Roténone
La roténone est une molécule organique qui fait partie de la classe des ichtyotoxines, naturellement produite par les racines et les tiges de certaines plantes tropicales. Ce flavonoïde est toxique pour les organismes qui possèdent des mitochondries, car c'est un inhibiteur du complexe I de la chaîne respiratoire mitochondriale, elles-mêmes indispensables à la respiration des cellules. Elle entre dans la composition de nombreux pesticides et insecticides.
| Roténone | |||
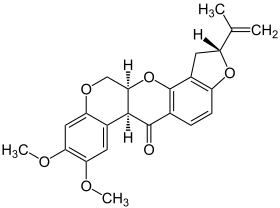
| |||
| Identification | |||
|---|---|---|---|
| Nom UICPA | (2R,6aS,12aS)-1,2,6,6a,12,12a-hexahydro-2-isopropényl-8,9-diméthoxychroméno[3,4-b]furo(2,3-h)chromén-6-one | ||
| Synonymes |
1,2,12,12a-tétrahydro-8,9-diméthoxy-2-(1-méthyléthényl-(1) benzopyrano (2,4-b) furo (2,3-h) (1)-benzopyran-6 (6H)-one; tubatoxine; paradéril |
||
| No CAS | |||
| No ECHA | 100.001.365 | ||
| No CE | 201-501-9 | ||
| No RTECS | DJ2800000 | ||
| PubChem | 6758 | ||
| ChEBI | 28201 | ||
| SMILES | |||
| InChI | |||
| Apparence | cristaux incolores[1] solide cristallin incolore à rouge et inodore[2] |
||
| Propriétés chimiques | |||
| Formule | C23H22O6 [Isomères] |
||
| Masse molaire[3] | 394,417 2 ± 0,021 7 g/mol C 70,04 %, H 5,62 %, O 24,34 %, |
||
| Propriétés physiques | |||
| T° fusion | 165 à 166 °C[1] - [2] | ||
| T° ébullition | 210 à 220 °C à 0,5 mmHg[2] | ||
| Solubilité | 0,17 mg·L-1 dans l'eau à 25 °C[2] sol. in éthanol, acétone, benzène, peu sol. in éther, très sol. in chloroforme[2] |
||
| Masse volumique | 1,27 g·cm-3 à 20 °C[2] - [1] | ||
| Pression de vapeur saturante | <0,01 µbar à 20 °C[2] | ||
| Cristallographie | |||
| Système cristallin | orthorhombique | ||
| Classe cristalline ou groupe d’espace | (no 19) [4] |
||
| Paramètres de maille | a = 836,90(10) pm b = 1977,1(2) pm c = 2326,9(2) pm |
||
| Précautions | |||
| SGH[5] | |||
  Danger |
|||
| Transport[2] | |||
|
|||
| Écotoxicologie | |||
| DL50 | 350 mg·kg-1 (souris oral)[2] 0,2 mg·kg-1 (rat, i.v.)[2] 2,8 mg·kg-1 (souris mâle, i.p.)[2] |
||
| LogP | 4,10[1] | ||
| Unités du SI et CNTP, sauf indication contraire. | |||


Étymologie
Le nom « roténone » vient de Roten, le nom japonais de la plante Derris elliptica Benth., (aujourd'hui Paraderris elliptica (Wall.) Adema). Il a été donné par le chimiste Nagai Nagayoshi qui a isolé cette molécule au Japon en 1902[6].
Histoire
Des plantes riches en roténone ont donc été utilisées et le sont encore pour les Nivrées (ou pêche à la nivrée) en Malaisie, en Afrique équatoriale et en Amazonie pour leurs vertus « ichtyotoxique » (qui tue les poissons).
Leur usage pourrait dater de plus de 2000 ans. Au Moyen Âge, les européens importaient via Venise, Gênes, Barcelone ou Marseille l’"Herbe Persanne" d'Indonésie ou de Malaisie en passant par le Moyen-Orient. Dès les années , on en a fait une poudre insecticide (produite à raison de milliers de tonnes par an à partir de plantes cultivées au Brésil, Pérou, Congo belge, en Afrique équatoriale française, au Cambodge et en Indonésie néerlandaise. Les importateurs les plus importants étaient néerlandais et belges avant les États-Unis et la France). À partir des années , le DDT ou l'HCH puis d'autres molécules insecticides ont remplacé la roténone. Puis pour des raisons environnementales (biodégradabilité), elle est à nouveau appréciée.
Chimie
La roténone a été isolée dans plusieurs espèces végétales qui poussent toutes sur des sols pauvres, généralement acides et en présence d’une atmosphère humide (75 % d'humidité minimum) :
- Derris elliptica Benth, Fabaceae, appelée Roten au Japon
- Lonchocarpus nicou D.C., Acesineae (liane) ; Emmanuel Geoffroy a extrait la même substance dès 1895 et donné le nom de nicouline (Mc Ewen & Stephenson 1976). Cette plante porte les noms de Barbasco en Espagne, Cubé au Brésil, Intchipari en Amazonie.
- Tephrosia vogelii Hook.f., nommée Bois Poison en Afrique.
Mais il en existe 64 autres dans la famille des Fabaceae.
Aspect
Poudre blanche cristalline.
Solubilité
La roténone est soluble dans l'acétone, le trichloréthylène, le chloroforme, certains solvants organiques mais non dans l'eau sous sa forme purifiée.
Réglementation
Sur le plan de la réglementation des produits phytopharmaceutiques dans l’Union européenne, cette substance active est interdite à la suite de l'examen relatif à l'inscription à l’annexe I de la directive 91/414/CEE. En France, les insecticides à base de roténone ont été retirés du marché à partir de 2008[7]. La décision de la Commission européenne no 2008/317/CE du demande à tous les États membres de retirer les autorisations des produits contenant de la roténone :
- à la commercialisation pour le .
- à l'utilisation pour le .
Par dérogation accordée par la Commission Européenne, la France bénéficie d'un délai supplémentaire pour des usages sur pommes, pêches, cerises, vigne et pommes de terre, en réservant l'emploi de la roténone aux utilisateurs professionnels munis d'un équipement de protection approprié. Le délai s'applique :
- à la commercialisation jusqu'au .
- à l'utilisation jusqu'au .
Par l'avis publié au Journal Officiel français du , la date limite d'écoulement des stocks en France a en fait été fixée :
- à la distribution au ,
- à l'utilisation au .
L'avis précise que, au cas où des alternatives en agriculture biologique ne seraient pas encore disponibles au , certaines autorisations de mise sur le marché pourraient être maintenues au titre des usages essentiels jusqu’à homologation d’une solution alternative, et au plus tard jusqu’au . Les usages qui seront potentiellement maintenus sont les suivants (sous réserve des dispositions relatives aux LMR) : pomme, poire, cerise, pêche, vigne et pommes de terre; avec une dérogation limitée aux utilisateurs professionnels avec les équipements de protection appropriés.
Effets physiopathologiques
Son activité toxique sur la physiologie, et sur les insectes en particulier, est liée au blocage de la synthèse de l'ATP, source d'énergie cellulaire, au niveau de la mitochondrie[8]. Elle bloque le flux d'électrons entre les centres fer-soufre (Fe-S) et l'ubiquinone, sur le complexe I (ou NADH - Coenzyme Q réductase) de la chaîne respiratoire mitochondriale[9].
Liens avec la Maladie de Parkinson
La maladie de Parkinson est la deuxième maladie neurodégénérative la plus fréquente après la maladie d'Alzheimer[10]. L'utilisation croissante de la roténone coïncide avec une augmentation des cas de la maladie de Parkinson[11] ; un doublement du nombre de cas est prévu entre et [10]. D'autres insecticides, synthétiques, sont également montrés du doigt comme le paraquat[12] - [13].
Chez l'humain, l'exposition à des pesticides inhibant le complexe I mitochondrial augmente le risque de développer la maladie de Parkinson (+ 70 %), et plus particulièrement la roténone (+ 150 %)[14] - [15]. Dans l'étude publiée par Tanner et ses collègues en dans la revue Environmental Health Perspectives les auteurs indiquent que :
« This study provides strong evidence of an association between rotenone use and PD in humans. PD developed 2.5 times as often in those who reported use of rotenone compared with nonusers, [...][14]. »
« Cette étude fournit des preuves solides d'un lien entre l'utilisation de la roténone et la maladie de Parkinson. Cette maladie est 2,5 fois plus fréquente chez les sujets ayant déclaré avoir utilisé de la roténone que chez ceux qui n'en utilisent pas, [...]. »
En , un modèle animal a été mis au point afin d'étudier l'effet d'une exposition à la roténone et un lien potentiel avec la maladie de Parkinson[16]. Ce modèle consistait à injecter de la roténone - à faibles doses, de manière chronique pendant cinq semaines et en intraveineuse via l'utilisation de pompes osmotiques - à des rats. Dans sa phase initiale, la neurotoxicité de la roténone n'entraîne aucun symptôme. Quand les premiers symptômes apparaissent, les dégâts sont déjà irrémédiables. La roténone conduit à l'apparition d'un syndrome Parkinsonien caractérisé par une hypokinésie, une posture voûtée et une sévère rigidité musculaire. Le modèle mis au point par Betarbet et ses collaborateurs était novateur en cela qu'il présentait des inclusions cytoplasmiques de type corps de Lewy, structures caractéristiques de la maladie de Parkinson[17]. La roténone a donc été utilisée pour mieux comprendre les mécanismes physiopathologiques et l'influence que pourrait jouer un facteur environnemental sur l'apparition de la maladie de Parkinson.
La roténone est un composé hautement lipophile, de ce fait il franchit facilement la barrière hémato-encéphalique l'amenant au contact du tissu cérébral. Sa toxicité provient, notamment, de son effet inhibiteur sur le complexe I de la chaîne respiratoire mitochondriale[17]. Ce faisant, elle génère un important stress oxydatif, inhiberait la formation des microtubules et l’activité du protéasome[18] - [19]. À la différence d'autres substances neurotoxiques, la roténone affecte de manière préférentielle les neurones dopaminergiques de la substance noire et conduit à leur dégénération[20]. La substance noire est une structure cérébrale impliquée, notamment, dans le contrôle de la motricité.
Si un certain nombre de critiques ont été apportées au modèle de Betarbet (voir plus bas), d'autres modèles, chez la souris et chez la mouche, sont venus valider l'effet de la roténone sur les neurones dopaminergiques[21] - [22]. A l'heure actuelle, la recherche scientifique favorise des études sur l'exposition à de faibles doses, plus proches de la réalité, et notamment lors de périodes de vulnérabilité accrue comme le développement[23] ou lorsque des composés chimiques sont mélangés (effet "cocktail")[24]. Par ailleurs, au sein de la population générale, la voie orale est habituellement considérée comme la voie d'exposition majoritaire[25].
C'est en ce sens qu'un modèle a été mis au point chez la souris[21]. Chez cet animal, l'ingestion chronique de roténone conduit à une dégénérescence des neurones dopaminergiques, de manière spécifique, une accumulation d'α-synucléine et des déficits comportementaux. Chez la mouche aussi, l'ingestion chronique d'une dose subléthale induit de sévères déficits locomoteurs ainsi qu'une perte des neurones dopaminergiques. Comme chez l'humain, l'ingestion de L-DOPA restaure partiellement les déficits locomoteurs. Les auteurs ne détectent cependant pas d'inclusions cytoplasmiques de types corps de Lewy dont un des constituants est absent chez la mouche. L'effet observé sur les neurones dopaminergiques, dans différentes espèces et avec différents modes d'administration, suggère fortement que ce pesticide pourrait participer à l'apparition des cas sporadiques de la maladie de Parkinson qui représentent 90 à 95 % des cas[22].
- Limites des modèles de la maladie de Parkinson
Comme tous les modèles d'études, celui mis au point par Betarbet et ses collaborateurs[16] présentait des limites et ne reproduisait que certains aspects de la maladie de Parkinson présents chez l'humain[17]. Un certain nombre de critiques ont ainsi été formulées concernant la variabilité individuelle de la réponse à la toxine, la forte mortalité associée à l'injection de roténone - due à une importante toxicité pour le cœur, le foie, les reins et le système gastro-intestinal - et enfin le fait que d'autres populations neuronales des ganglions de la base soient atteintes[18]. Il n'en reste pas moins que ce modèle est un jalon majeur dans l'étude de la maladie de Parkinson et qu'il a permis de très importantes avancées dans la compréhension des atteintes mitochondriales intervenant dans le cadre de cette maladie[17].
Dans l'expérience de l'université Emory, la roténone a été mélangée à des produits qui ne sont pas utilisés en agriculture et qui ont la propriété de favoriser la pénétration des molécules à travers les tissus. Néanmoins l'usage de pénétrants, adjuvants ajoutés aux préparations commerciales, qui permettent de faire passer les molécules actives dans les plantes ou les insectes, peut avoir un effet similaire et augmenter considérablement la toxicité de la roténone. L'objet de l'étude n'était pas d'incriminer l'utilisation agricole de la roténone dans le développement de la maladie de Parkinson. L'auteur précise d'ailleurs lui-même que la roténone a une toxicité faible quand elle est administrée par voie orale. Le but du chercheur était simplement de trouver un moyen d'obtenir des rats atteints de la maladie de façon régulière pour mieux pouvoir étudier les lésions qu'elle provoque dans les tissus du cerveau.
Hypothèse intestinale
La recherche de modèles plus conformes à la maladie de Parkinson a amené à diminuer les doses de roténone auxquelles étaient soumis les animaux[26]. Ainsi, dans une étude publiée en , les doses administrées oralement à des souris ne permettaient pas de détecter la présence du pesticide dans le sang ou le système nerveux central ; le complexe I mitochondrial n'apparaissait pas inhibé non plus. Les auteurs de l'étude observèrent cependant une accumulation d'α-synucléine dans le système nerveux entérique, le noyau dorsal du nerf vague et la substance noire. Les souris présentaient également des signes d’inflammation, une perte de neurones dopaminergiques dans la substance noire et une altération de la coordination motrice après trois mois de traitement. Les auteurs ont alors fait l’hypothèse que les lésions produites par la roténone se propageaient à partir du système nerveux entérique vers les régions cérébrales auxquelles il est connecté par des synapses.
Ces résultats sont venus renforcer une hypothèse selon laquelle la maladie de Parkinson aurait une origine intestinale[27]. En effet, si la maladie est surtout connue pour ses manifestations comportementales, il existerait une phase pré-symptomatique qui pourrait être longue (une dizaine d'années) et caractérisée par d'autres symptômes non moteurs, notamment gastro-intestinaux. Durant cette période, une forme de l'α-synucléine présentant un défaut de conformation pourrait se propager, de cellule en cellule, à la manière des protéines de type prion[28]. Lorsque cette α-synucléine mal repliée est capturée par un autre neurone, elle va servir de modèle pour le repliement incorrect d’autres α-synucléines saines présentent dans le neurone. L'accumulation de ces α-synucléines mal repliées conduit à la formation d’agrégats et éventuellement des corps de Lewy. Si ces derniers ont été décrits, à l'origine, dans le cerveau, on en retrouve également dans le système nerveux périphérique, y compris le système nerveux entérique.
Une étude pathologique a révélé une progression anatomique chronologique de la maladie de Parkinson[29]. Une atteinte précoce du bulbe olfactif et du nerf vague suggère une diffusion, à partir de ces sites périphériques, vers le cerveau, et ce d'autant plus que l'α-synucléine peut être transportée de manière antérograde et surtout rétrograde dans le nerf vague[27]. Cette hypothèse est fortement renforcée par le fait qu'une vagotomie - la section du nerf vague - empêche la progression du syndrome Parkinsonien chez le modèle de souris ayant consommé de la roténone[30]. Chez l'humain, les patients ayant subi une vagotomie ont également moins de risque de développer la maladie de Parkinson[27]. La roténone induit un relargage d'α-synucléine par les neurones entériques, celle-ci est captée par les terminaisons pré-synaptiques puis transportée de manière rétrograde jusqu'aux noyaux moteurs dorsaux du nerf vague[30].
La porte d'entrée depuis la lumière intestinale vers le système nerveux entérique puis le nerf vague pourrait être une catégorie de cellules particulières, les cellules entéro-endocrines[31]. Ces cellules, localisées dans la muqueuse du tractus gastro-intestinal, sont excitables électriquement (comme les neurones) et présentent des capacités chimiosensorielles. Leur face apicale est orientée vers la lumière de l'intestin et la face basale contient de nombreux granules sécrétoires. Ces cellules secrètent des hormones en réponse à la présence de certains nutriments ou bactéries dans l'intestin. Elles possèdent également de nombreuses caractéristiques similaires aux neurones, récepteurs aux neurotrophines, protéines pré- et post-synaptiques, neuropodes. Un circuit neuronal relie donc de cette manière la lumière de l'intestin au système nerveux et constitue ainsi un porte d’entrée éventuelle pour des composés toxiques comme la roténone[27].
Atteintes neurodéveloppementales
Le développement cérébral est une période de vulnérabilité accrue pour le système nerveux[32]. Durant cette période, le cerveau ne dispose pas encore de structures de soutien, comme les astrocytes, pouvant le protéger contre les agressions de xénobiotiques tels que les pesticides. L'exposition à ce type de molécules pourrait être à l'origine de l'augmentation récente des troubles neurodéveloppementaux[33]. Même à faibles doses, certains composés pourraient avoir des conséquences importantes sur le développement du système nerveux et plus tard sur le comportement chez l'adulte[34].
L'effet de la roténone sur le développement cérébral a été évalué en utilisant un modèle organotypique de cerveau humain appelé sphéroïdes cérébraux[35]. Ce modèle est une mise en culture, en trois dimensions, de cellules neurales issues de cellules souches pluripotentes induites (CSPi) humaines. Ces cellules vont générer des sphéroïdes constitués de différents types cellulaires - neurones, cellules gliales - permettant de reproduire en culture certains aspects des phases initiales du développement cérébral. L'un des intérêts majeur de ce modèle étant qu'il est possible de générer ce type de culture à partir de CSPi issues de chaque individu et ainsi de réaliser une médecine personnalisée en fonction du patrimoine génétique de l'individu.
Les auteurs vont mettre en évidence un pouvoir de toxicité accrue de la roténone - caractérisé par la génération de dérivés réactifs de l'oxygène et un dysfonctionnement mitochondrial - dans les phases précoces de différenciation cellulaires. Des analyses morphologiques vont indiquer que la roténone s’avère toxique pour les neurones dopaminergiques, spécifiquement, à des doses normalement considérés comme non-toxiques (10−6 mol/l). Les autres populations neuronales et les astrocytes sont affectées à des concentrations 25 fois supérieures. L'analyse de l'expression des gènes et du métabolisme à large échelle indiquent que la roténone pourrait agir sur le neurodéveloppement en altérant la croissance des neurites, venant ainsi confirmer d'autres études[36].
Ecotoxicité
La molécule étant instable dans l'eau et exposée à la lumière, elle se dégrade et perd sa toxicité avec le temps, plus ou moins vite selon l'environnement. Il semble que dans le cas des poissons, la roténone agisse dès le contact avec les branchies. Elle ne pénétrerait pas la chair, qui peut donc être consommée, d'autant que la cuisson dégrade la molécule.
Rituellement, les indiens wayana ne doivent pas boire une autre boisson que de l'eau réservée sur laquelle flottent des roseaux durant l'opération de la nivrée. Le sens donné à cet acte est-il uniquement symbolique (il s'agirait par une relation magique de faire en sorte que comme le roseau qui flotte, les poissons asphyxiés remontent et flottent aussi), ou la tradition indique-t-elle aussi qu'il ne faut pas boire l'eau empoisonnée par le suc des lianes utilisées pour tuer les poissons ?
Notes et références
- ROTENONE, Fiches internationales de sécurité chimique
- PubChem CID 6758
- Masse molaire calculée d’après « Atomic weights of the elements 2007 », sur www.chem.qmul.ac.uk.
- Structure cristallographique (CCDC)
- Numéro index dans le tableau 3.1 de l'annexe VI du règlement CE N° 1272/2008 (16 décembre 2008)
- Dajoz 1969-Matsumura 1975
- France. « Code rural », Mise sur le marché des produits antiparasitaires à usage agricole, art. L. 253-1 à L. 253-17. (version en vigueur : 21 août 2008) [lire en ligne (page consultée le 15 mai 2020)]
- (en) Donald Voet et Judith G. Voet, Biochemestry, New York, Wiley, , 2e éd., 1392 p. (ISBN 978-0-471-58651-7), chap. 20 (« Electron Transport and Oxidative Phosphorylation »), p. 569-575.
- (en) Cicchetti F, Drouin-Ouellet J et Gross RE., « Environmental Toxins and Parkinson's Disease: What Have We Learned From Pesticide-Induced Animal Models? », Trends Pharmacol Sci., vol. 30, no 9, , p. 475-83 (PMID 19729209, DOI 10.1016/j.tips.2009.06.005, lire en ligne, consulté le )
- (en) Poewe W, Seppi K, Tanner CM, Halliday GM, Brundin P, Volkmann J, Schrag AE et Lang AE., « Parkinson disease. », Nat Rev Dis Primers., vol. 3, , p. 17013 (PMID 28332488, DOI 10.1038/nrdp.2017.13, lire en ligne [PDF], consulté le )

- « la-rotenone »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?)
- « Parkinson-de-nouveaux-pesticides-incriminés »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?)
- Inserm (direction), Pesticides : Effets sur la santé, Paris, Inserm, coll. « Expertise collective », , XII-1001 p. (ISBN 978-2-85598-905-1, OCLC 875398182, lire en ligne), chap. 12 (« Maladie de Parkinson »).
- (en) Tanner CM, Kamel F, Ross GW, Hoppin JA, Goldman SM, Korell M et al., « Rotenone, paraquat, and Parkinson's disease. », Environ Health Perspect., vol. 119, no 6, , p. 866-72 (PMID 21269927, PMCID 3114824, DOI 10.1289/ehp.1002839, lire en ligne [PDF], consulté le )
- Inserm 2013, chap. 12, p. 452.
- (en) Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV et Greenamyre JT., « Chronic systemic pesticide exposure reproduces features of Parkinson's disease. », Nat Neurosci., vol. 3, no 12, , p. 1301-6 (PMID 11100151, DOI 10.1038/81834, lire en ligne [PDF], consulté le )
- (en) Blandini F et Armentero MT., « Animal models of Parkinson's disease. », FEBS J., vol. 279, no 7, , p. 1156-66 (PMID 22251459, DOI 10.1111/j.1742-4658.2012.08491.x, lire en ligne [EPUB], consulté le )
- (en) Bové J et Perier C., « Neurotoxin-based models of Parkinson's disease. », Neuroscience, vol. 211, , p. 51-76 (PMID 22108613, DOI 10.1016/j.neuroscience.2011.10.057, lire en ligne, consulté le )
- (en) Choi WS, Palmiter RD et Xia Z., « Loss of mitochondrial complex I activity potentiates dopamine neuron death induced by microtubule dysfunction in a Parkinson's disease model. », J Cell Biol., vol. 192, no 5, , p. 873-82 (PMID 21383081, PMCID 3051820, DOI 10.1083/jcb.201009132, lire en ligne [PDF], consulté le )
- (en) Sherer TB, Betarbet R, Testa CM, Seo BB, Richardson JR, Kim JH, Miller GW, Yagi T, Matsuno-Yagi A et Greenamyre JT., « Mechanism of toxicity in rotenone models of Parkinson's disease. », J Neurosci., vol. 23, no 34, , p. 10756-64 (PMID 14645467, PMCID 6740985, DOI 10.1523/JNEUROSCI.23-34-10756.2003, lire en ligne [PDF], consulté le )
- (en) Inden M, Kitamura Y, Abe M, Tamaki A, Takata K et Taniguchi T., « Parkinsonian rotenone mouse model: reevaluation of long-term administration of rotenone in C57BL/6 mice. », Biol Pharm Bull., vol. 34, no 1, , p. 92-6 (PMID 21212524, DOI 10.1248/bpb.34.92, lire en ligne [PDF], consulté le )
- (en) Coulom H et Birman S., « Chronic exposure to rotenone models sporadic Parkinson's disease in Drosophila melanogaster. », J Neurosci., vol. 24, no 48, , p. 10993-8 (PMID 15574749, PMCID 6730201, DOI 10.1523/JNEUROSCI.2993-04.2004, lire en ligne [PDF], consulté le )
- (en) Grandjean P, Bellinger D, Bergman A et al., « The faroes statement: human health effects of developmental exposure to chemicals in our environment. », Basic Clin Pharmacol Toxicol., vol. 102, no 2, , p. 73-5 (PMID 18226057, DOI 0.1111/j.1742-7843.2007.00114.x, lire en ligne [EPUB], consulté le )
- (en) Graillot V, Takakura N, Hegarat LL, Fessard V, Audebert M et Cravedi JP., « Genotoxicity of pesticide mixtures present in the diet of the French population. », Environ Mol Mutagen., vol. 53, no 3, , p. 173-84 (PMID 22389207, DOI 10.1002/em.21676, lire en ligne, consulté le )
- Inserm 2013, chap. 2, p. 43.
- (en) Pan-Montojo F, Anichtchik O, Dening Y, Knels L, Pursche S, Jung R, Jackson S, Gille G, Spillantini MG, Reichmann H et Funk RH., « Progression of Parkinson's disease pathology is reproduced by intragastric administration of rotenone in mice. », PLoS One., vol. 5, no 1, , e8762 (PMID 20098733, PMCID 2808242, DOI 10.1371/journal.pone.0008762, lire en ligne [PDF], consulté le )
- (en) Liddle RA., « Parkinson's disease from the gut. », Brain Res., vol. 1693, no Pt B, , p. 201-206 (PMID 29360467, PMCID 6003841, DOI 10.1016/j.brainres.2018.01.010, lire en ligne [PDF], consulté le )
- (en) Desplats P, Lee HJ, Bae EJ, Patrick C, Rockenstein E, Crews L, Spencer B, Masliah E et Lee SJ., « Inclusion formation and neuronal cell death through neuron-to-neuron transmission of alpha-synuclein. », Proc Natl Acad Sci U S A., vol. 106, no 31, , p. 13010-5 (PMID 19651612, PMCID 2722313, DOI 10.1073/pnas.0903691106, lire en ligne [PDF], consulté le )
- (en) Braak H, Del Tredici K, Bratzke H, Hamm-Clement J, Sandmann-Keil D et Rüb U., « Staging of the intracerebral inclusion body pathology associated with idiopathic Parkinson's disease (preclinical and clinical stages). », J Neurol., vol. 249, no Suppl 3, , III/1-5 (PMID 12528692, DOI 10.1007/s00415-002-1301-4, lire en ligne, consulté le )
- (en) Pan-Montojo F, Schwarz M, Winkler C, Arnhold M, O'Sullivan GA, Pal A, Said J, Marsico G, Verbavatz JM, Rodrigo-Angulo M, Gille G, Funk RH et Reichmann H., « Environmental toxins trigger PD-like progression via increased alpha-synuclein release from enteric neurons in mice. », Sci Rep., vol. 2, , p. 898 (PMID 23205266, PMCID 3510466, DOI 10.1038/srep00898, lire en ligne [PDF], consulté le )
- (en) Chandra R, Hiniker A, Kuo YM, Nussbaum RL et Liddle RA., « α-Synuclein in gut endocrine cells and its implications for Parkinson's disease. », JCI Insight., vol. 2, no 12, , p. 92295 (PMID 28614796, PMCID 5470880, DOI 10.1172/jci.insight.92295, lire en ligne [PDF], consulté le )
- (en) Rice D et Barone S Jr., « Critical periods of vulnerability for the developing nervous system: evidence from humans and animal models. », Environ Health Perspect., vol. 108, no Suppl 3, , p. 511-33 (PMID 10852851, PMCID 1637807, DOI 10.1289/ehp.00108s3511, lire en ligne [PDF], consulté le )
- (en) Grandjean P et Landrigan PJ., « Developmental neurotoxicity of industrial chemicals. », Lancet, vol. 368, no 9553, , p. 2167-78 (PMID 17174709, DOI 10.1016/S0140-6736(06)69665-7, lire en ligne [PDF], consulté le )
- (en) Grandjean P et Landrigan PJ., « Neurobehavioural effects of developmental toxicity. », Lancet Neurol., vol. 13, no 3, , p. 330-8 (PMID 24556010, PMCID 4418502, DOI 10.1016/S1474-4422(13)70278-3, lire en ligne [PDF], consulté le )
- (en) Pamies D, Block K, Lau P, Gribaldo L, Pardo CA, Barreras P, Smirnova L, Wiersma D, Zhao L, Harris G, Hartung T et Hogberg HT., « Rotenone exerts developmental neurotoxicity in a human brain spheroid model. », Toxicol Appl Pharmacol., vol. 354, , p. 101-114 (PMID 29428530, PMCID 6082736, DOI 10.1016/j.taap.2018.02.003, lire en ligne [EPUB], consulté le )
- (en) Krug AK, Balmer NV, Matt F, Schönenberger F, Merhof D et Leist M., « Evaluation of a human neurite growth assay as specific screen for developmental neurotoxicants. », Arch Toxicol., vol. 87, no 12, , p. 2215-31 (PMID 23670202, DOI 10.1007/s00204-013-1072-y, lire en ligne, consulté le )
Annexes
Bibliographie
- Thany SH, Reynier P et Lenaers G., « Neurotoxicité des pesticides : Quel impact sur les maladies neurodégénératives ? », Med Sci (Paris)., vol. 29, no 3, , p. 273-8 (PMID 23544381, DOI 10.1051/medsci/2013293013, lire en ligne, consulté le ).
Articles connexes
Liens externes
- Ressources relatives à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- (fr) Roténone et Parkinson
- Substance active d'un produit phytopharmaceutique
- Liste de substances actives de produits phytosanitaires
- Liste de substances actives de produits phytopharmaceutiques autorisées par l'Union européenne
- Liste de substances actives de produits phytopharmaceutiques interdites par l'Union européenne