Alierasaurus
Alierasaurus ronchii

Alierasaurus est un genre de synapsides Caseidae de très grande taille ayant vécu au début du Permien moyen (Roadien)[1] dans ce qui est aujourd’hui le nord-ouest de la Sardaigne[2].
Le genre n’est représenté que par son espèce type, Alierasaurus ronchii, fondée sur un squelette postcrânien partiel (MPUR NS 151) découvert dans la formation de Cala del Vino. Les dimensions des éléments préservés du pied et des vertèbres caudales suggèrent qu'Alierasaurus devait mesurer entre 6 et 7 m de longueur[3] - [4] - [5]. Il était donc d'une taille comparable, voire supérieure, à celle de l'espèce géante nord-américaine Cotylorhynchus hancocki de la formation de San Angelo au Texas, dont la taille avoisinait les six mètres de longueur. Alierasaurus diffère du genre Cotylorhynchus principalement par l’anatomie des os du pied en ayant un quatrième métatarsien plus long et plus mince, ainsi qu’une phalange unguéale pointue et en forme de griffe au lieu d’être aplatie. Alierasaurus et Cotylorhynchus avaient tous les deux une très grande cage thoracique en forme de tonneau, indiquant qu’il s'agissait d'herbivores se nourrissant principalement de végétaux riches en fibres[3].
Étymologie
Le nom de genre fait référence à Aliera, le nom de la ville d’Alghero en dialecte local, et de saurus signifiant lézard. L’espèce honore Ausonio Ronchi, le découvreur du spécimen[3].
Description

Les paléontologues Marco Romano (d) et Umberto Nicosia (d) ont identifié plusieurs autapomorphies dans l’anatomie du pied d’Alierasaurus : métatarsien IV avec une zone axiale distincte, environ deux fois plus longue que celle de la phalange proximale correspondante (au lieu d’un métatarsien court et massif comme chez les autres grands Caseidae) ; tête proximale du métatarsien IV non orthogonale à l'axe de l'os, formant un angle de 120° par rapport à la diaphyse : avec cette conformation, les têtes proximale et distale sont beaucoup plus proches le long du côté médial du métatarse ; phalanges unguéales en forme de griffe proportionnellement plus courtes que chez Cotylorhynchus , avec un double tubercule fléchisseur ventral très proche du bord proximal de la phalange ; axe des phalanges unguéales courbé vers le bas et médialement ; section transverse subtriangulaire et non spatulée comme chez Cotylorhynchus[3].
Découverte


L’holotype d’Alierasaurus fut découvert dans les niveaux supérieurs de la formation de Cala del Vino, au sommet du promontoire de la Tour de Porticciolo, lequel sépare le golfe de Porticciolo de la côte nord (près de la ville d’Alghero, Nurra, nord-ouest de la Sardaigne)[2]. Quelques os furent découverts à la surface du sol, dégagés par l’érosion, et d’autres encore enchâssés dans la roche constituée de mudstone et de siltstone. Ces sédiments furent déposés dans une ancienne plaine alluviale sous un climat semi-aride[2]. Le matériel connu est constitué de huit vertèbres caudales articulées, deux vertèbres caudales isolées, quatre vertèbres caudales distales, de nombreux gros fragments appartenant à au moins huit autres vertèbres, des portions proximales de sept chevrons, trois parties proximales de côtes dorsales, dix côtes fragmentaires indéterminées, une scapula droite mal conservée, un coracoïde droit écrasé, une extrémité distale de l’ulna gauche, et plusieurs os du pied représentés par un calcanéum fragmentaire, trois métapodes, cinq phalanges non-unguéales, une phalange unguéale presque complète, et deux phalanges unguéales partielles car dépourvues de leur extrémité distale[3]. Ces restes furent d’abord regardés comme appartenant possiblement à Cotylorhynchus (cf. Cotylorhynchus dans l’article de Ausonio Ronchi et al.) ou à un autre taxon étroitement apparenté. Plus tard, en dépit de l’absence des éléments du squelette les plus diagnostiques (notamment le crâne) pour la comparaison avec les autres Caseidae en général et avec Cotylorhynchus en particulier, le spécimen Sarde fut attribué à un nouveau genre nommé Alierasaurus, sur la base de quelques différences dans l’anatomie des os du pied[2] - [3]. En 2017, Marco Romano et des collègues décrivirent d’autres os appartenant au même individu (quelques vertèbres caudales et des fragments de chevrons et de côtes)[4]. Plus récemment, les mêmes niveaux ont livré des restes encore non décrits d'un pélycosaure sphénacodontidé et des empreintes de pas d'un troisième animal, lequel n'était connu que dans le sud de la France dans des terrains légèrement plus récents (ichnogenre Merifontichnus du Wordien de la formation de La Lieude dans le bassin de Lodève)[5] - [1].
Taphonomie

Les fossiles d’Alierasaurus furent découverts regroupés sur une surface de quelques mètres carré. Certains reposaient à même le sol, dégagés par l’érosion alors que d’autres se trouvaient encore dans le sédiment. La plupart des os étaient isolés à l’exception de deux os du pied et de 8 vertèbres caudales retrouvés articulés. Les os encore en place dans la roche ne se trouvaient pas tous sur le même plan de litage mais étaient enfouies à des profondeurs différentes au sein d’une couche de siltite rouge de 40 cm d’épaisseur. Plusieurs ossements furent fracturés avant l’enfouissement. La taphonomie du site indique un processus d’enfouissement complexe en plusieurs phases. Peu de temps après la mort de l’animal, la carcasse fut transportée du lieu du décès vers un second lieu de dépôt. Ce transport de courte durée fut suffisamment violent pour briser certains os. Par la suite, le corps, toujours à la surface des sédiments, a subi une nouvelle phase de décomposition assez courte avant qu’une nouvelle crue éclair n’emballe tous les restes dans une grande quantité de sédiments à grains fins, les transporte, et finalement les dépose tous ensemble dans un troisième endroit proche du précédent. Cette troisième phase de dépôt explique pourquoi les os se trouvent à différentes profondeurs dans la couche sédimentaire[2].
Paléogéographie
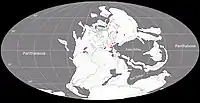
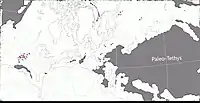
Au Guadalupien, la plupart des terres émergées étaient réunies en un seul supercontinent, la Pangée. Celui-ci avait alors grossièrement la forme d’un C : ses parties nord (la Laurasia) et sud (le Gondwana) étaient reliés à l'ouest mais séparés à l'est par une très vaste baie océanique - la mer de Téthys[6]. Un long chapelet de microcontinents, regroupés sous le nom de Cimmérie, divisait la Téthys en deux : la Paleo-Tethys au nord, et la Neo-Tethys au sud[7]. La Sardaigne était située dans la ceinture équatoriale de l’époque, au niveau du 10e parallèle nord. Elle ne constituait pas du tout une île et faisait partie intégrante de la Pangée. A cette époque, la Sardaigne (et la Corse) était connectée à ce qui est aujourd’hui le sud-est de la France[8] - [9]. La paléoposition précise du bloc Sardaigne-Corse a été déterminée au début des années 2000 à partir de corrélations lithostratigraphiques détaillées entre les successions du Permien et du Trias de Nurra dans le nord-ouest de la Sardaigne et du bassin de Toulon-Cuers dans le Var où la formation de Cala del Vino a comme équivalent la formation de Saint-Mandrier. Les similitudes lithologiques remarquables de la région de Nurra avec le bassin de Toulon-Cuers indiquent que les deux régions étaient positionnées face à face et constituaient deux parties d’un seul et même bassin sédimentaire[9] - [10] - [N 1]. Le bloc Sardaigne-Corse était tourné de 60° dans le sens des aiguilles d’une montre par rapport à son orientation actuelle. Le sud de la Sardaigne était alors situé près de l’est des Pyrénées (dont les montagnes n’existaient pas encore), et le nord-ouest de la Corse était positionnée devant l’Esterel (les rhyolites de la presqu’île de Scandola s’alignant avec celles de l’Esterel, d’âge et de composition similaire)[9] - [11].
Age
Aucune datation radiométrique n’est disponible pour la formation de Cala del Vino. Les estimations de son âge vont du Kungurien supérieur au Capitanien inférieur[2] - [1] - [12] - [13]. Ces âges sont inférés sur la base de corrélations stratigraphiques directes et indirectes avec les bassins permiens de Provence (dont le bassin de Toulon-Cuers qui constituait un seul bassin sédimentaire avec la région de Nurra) couplé à des données paléontologiques de Provence et d’Occitanie. La formation de Cala del Vino est corrélée lithostratigraphiquement avec la formation de Saint‐Mandrier du bassin de Toulon-Cuers. La formation de Saint-Mandrier n’a pas encore livré de fossiles, mais elle date probablement du Guadalupien car elle surmonte localement des calcaires lacustres et des mudstones noirs du Membre de Bau Rouge de la formation des Salettes qui ont livré respectivement une macroflore et une microflore suggérant un âge Kungurien supérieur - Roadien inférieur. La partie inférieure de la formation de Saint‐Mandrier est quant à elle corrélée avec la formation des Pradineaux du bassin de l’Esterel. Cette formation surmonte, au-dessus d’une lacune d’érosion, une formation rhyolitique (la Rhyolite A7) datée à 272,5 ± 0,3 Ma[10] - [14]. Cette âge absolue, autrefois considéré comme Kungurien supérieur[10] - [14], correspond aujourd’hui au Roadien inférieur[12]. D’autre part, la formation des Pradineaux contient dans sa partie inférieure une couche de rhyolite (la Rhyolite A11) qui elle-même est non datée mais qui est traversée par un filon de fluorite-baryte à adulaire datée à 264 ± 2 Ma correspondant à l’étage Capitanien, ce qui indique un âge plus ancien pour la Rhyolite A11 et la formation des Pradineaux[15]. Marc Durand suggère un âge Wordien, la lacune d’érosion au sommet de la Rhyolite A7 correspondant selon lui à une grande partie du Roadien[10] - [15]. La formation des Pradineaux a cependant livrée des plantes et des pollens fossiles suggérant un âge Roadien, une faune d’ostracodes indiquant un âge Roadien supérieur, et des pistes de vertébrés incluant l’ichnogenre Brontopus caractéristique du Guadalupien[10] - [16] - [17] - [12] - [18]. Par le biais des corrélations stratigraphiques, la formation Sarde de Cala del Vino pourrait ainsi être datée du Roadien - Wordien. Selon Werneburg et des collègues l’âge de la formation de Cala del Vino pourrait aussi s’étendre du Roadien au Capitanien inférieur comme la formation de La Lieude en Occitanie, en raison de ressemblances sédimentologiques et de la cooccurrence dans les deux formations des synapsides Caseidae et de l’ichnogenre Merifontichnus[13].
Phylogénie
En 2017 Marco Romano et des collègues ont publié la première analyse phylogénétique incluant le genre Alierasaurus. Celui-ci est récupéré comme le taxon frère du genre Cotylorhynchus[4].
En décrivant le genre Martensius en 2020, Berman et des collègues ont publié deux cladogrammes. Dans le premier, la position des caséidés plus dérivés que Martensius est mal résolue. Alierasaurus forme une polytomie avec Angelosaurus romeri et les trois espèces de Cotylorhynchus. Dans le second cladogramme, Alierasaurus est positionné au-dessus du genre Angelosaurus est forme une polytomie avec Cotylorhynchus romeri et un clade contenant les espèces C. bransoni et C. hancocki[19].
Ci-dessous, les deux cladogrammes des Caseidae publié par Berman et des collègues en 2020[19].
En 2022 Werneburg et des collègues décrivent le genre Lalieudorhynchus et publient une analyse phylogénétique qui conclut à la paraphylie des genres Angelosaurus et Cotylorhynchus, lesquels ne seraient représentés que par leur espèce type. Dans cette analyse, Cotylorhynchus romeri est positionnée juste au-dessus du genre Angelosaurus, et forme une polytomie avec un clade contenant Ruthenosaurus et Caseopsis et un autre clade contenant Alierasaurus, les deux autres espèces de Cotylorhynchus et Lalieudorhynchus. Au sein de ce dernier clade, Alierasaurus est le groupe frère de “Cotylorhynchus” bransoni et d’un clade plus dérivé incluant Lalieudorhynchus et “Cotylorhynchus” hancocki[13].
Ci-dessous, le cladogramme publié par Werneburg et des collègues en 2022[20].
Publication originale
- (en) Marco Romano et Umberto Nicosia, « Alierasaurus ronchii, gen. et sp. nov., a caseid from the Permian of Sardinia, Italy », Journal of Vertebrate Paleontology, SVP et Taylor & Francis, vol. 34, no 4, , p. 900-913 (ISSN 0272-4634 et 1937-2809, OCLC 238100068, DOI 10.1080/02724634.2014.837056)
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- Le bloc Corso-Sarde ne se détachera de ce qui deviendra le sud-est de la France qu'à partir de l'Oligocène (Durand, 2008).
Références
- (en) P. Citton, A. Ronchi, S. Maganuco, M. Caratelli, U. Nicosia, E. Sacchi et M. Romano, « First tetrapod footprints from the Permian of Sardinia and their palaeontological and stratigraphical significance », Geological Journal, , p. 1–15 (DOI 10.1002/gj.3285)
- (en) A. Ronchi, E. Sacchi, M. Romano et U. Nicosia, « A huge caseid pelycosaur from north-western Sardinia and its bearing on European Permian stratigraphy and palaeobiogeography », Acta Palaeontologica Polonica, vol. 56, no 4, , p. 723–738 (DOI 10.4202/app.2010.0087)
- Romano et Nicosia 2014, p. 900–913
- (en) M. Romano, A Ronchi, S. Maganuco et Nicosia, « New material of Alierasaurus ronchii (Synapsida, Caseidae) from the Permian of Sardinia (Italy), an dits phylogenetic affinities », Palaeontologia Electronica, vol. 20.2.26A, , p. 1–27 (DOI 10.26879/684)
- (en) M. Romano, P. Citton, S. Maganuco, E. Sacchi, M. Caratelli, A. Ronchi et U. Nicosia, « New basal synapsid discovery at the Permian outcrop of Torre del Porticciolo (Alghero, Italy) », Geological Journal, , p. 1–13 (DOI 10.1002/gj.3250).
- (en) S. McLoughlin, « The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism », Australian Journal of Botany, vol. 49, , p. 271-300 (DOI 10.1071/BT00023)
- (en) A.M.C. Şengör, « Tectonics of the Tethysides: orogenic collage development in a collisional setting », Annual Review of Earth and Planetary Sciences, vol. 15, , p. 214-244 (DOI 10.1146/annurev.ea.15.050187.001241)
- M. Westphal, J. Orsini et P. Vellutini, « Le microcontinent Corso-Sarde, sa position initiale: données paléomagnétiques et raccords géologiques », Tectonophysics, vol. 30, nos 1-2, , p. 141-157 (DOI 10.1016/0040-1951(76)90142-6)
- (en) G. Cassinis, M. Durand et A. Ronchi, « Permian-Triassic continental sequences of Northwest Sardinia and South Provence: stratigraphic correlations and palaeogeographical implications », Bollettino della Società Geologica Italiana (Volume speciale), vol. 2, , p. 119-129 (lire en ligne)
- (en) M. Durand, « The problem of the transition from the Permian to the Triassic Series in southeastern France: comparison with other Peritethyan regions », dans Lucas, S.G., Cassinis, G. and Schneider, J.W., Non-Marine Permian Biostratigraphy and Biochronology, London, Geological Society, Special Publications, 265, (ISBN 978-1-86239-206-9), p. 281–296
- « Le volcanisme rhyolitique permien du Sud de la France : Estérel, Corse, Briançonnais et Béarn », sur Planet-Terre, Pierre Thomas (consulté le )
- (en) J.W. Schneider, S.G. Lucas, F. Scholze, S. Voigt, L. Marchetti, H. Klein, S. Opluštil, R. Werneburg, V.K. Golubev, J.E. Barrick, T. Nemyrovska, A. Ronchi, M.O. Day, V.V. Silantiev, R. Rößler, H. Saber, U. Linnemann, V. Zharinova et S-Z. Shen, « Late Paleozoic–early Mesozoic continental biostratigraphy — Links to the Standard Global Chronostratigraphic Scale », Palaeoworld, vol. 29, no 2, (DOI 10.1016/j.palwor.2019.09.001)
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2)
- (en) G. Gand et M. Durand, « Tetrapod footprint ichno-associations from French Permian basins. Comparisons with other Euramerican ichnofaunas », dans Lucas, S.G., Cassinis, G. and Schneider, J.W., Non-Marine Permian Biostratigraphy and Biochronology, London, Geological Society, Special Publications, 265, (ISBN 978-1-86239-206-9), p. 157–177
- (en) M. Durand, « Permian to Triassic continental successions in southern Provence (France): an overview », Bollettino della Società Geologica Italiana, vol. 127, no 3, , p. 697-716 (lire en ligne)
- (en) L. Marchetti, H. Klein, M. Buchwitz, A. Ronchi, R.M.H . Smith, W.J. De Klerk, L. Sciscio et G.H. Groenewald, « Permian-Triassic vertebrate footprints from South Africa: Ichnotaxonomy, producers and biostratigraphy through two major faunal crises », Gondwana Research, vol. 72, , p. 139-168 (DOI 10.1016/j.gr.2019.03.009)
- (en) L. Marchetti, S. Voigt et S.G. Lucas, « An anatomy-consistent study of the Lopingian eolian tracks of Germany and Scotland reveals the first evidence of the end-Guadalupian mass extinction at low paleolatitudes of Pangea », Gondwana Research, vol. 73, , p. 32-53 (DOI 10.1016/j.gr.2019.03.013)
- (en) L. Marchetti, A. Logghe, E. Mujal, B. Barrier, C. Montenat, A. Nel, J.-M. Pouillon, R. Garrouste et J.-S. Steyer, « Vertebrate tracks from the Permian of Gonfaron (Provence, Southern France) and their implication for the late Capitanian terrestrial extinction event », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 599, , p. 111043 (DOI 10.1016/j.palaeo.2022.111043)
- (en) D.S. Berman, H.C. Maddin, A.C. Henrici, S.S. Sumida, D. Scott et R.R. Reisz, « New primitive Caseid (Synapsida, Caseasauria) from the Early Permian of Germany », Annals of Carnegie Museum, vol. 86, no 1, , p. 43-75 (DOI 10.2992/007.086.0103)
- (en) R. Werneburg, F. Spindler, J. Falconnet, J.-S. Steyer, M. Vianey-Liaud et J.W. Schneider, « A new caseid synapsid from the Permian (Guadalupian) of the Lodève basin (Occitanie, France) », Palaeovertebrata, vol. 45 (2)-e2, , p. 1-36 (DOI 10.18563/pv.45.2.e2).