Callibrachion
Callibrachion gaudryi
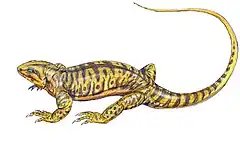
Callibrachion est un genre éteint de synapsides Caseidae ayant vécu dans le centre-est de la France durant le Permien inférieur (Assélien). L’holotype et unique spécimen connu (MNHN.F.AUT490) est représenté par un squelette postcrânien presque complet associé à des fragments de crâne découvert à la fin du XIXe siècle dans le bassin Permien d’Autun dans le département de Saône-et-Loire en Région Bourgogne-Franche-Comté. Il représente un individu immature mesurant moins de 1,5 m de long. Callibrachion fut longtemps considéré comme un synonyme du genre Haptodus et classé parmi les pélycosaures sphénacodontidés. En 2015, une nouvelle étude a révélé que Callibrachion était un animal différent d’Haptodus et qu’il appartenait aux caséasaures plutôt qu’aux sphénacodontidés. Ceci a été confirmé en 2016 par une analyse cladistique qui a récupéré Callibrachion comme un caséidé basal. Les dents pointues et la cage thoracique non élargie de Callibrachion indique que cet animal était probablement faunivore[1] - [2].
Description
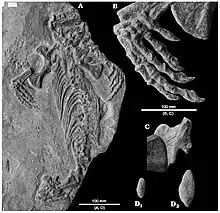
Le squelette de Callibrachion est relativement complet mais est mal conservé. Il est préservé sur une plaque de schiste noir qui expose la face ventrale du squelette. Ce dernier se compose d’un crâne fragmentaire, de la cage thoracique, des membres antérieurs, et du membre postérieur droit. Le membre postérieur gauche et la queue sont manquants ainsi que quelques éléments des épaules et du bassin. Le canal notochordal exposé et la désarticulation du scapulocoracoïde et de l’ilion (où la vascularisation osseuse est visible) révèle un stade ontogénétique immature. En revanche les métapodes bien ossifiés indiquent qu’il ne s’agit pas d’un juvénile précoce. Bien que presque complet, aucun caractère autapomorphique ne peut être identifié chez Callibrachion en raison de l’état de conservation généralement médiocre du spécimen ainsi que de la grande incomplétude du crâne qui concentre habituellement de nombreux caractères distinctifs. Callibrachion est défini par une combinaison de caractères tels que des dents coniques fines et droites, des côtes modérément épaissies, la possible présence d’un foramen ectépicondylien fermé, un ilion avec la lame dorsale haute typique des caséidés, des phalanges robustes mais non élargies, et une formule phalangienne non réduite[2].
Le crâne est représenté par plusieurs fragments dont seul le maxillaire (et possiblement une portion du prémaxillaire) est clairement identifiable par sa rangée de dents marginales. Le bord ventral est légèrement convexe, avec une région de dents caniniformes à l'extrémité antérieure indiquée par les bases dentaires larges contrairement aux plus petites conservées postérieurement à ces dernières. Une deuxième région de dents élargies est présente postérieurement aux positions caniniformes et rappelle ce qui est observé chez les éothyrididés et chez le caséidé basal Martensius. Il n’est pas possible de fournir un nombre précis de dents ou une estimation de la longueur du maxillaire, car le contour de l’élément est incertain. Les dents conservées ont une couronne mince et conique avec une surface lisse, dépourvue de toute courbure significative sauf dans la partie antérieure de la rangée de dents conservées. La mandibule est représentée par une branche mince et plutôt peu profonde de la mâchoire droite. Sa longueur conservée d’environ 70 mm pourrait être proche de sa longueur d’origine, car des dents larges et légèrement incurvées se trouvent dans ce qui est très probablement l’extrémité du dentaire[2].
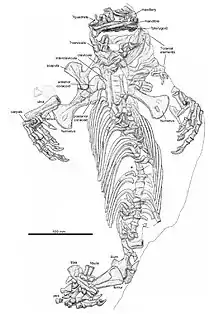
La colonne vertébrale n'est représentée que par environ douze centra de vertèbres dorsales, ainsi que des restes incertains de la série cervicale et des arcs neuraux. La queue n’est pas conservée. Deux centres cervicaux à peine visibles semblent se situer entre le crâne et la ceinture scapulaire. Certaines vertèbres ont été endommagées par la préparation, tandis que d’autres sont recouvertes par la ceinture scapulaire. Néanmoins, grâce à l’articulation du squelette, il est possible de fournir une estimation du nombre présacré. En particulier, la série ininterrompue d’environ 21 côtes dorsales indique un nombre présacré minimal de 23 vertèbres. Les côtes sont robustes. Dans la région dorsale postérieure, les côtes diminuent en longueur mais pas en largeur[2].
Les membres sont également robustes. La scapula se distingue notamment en étant beaucoup plus courte que l’humérus. Les deux mains sont assez bien conservées et articulées. Les os de la main sont robustes et bien développés. Cependant, les métacarpes et les phalanges ne présentent pas la morphologie trapue et élargie des caséidés dérivés. Leurs proportions sont plutôt similaires à celles observés chez Eocasea, Martensius, et Casea broilii. Le doigt IV est le plus long de la série. Une de ses phalanges médianes est raccourcie, mesurant environ 70 % de la longueur des phalanges voisines. Les phalanges terminales sont fortement incurvées, avec un fort tubercule fléchisseur et un toit dorsal plat élargi. La main présente la condition plésiomorphe des premiers amniotes avec une formule phalangienne de 2-3-4-5-3[N 1]. Les éléments conservés du pied droit sont pour la plupart désarticulés. Il n’est pas possible de préciser sa formule phalangienne mais les éléments individuels semblent être aussi robustes et fortement développés que ceux de la main[2].
Répartition stratigraphique
L’holotype de Callibrachion fut découvert au XIXe siècle sur le site de Margenne lors de l’exploitation du schiste bitumineux alors en cours dans le bassin d’Autun. La richesse paléontologique de ce bassin permit à l’époque de définir l’Autunien comme étage de référence pour le Permien inférieur continental d’Europe[3]. Selon la stratigraphie moderne, le site de Margenne appartient à la formation de Millery qui est l’une des formations les plus jeunes du bassin d’Autun[4]. Elle correspond à la partie supérieure de l’Autunien des anciens auteurs. En se basant sur les fossiles de plantes, d’invertébrés et d’amphibiens, l’Autunien fut considéré comme un âge équivalent à l’ensemble des étages marins globaux du Gzhélien supérieur, de l’Assélien, du Sakmarien et de l’Artinskien[3] - [5] - [4]. Depuis la fin des années 2010, des datations radiométriques de cendres volcaniques, altérées en tonsteins, et intercalées dans les couches sédimentaires de plusieurs formations géologiques constituant l’Autunien d’Autun ont révélé que l’Autunien correspondait presque exclusivement à l’étage Assélien de l’échelle stratigraphique internationale[6] - [7]. L’Autunien inférieur est représenté par les formations d’Igornay et de Muse. La partie médiane de la Formation d'Igornay est datée à 299,9 ± 0,38 Ma et représente le Gzhélien supérieur (Carbonifère terminal), confirmant les premières estimations de l’âge des plus anciens niveaux autuniens. Le lit des schistes bitumineux de Lally dans la partie inférieure de la formation de Muse est daté à 298,91 ± 0,08 Ma, ce qui correspond à ± 100 000 ans près, à la limite Carbonifère-Permien fixée en milieu marin dans l’Oural. La partie supérieure du lit des schistes bitumineux de Muse est datée entre 298,05 ± 0,19 et 298,57 ± 0,16 Ma. Ces datations démontrent que l’Autunien inférieur correspond au Gzhélien supérieur et à l’Assélien basal, et englobe donc la fin du Carbonifère supérieur et la base du Permien inférieur[6]. Deux autres niveaux de cendres volcaniques présents au sommet de la formation de Muse (limite Autunien inférieur-supérieur) et dans la partie supérieure de la formation de Millery ont donné des âges de 298,39 ± 0,09 Ma[N 2] et 297,7 ± 0,08 Ma, ce dernier indiquant un âge Assélien inférieur-moyen. Ces datations indiquent que l’Autunien supérieur (représenté par les formations de Surmoulin, de Millery, et de Curgy) n’a probablement pas dépassé un âge Assélien[8] - [7]. Ainsi, ces datations prouvent que les cinq formations géologiques du bassin d’Autun qui définirent l’Autunien représentent une durée géologique inférieure à 2,5 millions d’années et englobe la limite Carbonifère-Permien (Gzhélien supérieur - Assélien). Cette durée est nettement plus courte que les 10 millions d’années de dépôts estimés précédemment[7]. Ainsi, le genre Callibrachion de la formation de Millery, dont l’âge fut successivement considéré comme Sakmarien ou Artinskien, se révèle être plus vieux avec un âge Assélien inférieur ou moyen. Il est ainsi le plus ancien caséidé connu du Permien, seulement dépassé en âge par les taxons du Carbonifère supérieur Eocasea et Datheosaurus.
Paléoécologie
La formation de Millery atteint 250 m d’épaisseur et est constituée de schistes bitumineux gris foncé déposés dans un environnement lacustre[2] - [6]. Les cendres volcaniques altérées en tonsteins conservées dans ces dépôts sont liées à un volcanisme aérien explosif. L’origine exacte de ces cendres volcaniques est incertaine mais les volcans actifs les plus proches du bassin d’Autun durant la limite Carbonifère-Permien se trouvaient dans le nord du Massif central (régions de Blismes et Montreuillon) et dans la région de la Forêt-Noire et du Massif des Vosges[6]. Les rares amniotes trouvés dans ces couches lacustres, comme Callibrachion, le petit sphénacodontidé Haptodus et le taxon d’affinité incertaine “Belebey” augustodinensis[N 3] - [9] - [10] (venant tous les deux d’un autre site de la formation) sont allochtones. Le temnospondyle Onchiodon, un parent de plus petite taille du genre Eryops d’Amérique du Nord, est également présent[11] - [12] - [13].
Paléobiologie
Les dents coniques, minces, et peu recourbées du Callibrachion diffèrent nettement des dents spatulées ou en forme de feuille et pourvues de cuspides des caséidés herbivores plus tardif. Elles ressemblent davantage à celles des Eothyrididés et du caséidé basal Martensius. Les premiers étaient de petits prédateurs, le second était vraisemblablement insectivore au stade juvénile et omnivore à l’âge adulte[14]. Callibrachion ne possédait pas la cage thoracique élargie des caséidés herbivores qui abritait un intestin très développé nécessaire à la digestion de végétaux riches en fibre. L’absence de cette caractéristique chez Callibrachion couplé à sa denture particulière indiquent que cet animal avait probablement un régime alimentaire faunivore et devait se nourrir d’insectes et de petits vertébrés[2].
Classification
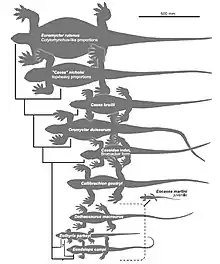
Depuis sa description en 1893 jusqu’à la fin des années 1970, la plupart des paléontologues ayant étudié Callibrachion l’ont estimé proche d’animaux classés aujourd’hui parmi les Sphenacodontidae. Certains auteurs pensaient également qu’il ne représentait pas un genre à part entière et qu’il pouvait être attribué au genre Haptodus. Dans les années 1990, Michel Laurent en est arrivé à douter de la nature synapside de Callibrachion (alors classé comme Haptodus gaudryi) et le considérait comme un nomen dubium[15]. Ce n’est qu’en 2015 que l’animal fut réétudié par Spindler et des collègues et identifié comme un caséasaure sur la base de ses proportions globales ainsi que de caractéristiques dentaires et ostéologiques qui l’excluent de tout autre clade de synapside[2]. En 2016, une étude phylogénétique des caséasaures récupère Callibrachion comme un caséidé basal proche d’Eocasea et Datheosaurus[16].
Ci-dessous, l’analyse phylogénétique publiée par Neil Brocklehurst et des collègues en 2016[16].
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- La formule phalangienne correspond au nombre de phalanges constituant chaque doigts des mains et des pieds des tétrapodes. Elle s’énumère en partant du doigt I (correspondant chez l’homme au pouce et au gros orteil) jusqu’au doigt V (l’équivalent de l’auriculaire et du petit orteil).
- L’incohérence entre cet âge de 298,39 ± 0,09 Ma du sommet de la formation de Muse avec l’âge plus jeune de 298,05 ± 0.19 Ma provenant d’un niveau pourtant plus ancien de la même formation pourrait s’expliquer par un âge rajeuni du tonstein daté à 298,05 Ma. Ce rajeunissement serait dû à la plus petite taille des grains de zircons datés et à une possible perte en plomb (réf. Mercuzot 2020, p. 249-250).
- Décrit comme un bolosauridé du genre Belebey par Jocelyn Falconnet (2012), ce taxon est considéré par Spindler et des collègues (2019) comme un nomen dubium, représentant possiblement un Edaphosauridae ou un Captorhinidae
Références
- M. Boule et P. Glangeaud, « Le Callibrachion, nouveau reptile du Permien d’Autun », Comptes rendus hebdomadaires des Séances de l’Académie des Sciences, vol. 117 (19), , p. 646-648 (lire en ligne)
- (en) F. Spindler, J. Falconnet et J. Fröbisch, « Callibrachion and Datheosaurus, two historical and previously mistaken basal caseasaurian synapsids from Europe », Acta Palaeontologica Polonica, vol. 361 (3), , p. 597-616 (DOI 10.4202/app.00221.2015)
- J. Broutin, J-J. Châteauneuf, J. Galtier et A. Ronchi, « L’Autunien d’Autun reste-t-il une référence pour les dépôts continentaux du Permien inférieur d’Europe ? Apport des données paléobotaniques », Géologie de la France, vol. 2, , p. 17-31
- (en) R. Werneburg et J.W. Schneider, « Amphibian biostratigraphy of the European Permo-Carboniferous », dans Lucas, S.G., Cassinis, G. and Schneider, J.W., Non-Marine Permian Biostratigraphy and Biochronology, London, Geological Society, Special Publications, 265, (ISBN 978-1-86239-206-9), p. 201-215
- (en) J.W. Schneider et R. Werneburg, « Insect biostratigraphy of the Euramerican continental Late Pennsylvanian and Early Permian », dans Lucas, S.G., Cassinis, G. and Schneider, J.W., Non-Marine Permian Biostratigraphy and Biochronology, London, Geological Society, Special Publications, 265, (ISBN 978-1-86239-206-9), p. 325-336
- (en) P. Pellenard, G. Gand, M. Schmitz, J. Galtier, J. Broutin et J-S. Steyer, « High-precision U-Pb zircon ages for explosive volcanism calibrating the NW European continental Autunian stratotype », Gondwana Resarch, vol. 51, , p. 118-136 (DOI 10.1016/j.gr.2017.07.014)
- P. Pellenard, M. Mercuzot, G. Gand, M. Schmitz, C. Ducassou, S. Bourquin et L. Beccaletto, « Un nouveau modèle d’âge pour l’Autunien du bassin d’Autun à partir de datations U-Pb sur des tonsteins », Conférence RST-27e édition de la Réunion des Sciences de la Terre, , p. 729 (lire en ligne)
- M. Mercuzot, « Reconstitutions paléoenvironnementales et paléoclimatiques en contexte tardi-orogénique : cas des bassins fini-carbonifères à permiens du nord-est du Massif central, France », Rennes, Ph.D. Thesis, Science de la Terre, Université de Rennes 1,
- (en) J. Falconnet, « First evidence of a bolosaurid parareptile in France (latest Carboniferous-earliest Permian of the Autun basin) and the spatiotemporal distribution of the Bolosauridae », Bulletin de la Société Géologique de France, vol. 183 (6), , p. 495-508 (DOI 10.2113/gssgfbull.183.6.495)
- (en) F. Spindler, S. Voigt et J. Fischer, « Edaphosauridae (Synapsida, Eupelycosauria) from Europe and their relationship to North American representatives », PalZ, vol. 94, no 1, , p. 125-153 (DOI 10.1007/s12542-019-00453-2)
- (de) R. Werneburg, « Der Eryopide Onchiodon (Amphibia) aus dem Rotliegend des Beckens von Autun (Frankreich) », Freiberger Forschungsheft, vol. C 466, , p. 167-181
- (en) R. Werneburg et J-S. Steyer, « Redescription of the holotype of Actinodon frossardi GAUDRY, 1866 from the Lower Permian of France (Autun) », Geobios, vol. 32 (4), , p. 599-607 (DOI 10.1016/S0016-6995(99)80009-6)
- (en) R.R. Schoch, « The life cycle in late Paleozoic eryopid temnospondyls: developmental variation, plasticity and phylogeny », Fossil Record, vol. 24, , p. 295-319 (DOI 10.5194/fr-24-295-2021)
- (en) D.S. Berman, H.C. Maddin, A.C. Henrici, S.S. Sumida, D. Scott et R.R. Reisz, « New primitive Caseid (Synapsida, Caseasauria) from the Early Permian of Germany », Annals of Carnegie Museum, vol. 86(1), , p. 43-75 (DOI 10.2992/007.086.0103)
- (en) M. Laurin, « Re-evaluation of Cutleria wilmarthi, an Early Permian synapsid from Colorado », Journal of Vertebrate Paleontology, vol. 14(1), , p. 134-138 (DOI 10.1080/02724634.1994.10011544)
- (en) N. Brocklehurst, R.R. Reisz, V. Fernandez et J. Fröbisch, « A Re-Description of ‘Mycterosaurus’ smithae, an Early Permian Eothyridid, and Its Impact on the phylogeny of Pelycosaurian-Grade Synapsids », PLoS ONE, vol. 11, no 6, , e0156810 (DOI 10.1371/journal.pone.0156810)