Caseoides
Caseoides sanangeloensis

Caseoides est un genre éteint et fossile de synapsides Caseidae ayant vécu à la fin du Permien inférieur (Kungurien) ou au début du Permien moyen (Roadien) dans ce qui est aujourd’hui la partie sud-centrale des États-Unis. Le genre n’est représenté que par son espèce type, Caseoides sanangeloensis, qui fut nommée en 1953 par Everett C. Olson et James Richard Beerbower (d), à partir de deux squelettes fragmentaires découverts dans la partie centrale de la formation de San Angelo, dans le comté de Knox au Texas.
Étymologie
Le nom de genre signifie semblable à (au genre) Casea et l’épithète spécifique sanangeloensis réfère à la formation de San Angelo qui a livré ce taxon.
Description
Caseoides est un petit caséidé représenté par des restes très fragmentaires. L’holotype CNHM UR 151 est représenté par un membre postérieur droit et une partie du pied, un fémur gauche incomplet, un humérus, et des fragments de vertèbres incluant deux centra découvert sur le site de Little Crotton Creek, dans le Comté de Knox au Texas. Un autre spécimen fragmentaire trouvé à moins de 8 mètres de distance de l’holotype et 3 mètres plus bas stratigraphiquement se compose des fémurs droit et gauche, la partie sommitale de l’ilion, une partie de la région symphysaire du pubis, un centre vertébral, et d’autres débris indéterminables[1] - [2] - [3]. Selon Olson et Beerbower, Caseoides se distinguerait par les os de ses membres proportionnellement larges par rapport à leur longueur comparé à ceux de Casea broilii. La tête du tibia est aussi proportionnellement grande, ressemblant à celle de Cotylorhynchus romeri. Les extrémités de l’humérus et leurs processus sont mal ossifiés. Les éléments tarsiens sont également incomplètement ossifiés. Les centres vertébraux sont profondément amphicoeles[1]. Plus récemment, Marco Romano a également précisé que Caseoides possède un humérus dont la diaphyse est proportionnellement plus robuste que celle de Cotylorhynchus[4]. Caseoides reste un animal mal connu et pas très bien défini scientifiquement. Selon Romano, Caseoides est peut-être bien un taxon valide en étant un caséidé de petite taille aux proportions robustes, mais cet auteur estime également que les restes de cet animal pourraient correspondre à des spécimens juvéniles de caséidés de plus grandes tailles présents dans la formation de San Angelo (Angelosaurus et Cotylorhynchus). Une hypothèse difficile à tester en raison notamment du manque de séries ontogénétiques suffisamment complètes chez les grands caséidés[4].
Répartition stratigraphique
L’âge de la formation de San Angelo a fait l’objet de nombreuses interprétations, celles-ci lui attribuant alternativement un âge Cisuralien terminal (Kungurien) et/ou Guadalupien basal (Roadien) [5]. La formation de San Angelo surmonte le Groupe de Clear Fork et est recouverte par la formation de Blaine. D’après Spencer G. Lucas et d’autres auteurs, des fusulines trouvées dans une intercalation marine de la formation de San Angelo, ainsi que des ammonoïdes présentes à la base de la formation de Blaine sus-jacente, indiquaient un âge Kungurien. De plus, selon ces auteurs, la base de la formation de San Andres, située plus à l’ouest et considérée comme un équivalent latéral de la formation de Blaine, se trouve dans la zone de conodontes de Neostreptognathodus prayi, la seconde des trois biozones de conodontes du Kungurien. La base de la formation de Blaine appartiendrait donc à cette biozone du Kungurien, ce qui suggère que la formation de San Angelo sous-jacente et Caseopsis seraient légèrement plus anciens que la zone de conodonte à N. prayi avec un âge Kungurien inférieur[6] - [7] - [8]. Cependant, Michel Laurin et Robert W. Hook ont fait valoir que l’intercalation marine à fusulines citée plus haut n’appartient pas à la formation de San Angelo dans laquelle elle fut incluse par erreur, et ne peut pas être utilisée pour dater cette dernière. Le nom de formation de San Angelo a été incorrectement appliqué à une grande variété de roches dans divers bassins sédimentaires situés dans l’ouest du Texas, alors que la Formation de San Angelo est limitée au plateau oriental et est exclusivement continentale et dépourvue de fossiles marins[5]. Par ailleurs, la révision taxonomique des ammonoïdes de la base de la formation de Blaine indique un âge Roadien plutôt que Kungurien[N 1] - [5]. La formation de San Angelo a quant à elle livré une flore fossile dominée par les les conifères voltziens, un assemblage plutôt caractéristique du Guadalupien et du Lopingien. Ainsi, selon Laurin et Hook la formation de San Angelo pourrait dater du Kungurien terminal ou du Roadien basal, ou plus probablement pourrait chevaucher la limite Kungurien/Roadien[5].
Paléoenvironnements
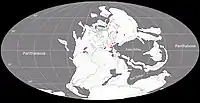
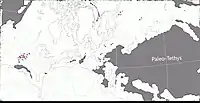
Au Permien, la plupart des terres émergées étaient réunies en un seul supercontinent, la Pangée. Celui-ci avait alors grossièrement la forme d’un C : ses parties nord (la Laurasia) et sud (le Gondwana) étaient reliés à l'ouest mais séparés à l'est par une très vaste baie océanique - la mer de Téthys[9]. Un long chapelet de microcontinents, regroupés sous le nom de Cimmérie, divisait la Téthys en deux : la Paleo-Tethys au nord, et la Neo-Tethys au sud[10]. La formation de San Angelo correspond à des sédiments fluviatiles et/ou éoliens déposés dans une vaste plaine deltaïque parsemée de lacs et de lagunes. Cette plaine côtière était bordée à l’ouest par une mer qui occupait ce qui est maintenant le Golfe du Mexique et la partie la plus méridionale de l'Amérique du Nord. Les cours d’eau alimentant le delta se dirigeaient depuis des reliefs modestes situés plus à l’est et correspondant aux soulèvements ancestraux des montagnes Ouachita, Arbuckle et Wichita. Le climat était subtropical avec des pluies modérées et saisonnières. Il existait une mousson d’été ainsi qu’une saison sèche hivernale. La mousson était relativement faible en raison de la taille limitée de la mer et du faible différentiel entre les températures d’été et d’hiver. La présence d’évaporites indique une aridité importante interrompue par des inondations saisonnières[1] - [2] - [11] - [12] - [13] - [8].
La formation de San Angelo est composée à sa base de grès dur, vert, gris et bruns, et de conglomérats fins, non fossilifères. La partie centrale de la formation est principalement constituée de mudstones rouges interprétés comme des boues argileuses et limoneuses déposées dans les plaines côtières lors d’épisodes d’inondations périodiques. Ces mudstones rouges sont intercalés d’un mince niveau de grès vert, de mudstones sableux, et d’évaporites. Ceux-ci correspondraient à un empiètement mineur et de courte durée des estuaires, des lagunes, et des mers très peu profondes sur la partie terrestre du delta. Caseoides et le caséidé Angelosaurus dolani proviennent des mudstones rouges de cette partie de la formation[N 2]. La partie supérieure de la formation de San Angelo est caractérisée par la prépondérance des sédiments grossiers tels que grès et conglomérats, mais incluant aussi à sa base des mudstones sableux et à son sommet des mudstones rouges purs. D’après Olson, ces sédiments furent déposés par des rivières plus larges et aux courants plus puissants que celles de la partie centrale de la formation[2]. Cependant, en Oklahoma, les strates équivalentes à la formation de San Angelo, qui étaient elles aussi considérées comme des dépôts fluviaux-deltaïques et fluviaux côtiers, ont été réinterprétées comme étant d’origine éolienne[14]. La faune de la partie supérieure de la formation de San Angelo est dominée (par le nombre de spécimens) par le caséidé Cotylorhynchus hankocki, et comprend, entre autres[N 3], le caséidé Caseopsis agilis, le sphénacodontidé Dimetrodon angelensis, les Captorhinidae Rothianiscus multidonta, et Kahneria seltina, et le dvinosaure Tupilakosauridae Slaugenhopia[1] - [2]. Quelques pistes de tétrapodes indiquent également la présence de paréiasauromorpha nycteroleteridés (ichnotaxon Pachypes ollieri)[15], dont un squelette partiel est connu dans les dépôts légèrement plus récents de la formation de Chickasha en Oklahoma[16] - [15]. Une flore inhabituelle a été retrouvée dans les chenaux de la partie supérieure de la formation de San Angelo. Elle est dominée par les gymnospermes et est remarquable par sa composition unique incluant à la fois des taxons typiques du Permien inférieur tels Walchia ou Culmitzschia mais aussi des formes qui n’étaient jusque-là connus que dans le Permien moyen ou supérieur comme diverses espèces d’Ulmannia, Pseudovoltzia liebeana, et le taxon d’affinité incertaine Taeniopteris eckardtii, ou dans les strates du Mésozoïque comme la bennettitale Podozamites et le cycadidé putatif Dioonitocarpidium. Le reste de la flore est représentée par le ginkgoale Dicranophyllum, la cordaitale Cordaites, et l’équisetale cf. Neocalamites[17] - [18].
Phylogénie
En 1968 Olson suggérait que Caseoides était le dernier représentant d’une ligné évolutive incluant également “Casea” nicholsi et “Casea” halselli (deux espèces aujourd’hui considérées comme n’appartenant pas au genre Casea). Selon cet auteur, cette ligné montrerait au cours de son évolution une augmentation de la taille et de la robustesse des os des membres[3]. En 2015, Romano et Nicosia ont publié la première étude cladistique prenant en compte la plupart des caseidés fragmentaires à l’exception d’Alierasaurus ronchii et Angelosaurus greeni. Dans cette analyse, Caseoides est le groupe frère d’un clade incluant “Casea” nicholsi et Caseopsis agilis[19].
Ci-dessous, le cladogramme des Caseidae publié par Romano et Nicosia en 2015[19].
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- Pris individuellement les sept genres d’ammonoïdes de cette faune ont une distribution temporelle s’étendant du Cisuralien au Lopingien : trois sont connus dans les gisements du Cisuralien et du Guadalupien, deux s'étendent du Cisuralien au Lopingien, et deux sont enregistrés dans les roches du Guadalupien au Lopingien inférieur. Cependant, le Roadien (= base du Guadalupien) est le seul étage durant lequel les sept genres coexistent.
- De ces niveaux proviennent également les genres très fragmentaires Steppesaurus et Mastersonia. En 1953, Olson et Beerbower classèrent d’abord Steppesaurus parmi les pélycosaures Sphenacodontidae. Puis en 1962 Olson le considéra comme un thérapside Phtinosuchidae. Dans le même article Olson décrivit Mastersonia qu’il considérait comme un thérapside « éodinocéphalien ». Dans une courte note publiée en 1995, Sidor et Hopson ont rejeté la nature thérapside de ces deux taxons, leurs restes appartenant selon eux à des pélycosaures. Laurin et Hook (2022) souligne toutefois la nécessité de réétudier ces spécimens avec les méthodes modernes.
- Olson et Beerbower (1953), puis Olson (1962), ont nommés d’autres taxons fragmentaires de la partie supérieure de la formation de San Angelo qu’ils attribuèrent à divers groupes de thérapsides, comme Knoxosaurus, Gorgodon, Dimacrodon, Eosyodon, Driveria, et Tappenosaurus. En 1995 Sidor et Hopson ont réinterprété les fossiles de tous ces taxons comme des restes de pélycosaures caséidés et sphénacodontidés plus ou moins déterminables. Cependant, une nouvelle étude de ces spécimens par des méthodes modernes serait nécessaire (Laurin et Hook, 2022).
Références
- (en) E.C. Olson et J.R. Beerbower, « The San Angelo formation, Permian of Texas and its vertebrates », The Journal of Geology, vol. 61, no 5, , p. 384-423
- (en) E.C. Olson, « Late Permian terrestrial vertebrates, U.S.A and U.S.S.R. », Transactions of the American Philosophical Society, New Series, no 52, , p. 1-224 (DOI 10.2307/1005904)
- (en) E.C. Olson, « The family Caseidae », Fieldianna (Geology), vol. 17, , p. 225–349
- (en) M. Romano, « Long bone scaling of caseid synapsids: a combined morphometric and cladistic approach », Lethaia, vol. 50, no 4, , p. 511-526 (DOI 10.1111/let.12207)
- (en) M. Laurin et R.W. Hook, « The age of North America’s youngest Paleozoic continental vertebrates : a review of data from the Middle Permian Pease River (Texas) and El Reno (Oklahoma) Groups », BSGF – Earth Sciences Bulletin, vol. 193, no 10, , p. 1-30 (DOI 10.1051/bsgf/2022007)
- (en) S.G. Lucas, « Global Permian tetrapod biostratigraphy and biochronology », dans Lucas, S.G., Cassinis, G. and Schneider, J.W., Non-Marine Permian Biostratigraphy and Biochronology, London, Geological Society, Special Publications, 265, (ISBN 978-1-86239-206-9), p. 65–93
- (en) S.G. Lucas et V.K. Golubev, « Age and duration of Olson’s Gap, a global hiatus in the Permian tetrapod fossil record », Permophiles, vol. 67, , p. 20- 23 (lire en ligne)
- (en) J.W. Schneider, S.G. Lucas, F. Scholze, S. Voigt, L. Marchetti, H. Klein, S. Opluštil, R. Werneburg, V.K. Golubev, J.E. Barrick, T. Nemyrovska, A. Ronchi, M.O. Day, V.V. Silantiev, R. Rößler, H. Saber, U. Linnemann, V. Zharinova et S-Z. Shen, « Late Paleozoic–early Mesozoic continental biostratigraphy — Links to the Standard Global Chronostratigraphic Scale », Palaeoworld, vol. 29, no 2, , p. 186-238 (DOI 10.1016/j.palwor.2019.09.001)
- (en) S. McLoughlin, « The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism », Australian Journal of Botany, vol. 49, , p. 271-300 (DOI 10.1071/BT00023)
- (en) A.M.C. Şengör, « Tectonics of the Tethysides: orogenic collage development in a collisional setting », Annual Review of Earth and Planetary Sciences, vol. 15, , p. 214-244 (DOI 10.1146/annurev.ea.15.050187.001241)
- (en) G.E. Smith, « Depositional Systems, San Angelo Formation (Permian), North Central Texas – Facies Control of Red-Bed Copper Mineralization », The University of Texas at Austin Bureau of Economic Geology Report of Investigation, vol. 80, , p. 1-74 (DOI 10.23867/RI0080D)
- (en) T.S. Kemp, « Pelycosaurs », dans Kemp, T.S., Mammal-like reptiles and the origin of Mammals, London, Academic Press, (ISBN 978-0124041202), p. 73
- (en) M.J. Soreghan, M.M. Swift et G.S. Soreghan, « Provenance of Permian eolian and related strata in the North American midcontinent: Tectonic and climatic controls on sediment dispersal in western tropical Pangea », dans Ingersoll, R.V., Lawton, T.F., and Graham, S.A., Tectonics, Sedimentary Basins, and Provenance: A Celebration of William R. Dickinson’s Career, Boulder, Geological Society of America Special Paper 540, (ISBN 978-0813725406, DOI 10.1130/2018.2540(28)), p. 661-688
- (en) T.M. Foster, G.S. Soreghan, M.J. Soreghan, K.C. Benison et R.D. Elmore, « Climatic and paleogeographic significance of eolian sediment in the Middle Permian Dog Creek Shale (Midcontinent U.S.) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 402, , p. 12-29 (DOI 10.1016/j.palaeo.2014.02.031)
- (en) L. Marchetti, S. Voigt, E. Mujal, S.G. Lucas, H. Francischini, J. Fortuny et V.L. Santucci, « Extending the footprint record of Pareiasauromorpha to the Cisuralian : earlier appearance and wider palaeobiogeography of the group », Papers in Palaeontology, vol. 7, no 3, , p. 1297-1319 (DOI 10.1002/spp2.1342)
- (en) R.R. Reisz et M. Laurin, « The reptile Macroleter: First vertebrate evidence for correlation of Upper Permian continental strata of North America and Russia », Geological Society of America Bulletin, vol. 113, no 9, , p. 1229-1233 (DOI 10.1130/0016-7606(2001)113<1229:TRMFVE>2.0.CO;2)
- (en) W.A. DiMichele, S.H. Mamay, D.S. Chaney, R.W. Hook et W.J. Nelson, « An Early Permian flora with Late Permian and Mesozoic affinities from north-central Texas », Journal of Paleontology, vol. 75, no 2, , p. 449-460 (DOI 10.1666/0022-3360(2001)075˂0449:AEPFWL˃2.0.CO;2)
- (en) W.A. DiMichele, N.J. Tabor, D.S. Chaney et W.J. Nelson, « From wetlands to wet spots: Environmental tracking and the fate of Carboniferous elements in Early Permian tropical floras », dans Greb, S.F. & DiMichele, W.A., Wetlands through time, Boulder, The Geological Society of America Special Paper 399, (DOI 10.1130/2006.2399(11)), p. 223-248
- (en) M. Romano et U. Nicosia, « Cladistic analysis of Caseidae (Caseasauria, Synapsida): using the gap-weighting method to include taxa based on incomplete specimens », Palaeontology, vol. 58, no 6, , p. 1109–1130 (DOI 10.1111/pala.12197)