôts d’un ancien chenal d’environ 1,50 m d’épaisseur et 4,6 m de largeur où se sont accumulés les squelettes de l’espèce Angelosaurus romeri, mais aussi ceux d’un second caséidé, Cotylorhynchus bransoni, et ceux du captorhinidé Rothianiscus robustus[5] - [7] - [8]. Ailleurs dans cette formation sont connus le Xenacanthiforme Orthacanthus, le Nectridea Diplocaulus[8], le temnospondyle Dissorophidae Nooxobeia[20], le pareiasauromorpha Nycteroleteridae Macroleter[12] - [21], et les varanopidés Varanodon et Watongia[7] - [13].
Phylogénie
Toutes les études phylogénétiques des caséidés considèrent Angelosaurus comme un taxon proche des genres Ennatosaurus et Cotylorhynchus. Dans la première analyse phylogénétique des caséidés publiée en 2008, l’espèce Angelosaurus dolani est récupérée comme le groupe frère de Cotylorhynchus romeri[22].
Ci-dessous le premier cladogramme des Caseidae publié par Maddin et al. en 2008[22].
| Caseasauria |
| ||||||||||||||||||||||||||||||||||||
Une autre analyse phylogénitique réalisée en 2012 par Benson n’inclut que l’espèce la mieux connue, A. romeri. Celle-ci est identifiée comme le groupe frère du genre Cotylorhynchus[23].
Ci-dessous, le cladogramme des Caseasauria publié par Benson en 2012[23].
En 2015, Romano et Nicosia ont publié la première étude cladistique incluant presque tous les Caseidae, à l’exception de taxons très fragmentaires comme Alierasaurus ronchii et Angelosaurus greeni. Dans cette analyse, Angelosaurus forme un clade avec le caséidé Russe Ennatosaurus, et ce clade est le groupe frère d’un clade contenant les genres Cotylorhynchus et Ruthenosaurus[24].
Ci-dessous, le cladogramme des Caseidae publié par Romano et Nicosia en 2015[24].
En 2020, deux cladogrammes publiés par Berman et des collègues montre que le genre Angelosaurus (représenté par A. romeri) occupe une position plus dérivée que Ennatosaurus. Dans le premier cladogramme, Angelosaurus forme avec Alierasaurus et les trois espèces de Cotylorhynchus, une polytomie non résolue. Dans le second cladogramme, Angelosaurus est le groupe frère d’un clade contenant Alierasaurus et Cotylorhynchus[25].
Ci-dessous, les deux cladogrammes des Caseidae publié par Berman et des collègues en 2020[25].
Une analyse phylogénétique publiée en 2022 par Werneburg et des collègues suggère que les genres Angelosaurus et Cotylorhynchus seraient paraphylétiques, les taxons autres que leur espèce type pouvant appartenir à des genres différents[4].
Ci-dessous, le cladogramme publié par Werneburg et des collègues en 2022[26].
Publication originale
- (en) Everett C. Olson et James R. Beerbower, « The San Angelo Formation, Permian of Texas, and Its Vertebrates », The Journal of Geology, Chicago, University of Chicago Press, vol. 61, no 5, , p. 389-423 (ISSN 0022-1376 et 1537-5269, OCLC 1608377, DOI 10.1086/626109)
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- Ces mudstones furent initialement attribués par Olson (1962) à la partie inférieure de la formation de Flowerpot. En 1974, Gary E. Smith a redéfini ces mudstones comme formant le sommet de la formation de San Angelo.
- Pris individuellement les sept genres d’ammonoïdes de cette faune ont une distribution temporelle s’étendant du Cisuralien au Lopingien : trois sont connus dans les gisements du Cisuralien et du Guadalupien, deux s'étendent du Cisuralien au Lopingien, et deux sont enregistrés dans les roches du Guadalupien au Lopingien inférieur. Cependant, le Roadien (= base du Guadalupien) est le seul étage durant lequel les sept genres coexistent.
Références taxonomiques
- (en) Référence Paleobiology Database : Angelosaurus Olson & Beerbower, 1953 † (consulté le )
- (en) Caseidae
Références
- (en) E.C. Olson, « The family Caseidae », Fieldianna (Geology), vol. 17, , p. 225–349
- (en) E.C. Olson, « Late Permian terrestrial vertebrates, U.S.A and U.S.S.R. », Transactions of the American Philosophical Society, New Series, vol. 52, , p. 1-224 (DOI 10.2307/1005904)
- (en) E.C. Olson et J.R. Beerbower, « The San Angelo formation, Permian of Texas and its vertebrates », The Journal of Geology, vol. 61, no 5, , p. 384-423
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2)
- (en) E.C. Olson et H. Barghusen, « Vertebrates from the Flowerpot formation, Permian of Oklahoma, Part I of Permian Vertebrates from Oklahoma and Texas », Oklahoma Geological Survey, circular 59, , p. 5-48 (lire en ligne)
- (en) A. Ronchi, E. Sacchi, M. Romano et U. Nicosia, « A huge caseid pelycosaur from north-western Sardinia and its bearing on European Permian stratigraphy and palaeobiogeography », Acta Palaeontologica Polonica, vol. 56, no 4, , p. 723-738 (DOI 10.4202/app.2010.0087)
- (en) E.C. Olson, « New Permian Vertebrates from the Chickasha formation in Oklahoma », Oklahoma Geological Survey, circular 70, , p. 1-70 (lire en ligne)
- (en) M. Laurin et R.W. Hook, « The age of North America’s youngest Paleozoic continental vertebrates : a review of data from the Middle Permian Pease River (Texas) and El Reno (Oklahoma) Groups », BSGF – Earth Sciences Bulletin, vol. 193, no 10, , p. 1-30 (DOI 10.1051/bsgf/2022007)
- (en) S.G. Lucas, « Global Permian tetrapod biostratigraphy and biochronology », dans Lucas, S.G., Cassinis, G. and Schneider, J.W., Non-Marine Permian Biostratigraphy and Biochronology, London, Geological Society, Special Publications, 265, (ISBN 978-1-86239-206-9), p. 65–93
- (en) S.G. Lucas et V.K. Golubev, « Age and duration of Olson’s Gap, a global hiatus in the Permian tetrapod fossil record », Permophiles, vol. 67, , p. 20- 23 (lire en ligne)
- (en) J.W. Schneider, S.G. Lucas, F. Scholze, S. Voigt, L. Marchetti, H. Klein, S. Opluštil, R. Werneburg, V.K. Golubev, J.E. Barrick, T. Nemyrovska, A. Ronchi, M.O. Day, V.V. Silantiev, R. Rößler, H. Saber, U. Linnemann, V. Zharinova et S-Z. Shen, « Late Paleozoic–early Mesozoic continental biostratigraphy — Links to the Standard Global Chronostratigraphic Scale », Palaeoworld, vol. 29, no 2, , p. 186-238 (DOI 10.1016/j.palwor.2019.09.001)
- (en) R.R. Reisz et M. Laurin, « The reptile Macroleter: First vertebrate evidence for correlation of Upper Permian continental strata of North America and Russia », Geological Society of America Bulletin, vol. 113, no 9, , p. 1229-1233 (DOI 10.1130/0016-7606(2001)113<1229:TRMFVE>2.0.CO;2)
- (en) R.R. Reisz et M. Laurin, « A reevaluation of the enigmatic Permian synapsid Watongia and its stratigraphic significance », Canadian Journal of Earth Science, vol. 41, no 4, , p. 377-386 (DOI 10.1139/e04-016)
- (en) S. Maho, B.M. Gee et R.R. Reisz, « A new varanopid synapsid from the early Permian of oklahoma and the evolutionary stasis in this clade », Royal Society Open Science, vol. 6, no 10, , p. 1-16 (DOI 10.1098/rsos.191297)
- (en) S. McLoughlin, « The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism », Australian Journal of Botany, vol. 49, , p. 271-300 (DOI 10.1071/BT00023)
- (en) A.M.C. Şengör, « Tectonics of the Tethysides: orogenic collage development in a collisional setting », Annual Review of Earth and Planetary Sciences, no 15, , p. 214-244 (DOI 10.1146/annurev.ea.15.050187.001241)
- (en) G.E. Smith, « Depositional Systems, San Angelo Formation (Permian), North Central Texas – Facies Control of Red-Bed Copper Mineralization », The University of Texas at Austin Bureau of Economic Geology Report of Investigation, vol. 80, , p. 1-74 (DOI 10.23867/RI0080D)
- (en) T.S. Kemp, « Pelycosaurs », dans Kemp, T.S., Mammal-like reptiles and the origin of Mammals, London, Academic Press, (ISBN 978-0124041202), p. 73
- (en) T.M. Foster, G.S. Soreghan, M.J. Soreghan, K.C. Benison et R.D. Elmore, « Climatic and paleogeographic significance of eolian sediment in the Middle Permian Dog Creek Shale (Midcontinent U.S.) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 402, , p. 12-29 (DOI 10.1016/j.palaeo.2014.02.031)
- (en) B.M. Gee, D.D. Scott et R.R. Reisz, « Reappraisal of the Permian dissorophid Fayella chickashaensis », Canadian Journal of Earth Sciences, vol. 55, no 10, , p. 1103-1114 (DOI 10.1139/cjes-2018-0053)
- (en) R.R. Reisz et M. Laurin, « Discussion and reply : The reptile Macroleter: First vertebrate evidence for correlation of Upper Permian continental strata of North America and Russia – Reply », Geological Society of America Bulletin, vol. 114, no 9, , p. 1176-1177
- (en) H.C. Maddin, C.A. Sidor et R.R. Reisz, « Cranial anatomy of Ennatosaurus tecton (Synapsida: Caseidae) from the Middle Permian of Russia and the evolutionary relationships of Caseidae », Journal of Vertebrate Paleontology, vol. 28, no 1, , p. 160-180 (DOI 10.1671/0272-4634(2008)28[160:CAOETS]2.0.CO;2)
- (en) R.B.J. Benson, « Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies », Journal of Systematic Palaeontology, vol. 10, no 4, , p. 601-624 (DOI 10.1080/14772019.2011.631042)
- (en) M. Romano et U. Nicosia, « Cladistic analysis of Caseidae (Caseasauria, Synapsida): using the gap-weighting method to include taxa based on incomplete specimens », Palaeontology, vol. 58, no 6, , p. 1109–1130 (DOI 10.1111/pala.12197)
- (en) D.S. Berman, H.C. Maddin, A.C. Henrici, S.S. Sumida, D. Scott et R.R. Reisz, « New primitive Caseid (Synapsida, Caseasauria) from the Early Permian of Germany », Annals of Carnegie Museum, vol. 86, no 1, , p. 43-75 (DOI 10.2992/007.086.0103)
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2).